蠲痹顆粒醇提物調控MAPK信號轉導抑制T細胞活化效應機制的研究*
2022-11-11 06:49:54鄒楠婷顧茜蘭楊海浩李小絲彭江云萬春平
云南中醫學院學報 2022年3期
關鍵詞:小鼠
鄒楠婷,吳 招,顧茜蘭,應 賽,楊海浩,祁 燕,李小絲,彭江云,萬春平
(云南中醫藥大學,云南 昆明 650500)
蠲痹顆粒是由云南省當代扶陽學派杰出代表、著名中醫學家吳生元教授研制的抗類風濕關節炎的經驗藥方[1],1995年獲院內制劑批件(滇ZJGF/1995-25),2008年獲再注冊批件(滇藥制字〈Z〉20082554A),2012年8月獲國家發明專利(專利號:201010185717.X)。處方由附片、川芎、桂枝、五加皮等藥味配伍而成,具有溫陽通絡、祛風除濕、散寒、消腫止痛的功效,主治風寒濕痹證。
前期臨床觀察和實驗研究證實:蠲痹顆粒對活動期類風濕關節炎有顯著臨床療效,能減輕病人的臨床癥狀,下調C-反應蛋白(CRP)和血沉(ESR)等有關治療指標。此外,蠲痹顆粒對大鼠佐劑性關節炎(AA)和小鼠膠原關節炎(CIA)有顯著的抗炎和鎮痛作用[2-4],提示蠲痹顆粒防治類風濕關節炎與其抑制T細胞介導的免疫反應密切相關。但該藥的確切免疫調節機制尚未明確,因此有必要進一步研究其治療類風濕關節炎的免疫學機制。
最新研究顯示[5-7],絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信號通路與RA時T淋巴細胞激活和細胞因子分泌具有密切聯系,已成為RA藥物治療的重要作用靶點。該通路主要由ERK、p38和JNK激酶組成的。本研究通過建立體外細胞分化模型,探討蠲痹顆粒醇提物(JBKL)對T、B淋巴細胞體外增殖的影響,及其調控MAPK信號轉導干預T細胞活化效應機制的研究,為扶陽法治療風寒濕痹證類風濕關節炎提供理論依據和科學基礎。
1 材料與方法
1.1 材料 BALB/c小鼠,雌性,78周齡,體質量(18±1)g,購自斯貝福(北京)生物技術有限公司,合格證號:SCXK(京)2019-0010;實驗動物在 SPF級動物房飼養1周后使用。環境溫度濕度分別為(22±1)℃和(55±5)%,12 h晝夜循環。動物自主攝取滅菌或消毒后的飼料和水。所有實驗均嚴格按照實驗動物相關條例執行。
蠲痹顆粒浸膏醇提物制備:稱取289.4 g蠲痹顆粒浸膏,用蒸餾水加熱溶解完全,冷卻后加入乙醇混勻,直至乙醇含量為80%,放置12 h后離心,取上清液過濾,并減壓回收乙醇,得蠲痹顆粒藥材浸膏,將浸膏置于真空干燥箱真空干燥約2 d,得蠲痹顆粒固體浸膏3.658 g,折合原藥材:1 g醇提物相當于79.11 g蠲痹顆粒浸膏。
RPMI-1640培養基購自 GibcoBRL公司(life Technologies,Grandisland,NY,USA);胎牛血清(fetal calf serum,FBS)和 MTT均購買于 Hyclone公司(Logan,Utah,USA);刀豆蛋白 A(Con A)和細菌脂多糖(LPS)均購自美國 SIGMA 公司(St.Louis,MO,USA);IL-10、IL-6、IL-17A 和 IFN-γ細胞因子 ELISA 檢測試劑盒和Anti-CD3抗體均購自BD PharMingen公司(San Diego,CA,USA);抗 phospho-ERK1/2,phosphop38購自Cell Signaling Technology;其它試劑為國產分析純。
