單核細(xì)胞與類風(fēng)濕關(guān)節(jié)炎①
2022-11-16 03:17:20陶夢情胡英豪王丹丹皖南醫(yī)學(xué)院弋磯山醫(yī)院中醫(yī)科蕪湖241000
中國免疫學(xué)雜志 2022年18期
陶夢情 胡英豪 王丹丹 左 堅(jiān)(皖南醫(yī)學(xué)院弋磯山醫(yī)院中醫(yī)科,蕪湖 241000)
類風(fēng)濕關(guān)節(jié)炎(rheumatoid arthritis,RA)是最為常見的自身免疫性疾病,全球總發(fā)病率高達(dá)1%,患者群體規(guī)模龐大[1-2]。RA以關(guān)節(jié)滑膜炎為主要病理特征。正常條件下,滑膜腔是一個(gè)潤滑的無菌空間,細(xì)胞分布稀少,主要由滑膜組織內(nèi)遷的巨噬細(xì)胞構(gòu)成。RA條件下,滑膜組織增生肥大,且伴隨大量炎癥細(xì)胞浸潤;聚集的免疫細(xì)胞通過促進(jìn)滑膜成纖維細(xì)胞增殖與破骨細(xì)胞激活直接參與滑膜炎發(fā)展進(jìn)程[3-4]。異常高表達(dá)的自身抗原被自身反應(yīng)性抗原遞呈細(xì)胞(antigen presenting cell,APC)識別(主要指固有免疫成員,包括單核-巨噬細(xì)胞,樹突狀細(xì)胞等),啟動(dòng)適應(yīng)性免疫應(yīng)答,激活CD4+T細(xì)胞增殖與分化,加速IL-17和IFN-γ等細(xì)胞因子分泌[5];Th1和Th17細(xì)胞依賴旁分泌途徑一方面刺激固有免疫系統(tǒng)合成促炎細(xì)胞因子,另一方面誘導(dǎo)B細(xì)胞向漿細(xì)胞分化,上調(diào)自身抗體合成率。隨后,自身抗體-抗原免疫復(fù)合物作用于特定免疫細(xì)胞表面的Fcγ受體,進(jìn)一步放大單核-巨噬細(xì)胞的促炎功能[6]。上述促炎免疫環(huán)境中,血管內(nèi)皮細(xì)胞、軟骨細(xì)胞、破骨細(xì)胞功能明顯改變,為血管新生、軟骨損傷、破骨增強(qiáng)等病理反應(yīng)發(fā)生創(chuàng)造了條件。
根據(jù)經(jīng)典理論模型,CD4+T細(xì)胞具有廣泛的病理活性,主導(dǎo)RA整體疾病進(jìn)程,在相關(guān)研究中備受關(guān)注,而以單核-巨噬細(xì)胞為代表的固有免疫系統(tǒng)被長期忽視。關(guān)節(jié)定植巨噬細(xì)胞的促炎及組織破壞功能得以較好地闡明,而單核細(xì)胞有關(guān)研究略顯不足。但單核細(xì)胞在RA發(fā)生、發(fā)展中同樣發(fā)揮關(guān)鍵作用(圖1):具備良好分化潛能,是樹突狀細(xì)胞、巨噬細(xì)胞、破骨細(xì)胞等RA病理效應(yīng)細(xì)胞的重要前體來源;具備強(qiáng)大外分泌功能,其典型產(chǎn)物包括細(xì)胞因子、趨化因子、生長因子、基質(zhì)金屬蛋白酶等,從而有效調(diào)控炎癥與骨代謝[7-9];可作為APC激活T細(xì)胞。因此,充分闡明RA條件下單核細(xì)胞的激活因素及調(diào)控方式,有利于加深RA病理機(jī)制理解,開發(fā)創(chuàng)新性RA靶向治療方案。