Megafuge l1.0R離心機(德國ThermoScientific Heraeus);二氧化碳培養箱購自美國Thermo-Fisher;Epoch連續波長酶標儀來自美國Bio-Tek公司;DM2500正置顯微鏡(德國萊卡儀器有限公司);伯樂小型垂直電泳套裝及通用電泳儀164-5070(美國Bio-Rad)、ODYSSEYCLx激光成像系統(美國 LICOR公司)。
1.2 脾淋巴細胞和純化CD4+T細胞制備 脫脊椎處死小鼠,摘取脾臟,研磨獲取脾細胞懸液,用紅細胞裂解液裂解紅細胞,用PBS洗2~3次,離心濾過后加入含2%FBS的RPMI-1640混勻得到脾淋巴細胞,并將細胞調至所需濃度。采用免疫磁珠分離法(陰性選擇),加入小鼠抗小鼠I-Ad/b、同山羊抗小鼠抗體IgG結合磁珠,除去與磁珠結合I-A+抗原遞呈細胞和Ig+的B細胞,獲得純化的T細胞。CD4+T細胞的純化在上述純化條件下加入抗CD8抗體標記磁珠,除去CD8+T細胞,得到純化CD4+T細胞。以上操作均需無菌處理。
1.3 JBKL對小鼠脾淋巴細胞增殖功能的影響 以5×105/孔的BALB/c小鼠脾淋巴細胞數接種于96孔板,藥物組加入低、中、高劑量(3、10、30 μg/m L)的JBKL,再加入 5 μg/mL ConA 或 LPS(10 μg/mL)刺激T、B淋巴細胞增殖。另外,設不加刺激劑和藥物的空白組,以及只加刺激劑的對照組,放入37℃,5%CO2培養箱,培養第44 h后每孔加入5 mg/mL MTT 20 μL,再孵育4 h結束后,吸除上清,每孔加入100μL DMSO,低速振蕩充分溶解反應后的MTT結晶,用酶標儀在570 nm波長處檢測各孔的吸光度。對于Anti-CD3 mAb誘導T細胞增殖,方法如下:5μg/m L Anti-CD3 mAb稀釋溶液(碳酸鹽緩沖液稀釋,pH 9.6)包被96孔培養板,誘導T淋巴細胞增殖,于37℃培養箱中放置2 h后,丟棄液體,用PBS洗1次,加入脾淋巴細胞(5×105/孔)和不同濃度JBKL,之后的方法同上均用MTT法檢測。
1.4 MTT檢測JBKL對小鼠脾淋巴細胞細胞毒性檢測往96孔板內加入4×105/孔的正常小鼠脾淋巴細胞,同時加入 JBKL(0、3、10、30 μg/m L)。37℃、5%CO2培養箱中培養44 h后,每孔加入20μL 5 mg/mL MTT,繼續孵育4 h。培養結束時,棄去原液,每孔加入100 μL DMSO,充分溶解結晶,用酶標儀于570 nm處測定吸光度。