1 單核細(xì)胞概況
人體中單核細(xì)胞占外周白細(xì)胞的3%~8%,是體積最大的細(xì)胞。單核細(xì)胞主要來源于CD34陽性造血干細(xì)胞(hematopoietic stem cell,HSC),在胎兒肝臟和成年人骨髓中,受多種集落刺激因子刺激,以集落刺激因子-1受體(colony-stimulating factor-1 receptor,CSF1R)依賴性方式首先分化為粒細(xì)胞-單核細(xì)胞前體或單核-樹突狀細(xì)胞前體,這些前體細(xì)胞繼續(xù)增殖并分化為原始單核細(xì)胞進(jìn)入血液,主要表達(dá)CD117、CD34、CD33、HLA-DR(MHCⅡ表面受體),CD13呈 弱 表 達(dá)[10];分 化 為 幼 稚 階 段 時(shí),CD33、CD13、HLA-DR表達(dá)逐漸增強(qiáng),同時(shí)開始表達(dá)CD64、CD11c、CD11b;進(jìn)入成熟階段后,CD14、CD15等標(biāo)志物開始表達(dá)[11]。血液中的單核細(xì)胞在骨髓和組織間遷移,并在組織內(nèi)轉(zhuǎn)變?yōu)槌墒炀奘杉?xì)胞,循環(huán)周期為1~5 d(圖1)。
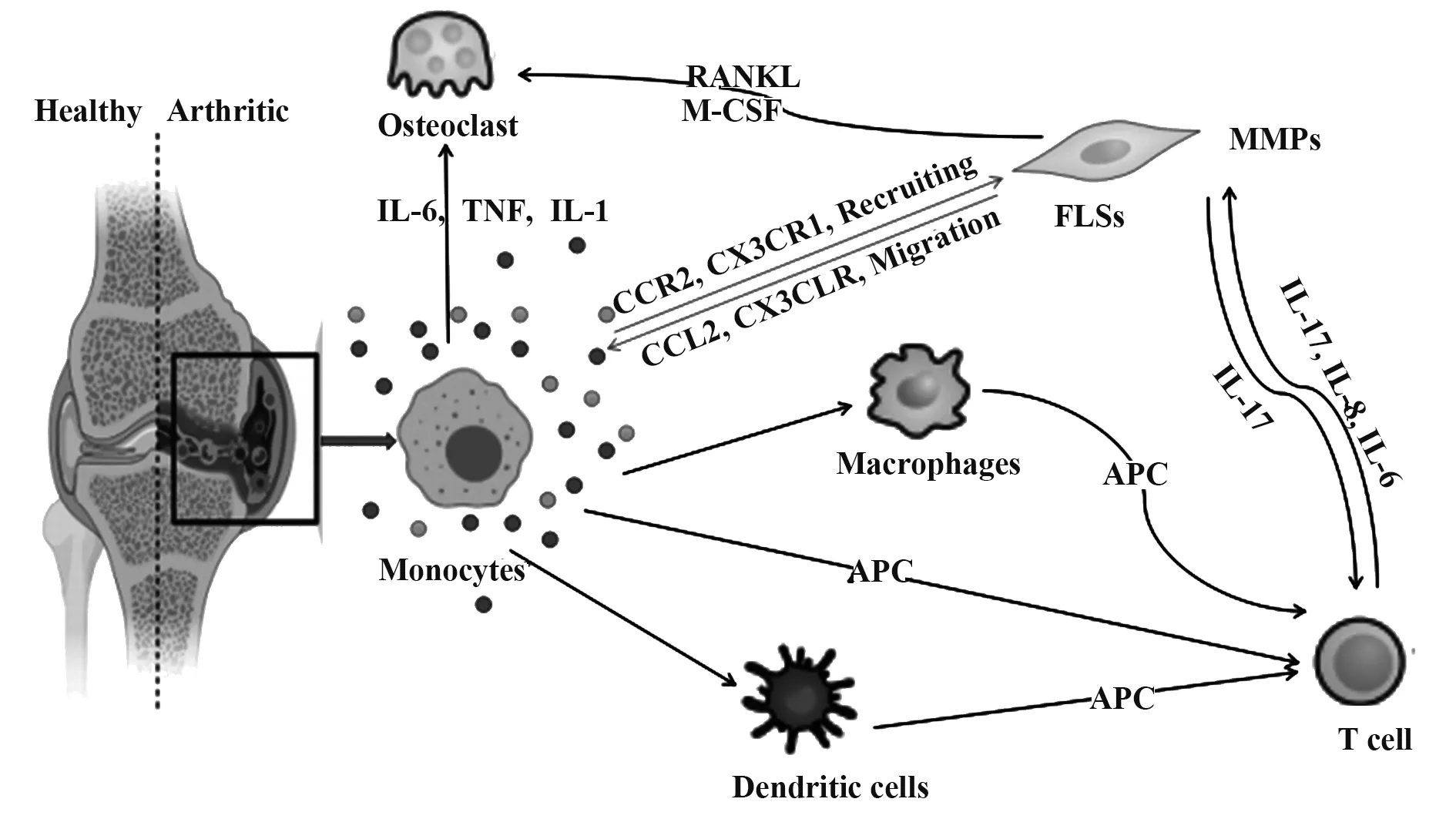
圖1 單核細(xì)胞對RA的推動(dòng)作用示意圖Fig.1 Schematic diagram of promoting effect of monocytes on RA
單核細(xì)胞在功能表型和表觀遺傳學(xué)上均具有高度可塑性。靜息狀態(tài)下,單核細(xì)胞通過高內(nèi)皮小靜脈從血液進(jìn)入淋巴結(jié),或通過傳入淋巴管從組織進(jìn)入淋巴結(jié)。非淋巴組織中的單核細(xì)胞在正常生理?xiàng)l件下一般具有3個(gè)去向:①維持其自身特性并最終凋亡;②CCR7表達(dá)上調(diào),遷移至淋巴結(jié);③在組織巨噬細(xì)胞生態(tài)位有空位的環(huán)境下分化為巨噬細(xì)胞。炎癥環(huán)境中,單核細(xì)胞向組織和淋巴結(jié)中大量浸潤,在微生物病原體存在下優(yōu)先分化為分泌TNF-α和iNOS的促炎樹突狀細(xì)胞或組織駐留巨噬細(xì)胞[12]。
以人類為例,單核細(xì)胞可根據(jù)脂多糖(lipopolysaccharide,LPS)輔助受體CD14和清道夫受體CD16(FcγRⅢ)分布狀態(tài)分為3個(gè)亞類:CD14++CD16-經(jīng)典型(classical monocytes,CMs)、CD14+CD16+中間型(intermediate monocytes,IMs)、CD14-CD16++非經(jīng)典型(non-classical monocytes,NCMs)。