1.5 JBKL對Anti-CD3 mAb誘導T細胞細胞因子產生的影響 純化的CD4+T淋巴細胞(4×105/孔)接種于包被著anti-CD3 mAb(5μg/mL)的24孔培養板中,同時加入不同濃度的 JBKL(0、3、10、30 μg/mL)。對照組則不含anti-CD3 mAb。37℃,5%CO2培養箱中培養,36 h后收集細胞上清液,離心(5 000 r/min,4℃,5 min),根據相應 IL-10、IL-6、IFN-γ 和 IL-17A的ELISA試劑盒說明書檢測細胞培養液中各細胞因子的含量。
1.6 CD4+T細胞MAPK信號通路的相關蛋白表達水平 PBS稀釋的anti-CD3 mAb(5μg/mL)加入24孔培養板,于4℃包被過夜,用PBS洗2次。加入CD4+T淋巴細胞(5×106/孔)、anti-CD28 mAb(2 μg/m L)和不同濃度的蠲痹顆粒提取物(0、3、10、30、100 μg/m L),刺激30 min,另設無刺激對照組。收集細胞于Eppendorf管中,每管分別加入等量的裂解溶液(RIPA、蛋白酶抑制劑和磷酸化蛋白酶抑制劑),4℃裂解 30 min,離心(4 ℃、10 min、12 000 r/min),吸取上清獲得細胞總蛋白。總蛋白按照BCA蛋白濃度測定試劑盒說明書測定后再把濃度調成一致,取相同體積樣品加入適量SDS,100℃水浴煮沸10 min變性總蛋白。將總蛋白進行垂直電泳分離、轉膜、封閉、洗膜,分別加入抗體p-ERK和p-P38,4℃孵育6 h,洗膜,加入相應二抗室溫避光孵育1 h,棄去二抗洗膜后于ODYSSEYCLx雙色紅外激光成像儀上掃膜,用ImageStudio軟件采集分析蛋白條帶圖像。以甘油醛-3-磷酸脫氫酶(GAPDH)的顯色為內參。
1.7 統計學分析 用SPSS 17.0軟件對數據結果進行分析,所有數據都以均數±標準差(±s)表示,組間比較使用t檢驗和單因素方差分析處理,P<0.05為差異有統計學意義。
2 結果
2.1 JBKL對小鼠脾淋巴細胞活性和細胞毒性的影響 圖1A結果顯示,正常小鼠T淋巴細胞在絲裂原Con A誘導下顯著增殖;與溶媒對照組比較,JBKL 30μg/mL藥物干預后,能夠顯著抑制Con A誘導小鼠T淋巴細胞增殖能力。圖1B結果表明,與溶媒對照組相比,JBKL 30μg/mL顯著抑制Anti-CD3 mAb誘導正常小鼠脾臟T淋巴細胞活化。圖1C結果表明,LPS顯著刺激正常小鼠B淋巴細胞發生增殖,與溶媒對照組相比,JBKL 30μg/m L藥物干預顯著抑制正常小鼠B淋巴細胞的增殖活性。圖1D結果表明,與溶媒對照組相比,JBKL在濃度(3~30)μg/m L對正常脾淋巴細胞活度無明顯的影響,提示 JBKL在(3~30)μg/mL對正常小鼠淋巴細胞無明顯細胞毒性作用。上述結果表明,JBKL體外具有顯著的免疫抑制活性。
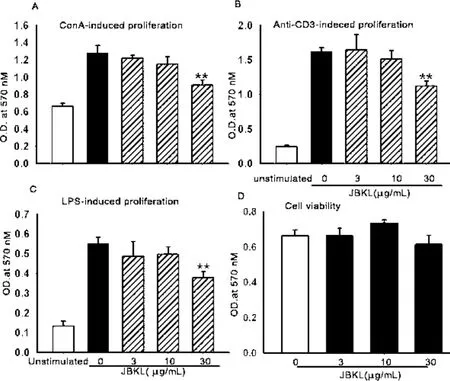
圖1 不同濃度蠲痹顆粒醇提物(JBKL)對 ConA(A)、Anti-CD3 m Ab(B)和 LPS(C)誘導的正常小鼠脾淋巴細胞增殖情況影響和細胞毒性(D)(±s,n=3)
2.2 JBKL對TCR信號誘導的CD4+T淋巴細胞細胞因子產生的影響 為進一步探究JBKL對細胞因子產生的影響,本研究應用ELISA法檢測JBKL對Anti-CD3 mAb誘導CD4+T淋巴細胞細胞因子產生的影響。ELISA檢測結果顯示,CD4+T淋巴細胞經 Anti-CD3 mAb誘導后分泌更多的 IFN-γ、IL-17A、Treg細胞因子(IL-10)和促炎細胞因子(IL-6)。給予不同濃度的JBKL作用后,產生的IFN-γ和IL-17A顯著低于溶媒對照組。同時大幅增加Treg負調控細胞因子(IL-10)的分泌量,降低IL-6產生水平(表1)。
表1 蠲痹顆粒醇提物(JBKL)對Anti-CD3 m Ab誘導CD4+T細胞細胞因子的影響(±s,n=3,pg/mL)
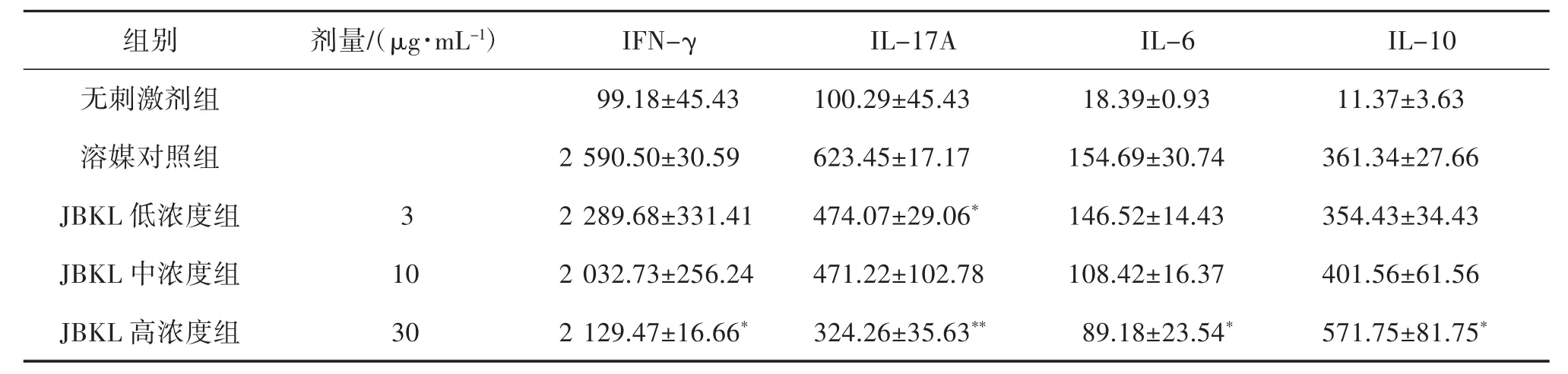
表1 蠲痹顆粒醇提物(JBKL)對Anti-CD3 m Ab誘導CD4+T細胞細胞因子的影響(±s,n=3,pg/mL)
注:與溶媒對照組比較,*P<0.05,**P<0.01。
組別劑量/(μg·mL-1)IFN-γIL-17A IL-6 IL-10無刺激劑組 99.18±45.43 100.29±45.43 18.39±0.93 11.37±3.63溶媒對照組 2 590.50±30.59 623.45±17.17 154.69±30.74 361.34±27.66 JBKL 低濃度組 3 2 289.68±331.41 474.07±29.06* 146.52±14.43 354.43±34.43 JBKL 中濃度組 10 2 032.73±256.24 471.22±102.78 108.42±16.37 401.56±61.56 JBKL 高濃度組 30 2 129.47±16.66* 324.26±35.63** 89.18±23.54* 571.75±81.75*
2.3 JBKL干預后,CD4+T淋巴細胞磷酸化p38和ERK蛋白水平變化 圖2結果顯示,30μg/mL和100μg/m L JBKL作用后磷酸化ERK表達量較溶媒對照組減少。而在濃度100μg/mL,JBKL顯著抑制p38磷酸化表達水平,提示JBKL可能通過作用于ERK-MAPK和p38-MAPK信號通路來發揮抑制T細胞活化的作用。

圖2 蠲痹顆粒醇提物(JBKL)對Anti-CD3 m Ab誘導
3 討論
T細胞介導的免疫反應損傷是導致RA等系統性自身免疫病發病的主要原因之一[1,8]。臨床上大多數RA患者的病程較長,主要以關節的免疫損傷為特征,并且在病原生物的持續感染下,病變可從小關節波及到大關節,最終引起嚴重的炎癥損害[8]。但該病病因和發生機制目前尚未闡明。T淋巴細胞作為適應性免疫應答的主要效應細胞,其不同亞群發揮著不同的免疫效應,主要誘導分泌各類細胞因子或效應分子,從而參與調節體液免疫應答、炎癥反應和自身免疫病等過程。因此,類風濕性關節炎的發生與針對自身抗原的T淋巴細胞異常活化增殖有關[9-10]。