CMs占健康人外周血單核細(xì)胞的90%,可迅速被招募至炎癥部位,發(fā)揮吞噬作用,但無典型炎癥特征,高表達(dá)CCR2[13-14];NCMs通常被稱為巡邏單核細(xì)胞,在激活后可表現(xiàn)抗原呈遞及“炎癥”特征,低表達(dá)CCR2,高表達(dá)CX3CR1;IMs是兼具吞噬和炎癥功能的過渡性單核細(xì)胞,表達(dá)CCR2和CX3CR1,同時(shí)高表達(dá)HLADR、CD163及CCR5等[15]。根據(jù)趨化因子受體構(gòu)成差異,其他動(dòng)物來源單核細(xì)胞也可類似地分為經(jīng)典與非經(jīng)典兩種亞型。但由于表面標(biāo)志物種屬特異性,其流式表型判定標(biāo)準(zhǔn)有別于人類。小鼠中,經(jīng)典型和非經(jīng)典型單核細(xì)胞分別表征為Ly6Chi和Ly6Clow[16];大鼠中,兩者分別對應(yīng)CD172a+CD43-和CD172a+CD43+細(xì)胞亞群(表1)[17]。值得注意的是,不同于人類,多數(shù)研究表明鼠類CMs而非NCMs具有典型促炎功能[16-17]。但該結(jié)論仍存在爭議,因?yàn)樯鲜鰟?dòng)物來源NCMs同樣具備向M1型巨噬細(xì)胞分化的潛能。進(jìn)一步表明單核細(xì)胞具有高度可塑性,其病理生理功能復(fù)雜。
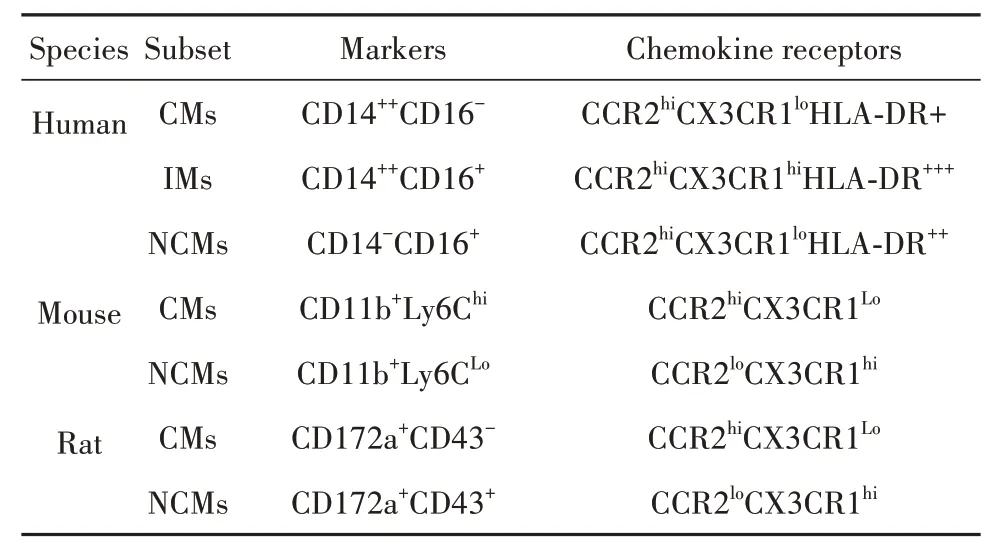
表1 單核細(xì)胞在人類及鼠類動(dòng)物的分群Tab.1 Subsets of monocytes in humans and murines
2 單核細(xì)胞推動(dòng)RA進(jìn)展
與健康人群相比,RA患者外周血CMs比例降低,而CD16+單核細(xì)胞亞群增加(含NCMs及IMs),且該類細(xì)胞占比與疾病活動(dòng)度呈正相關(guān)[18-20]。同時(shí),單核細(xì)胞的其他相關(guān)表面標(biāo)志物構(gòu)成也發(fā)生較大改變。相比于健康人群,RA早期患者體內(nèi)單核細(xì)胞,尤其是NCMs表面CD11c表達(dá)增加[21]。相似地,CD11b表達(dá)也得以上調(diào),且這一現(xiàn)象與RA關(guān)聯(lián)性受體CCR2和CX3CR1表達(dá)改變同步發(fā)生。考慮到上述趨化因子受體與單核細(xì)胞向炎癥性巨噬細(xì)胞和破骨細(xì)胞定向分化密切相關(guān),CD11b在RA患者中的異常高表達(dá)可視為RA特異性單核細(xì)胞的重要鑒定依據(jù)及疾病發(fā)展階段的輔助診斷指標(biāo)[22]。與此對應(yīng),有效的抗風(fēng)濕治療對單核細(xì)胞亞群分布具有恢復(fù)性調(diào)控作用。如經(jīng)甲氨蝶呤(methotrexate,MTX)治療后,隨著C-反應(yīng)蛋白水平及其他炎癥指標(biāo)下調(diào),RA患者外周血IMs減少,而CMs增多[19]。