前期研究顯示,蠲痹顆粒能有效抑制DNFB引發的遲發型超敏反應(DTH)。DNFB能夠刺激T細胞活化,釋放大量的細胞因子和炎癥介質,造成炎性損傷。基于此,我們進一步研究發現蠲痹顆粒能顯著改善膠原誘導性小鼠關節炎,維持Th17/Treg細胞平衡,減少致炎因子IL-17A分泌量,促進免疫抑制因子IL-10的釋放,動物模型實驗證明給予蠲痹顆粒后能阻礙T細胞異常活化而發生的免疫應答,所以蠲痹顆粒治療RA的免疫調節機制是值得探索的。因此,本實驗采用ConA、Anti-CD3 mAb和LPS誘導脾臟T、B淋巴細胞增殖的體外模型,并加入JBKL;MTT結果顯示,T、B細胞增殖活性在JBKL干預后減弱。且細胞毒性實驗結果表明JBKL對正常脾淋巴細胞無明顯細胞毒性作用,排除了JBKL抑制淋巴細胞增殖作用依賴于細胞毒性的可能,與前期體內結果一致,進而證實蠲痹顆粒具有顯著免疫抑制活性,這可能是蠲痹顆粒治療RA主要的依據之一[1,4,11]。
白細胞分化抗原3(CD3)是一類分布于細胞膜表面的蛋白質,CD3和 TCR肽鏈(α、β、γ)以非共價鍵相互連接,形成CD3-TCR復合體,在抗原刺激下使T細胞活化[12]。大量研究證實Anti-CD3 mAb可模擬抗原刺激直接作用于CD3,介導T細胞快速活化增殖,促進Th1、Th17、Treg等T細胞亞群分泌細胞因子和炎癥介質釋放[13]。結果表明,JBKL 30μg/mL顯著降低了IFN-γ、IL-17A和IL-6的水平,顯著提高了負調控因子IL-10的水平。IFN-γ除了激活巨噬細胞發揮抗腫瘤效應和促進炎癥反應外,還有免疫調節作用[14]。由Th17細胞分泌的IL-17A有較強的促炎癥作用,在RA的啟動與進展中發揮關鍵作用[15-17]。另外,Th17細胞在IL-6刺激下會發生增殖和分化[18],IL-6還可以活化B淋巴細胞,發揮以抗體為主導的特異性體液免疫效應。由Treg細胞分泌的IL-10是維持免疫耐受的關鍵因子,以防止自生免疫性疾病發生。課題組前期在膠原誘導性關節炎模型中發現JBKL能調控Th17細胞與Treg細胞平衡,下調促炎因子IL-17A表達[3]。本研究在體外細胞因子表達角度進一步證實JBKL能調控Th17/Treg細胞平衡,這可能是蠲痹顆粒治療類風濕關節炎的免疫學機制之一。
在RA發病時,MAPK信號通路是參與TCR信號轉導的重要途徑。T細胞激活信號通過多種途徑轉導入細胞核,其中MAPK信號通路是參與TCR信號轉導的重要途徑。MAPK的3種主要成分:ERK、JNK以及P38均參與T細胞激活。通過激活MAPK信號通路釋放多種核轉錄因子,參與T細胞免疫應答過程[19]。基于此,我們進一步應用Western blot檢測JBKL對MAPK信號通路中相關蛋白的影響。結果表明,JBKL給藥后磷酸化ERK和p38表達降低,提示抑制MAPK信號通路,可能是蠲痹顆粒免疫抑制作用及治療類風濕關節炎的分子機制之一。
綜上所述,本實驗結果證實了JBKL通過調控MAPK信號轉導通路中p38和ERK蛋白磷酸化,從而抑制T細胞活化增殖和促炎因子分泌。本研究可為深入探究中藥復方蠲痹顆粒治療類風濕關節炎的作用機制提供科學基礎。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