如上所述,病理?xiàng)l件下CD16+單核細(xì)胞具有獨(dú)特的受體表達(dá)譜,對RA發(fā)展具有推動(dòng)作用。究其原因,從宏觀層面而言,由于STAT1通路下調(diào),RA來源單核細(xì)胞大量分泌TNF-α[23]。TNF-α具有強(qiáng)大的促炎免疫活性,其過度合成是RA患者體內(nèi)細(xì)胞因子網(wǎng)絡(luò)紊亂的主要特征,與疾病進(jìn)展直接相關(guān);靶向阻斷TNF-α已被證明是RA治療的有效手段。RA患者單核細(xì)胞來源TNF-α表達(dá)不僅通過其自身下游受體途徑直接誘發(fā)病理性炎癥反應(yīng),同時(shí)可基于旁分泌途徑促進(jìn)其自身甚至其他免疫細(xì)胞分泌IL-1、IL-17等多種RA關(guān)鍵細(xì)胞因子,誘導(dǎo)T細(xì)胞異常分化[24]。
有研究進(jìn)一步從亞器官角度初步闡明了上述細(xì)胞的病理功能,RA患者單核細(xì)胞周轉(zhuǎn)率提高,在血液循環(huán)時(shí)間減少,具有顯著關(guān)節(jié)聚集傾向[25]。這種現(xiàn)象與RA關(guān)節(jié)免疫環(huán)境及單核細(xì)胞受體表達(dá)譜特征有關(guān):缺氧微環(huán)境中滑膜細(xì)胞高表達(dá)基質(zhì)細(xì)胞衍生因子SDF-1,大量招募CXCR4+單核細(xì)胞,并在局部分化為巨噬細(xì)胞[26]。導(dǎo)致RA患者滑膜中以CD16+亞群為主的單核細(xì)胞數(shù)幾乎是外周血的4倍[21]。上述聚集現(xiàn)象與內(nèi)皮細(xì)胞黏附分子表達(dá)增加也密切相關(guān)。定植于滑膜的單核細(xì)胞繼而有效改變局部免疫環(huán)境。首先,單核-巨噬細(xì)胞為B細(xì)胞長期存活提供環(huán)境,促進(jìn)其產(chǎn)生RA診斷標(biāo)志物抗環(huán)瓜氨酸抗體(anti-citrullinated protein antibodies,ACPA)[27]。ACCP陽性患者炎癥/抗炎單核細(xì)胞升高,分泌更多炎癥因子;而加劇的免疫紊亂又促進(jìn)CD14+單核細(xì)胞分化為破骨細(xì)胞[28-29]。這一反饋機(jī)制表明單核細(xì)胞是RA關(guān)節(jié)免疫穩(wěn)態(tài)破壞及骨損傷的重要推手。另一方面,IMs在滑膜中占主導(dǎo)地位,表達(dá)較多的HLA-DR和CD80/CD86,為CD4+T細(xì)胞激活提供第一和第二信號[24,30-31];并分泌更多的TNF-α、IL-6和IL-1β促進(jìn)IL-17分化[32-33]。T細(xì)胞激活引發(fā)的炎癥因子外溢又直接反饋性作用于單核細(xì)胞,并促進(jìn)CD16與IgG免疫復(fù)合物生成,推動(dòng)其向破骨細(xì)胞分化[34]。
3 RA單核細(xì)胞分化異常調(diào)控
3.1 經(jīng)典免疫信號因素Toll樣受體(Toll-like receptors,TLRs)是固有免疫細(xì)胞特征性模式識別受體,也是單核細(xì)胞主要的功能性免疫分子,通過識別宿主的細(xì)胞碎片和微生物成分激活防御性促炎免疫應(yīng)答。單核細(xì)胞的TLRs相關(guān)信號異常直接驅(qū)動(dòng)RA患者關(guān)節(jié)病理反應(yīng)發(fā)展。作為這一結(jié)論的直接證據(jù),RA患者外周血中CD14+單核細(xì)胞高表達(dá)多種類型的TLRs,包括TLR2、TLR5、TLR7和TLR9等;而TLR5/7 mRNA表達(dá)與RA關(guān)節(jié)炎評分DAS28直接相關(guān)[35-36]。功能方面,TLR1/2/5/9激活可促進(jìn)CMs等各型單核細(xì)胞分泌IL-6和TNF-α[37];TLR5/7上調(diào)可導(dǎo)致CD14+細(xì)胞分化為破骨細(xì)胞[38-39]。該類受體的信號介導(dǎo)功能是上述病理反應(yīng)的重要基礎(chǔ)。大部分TLRs結(jié)合配體后通過MyD88依賴性方式激活下游促炎通路,主要包括核因子-κB(nuclear factor kappa-B,NF-κB)和絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信號通路,進(jìn)而誘導(dǎo)單核細(xì)胞來源TNF-α和IL-6等促炎細(xì)胞因子分泌,促進(jìn)RA進(jìn)展[40]。
3.2 表觀遺傳因素 表觀遺傳因素是不容忽視的RA誘發(fā)因素。以甲基化為例,RODRíGUEZ-UBREVA等[41]發(fā)現(xiàn)活動(dòng)期RA患者病情與循環(huán)單核細(xì)胞DNA總體甲基化水平密切相關(guān);而炎癥性關(guān)節(jié)炎來源單核細(xì)胞DNA甲基化譜變化較正常人群更為顯著[42]。MOK等[43]進(jìn)一步發(fā)現(xiàn)單核細(xì)胞內(nèi)CYP2E1啟動(dòng)子低甲基化是RA活動(dòng)度的獨(dú)立預(yù)測因素;而CCL2啟動(dòng)子區(qū)H3K4、H3K27、H3K36和H3K79的三甲基化亦可推動(dòng)RA進(jìn)展[44]。非編碼RNA是DNA修飾外的另一重要表觀遺傳調(diào)控因素。RA患者中可見單核細(xì) 胞 內(nèi)lncRNA NTT/PBOV1軸 異 常 上 調(diào)[45]。lnc-RNA NTT是單核細(xì)胞炎癥的調(diào)節(jié)因子;而PBOV1在THP-1細(xì)胞中過表達(dá)可導(dǎo)致單核細(xì)胞G1期阻滯,分化為巨噬細(xì)胞[46]。兩者協(xié)同促進(jìn)了RA單核細(xì)胞向促炎細(xì)胞類型分化成熟。相比于以上領(lǐng)域的有限認(rèn)知,miRNAs在RA中的病理意義研究較為深刻,下面進(jìn)行重點(diǎn)探討。
RA患者外周血和滑膜中CD14+單核細(xì)胞高表達(dá)miR-155,且與疾病活動(dòng)度相關(guān)[47]。現(xiàn)有證據(jù)表明,miR-155可通過調(diào)節(jié)單核細(xì)胞極化、凋亡與分泌等功能在RA中發(fā)揮重要作用[47-50]。涉及的核心機(jī)制在于miR-155可選擇性結(jié)合細(xì)胞因子信號傳導(dǎo)抑制因子1(suppressor of cytokine signaling 1,SOCS1)的3'UTR區(qū)域,通過抑制其轉(zhuǎn)錄水平間接促進(jìn)促炎細(xì)胞因子分泌[48-49]。miR-33和miR-125a-5p也被證明與RA進(jìn)展有關(guān)。RA單核細(xì)胞中miR-33可能通過抑制線粒體耗氧率和誘導(dǎo)胞內(nèi)ROS積累刺激NLRP3炎癥小體信號[51]。miR-125a-5p異常高表達(dá)主要發(fā)現(xiàn)于幼年RA患者[52]。雖然其直接病理意義尚未闡明,但已明確這一現(xiàn)象與TLRs/NF-κB興奮有關(guān),而IL-4刺激則抑制其表達(dá)[53-54]。因此,miR-125a-5p總體有利于單核-巨噬細(xì)胞促炎表型形成。miR-129-3p和miR-518a-5p對單核細(xì)胞遷移等病理生理功能也表現(xiàn)出促進(jìn)作用[55-56]。
另一方面,miR-146a等對RA可能存在保護(hù)作用。miR-146a是TLRs信號通路主要的負(fù)調(diào)控因子;IMs和NCMs等單核細(xì)胞可能代償性上調(diào)miR-146a表達(dá),抑 制TRAF6和IRAK1表 達(dá),進(jìn)而 抑制TLRs/NF-κB活化,通過負(fù)反饋機(jī)制限制炎癥過度發(fā)展[57]。因此,膠原誘導(dǎo)關(guān)節(jié)炎(collagen-induced arthritis,CIA)小鼠中Ly6Chi單核細(xì)胞過表達(dá)miR-146a可緩解骨損傷[58]。但miR-146a-5p在銀屑病關(guān)節(jié)炎患者外周血CD14+單核細(xì)胞中的表達(dá)可誘導(dǎo)破骨細(xì)胞活化、骨吸收[59]。表明其病理生理功能仍需進(jìn)一步研究。臨床樣本分析發(fā)現(xiàn),ACPA陽性RA患者單核細(xì)胞出現(xiàn)miR-let7a特異性表達(dá)降低[60-61]。miRlet7a可有效抑制IL-1、IL-6表達(dá),導(dǎo)致RA患者炎癥反應(yīng)持續(xù)發(fā)展[61]。
3.3 能量代謝因素 能量代謝對免疫細(xì)胞分化、成熟甚至功能發(fā)揮關(guān)鍵作用。RA活動(dòng)期患者CD14+單核細(xì)胞耗氧量及線粒體代謝水平偏高。SHIRAI等[62]發(fā)現(xiàn),慢性炎癥中致病性單核細(xì)胞和巨噬細(xì)胞葡萄糖過度利用,并共享代謝線路。針對性機(jī)制研究發(fā)現(xiàn),RA來源單核-巨噬細(xì)胞中糖原合成激 酶-3β(glycogen synthase kinase 3β,GSK-3β)失活,線粒體電子傳遞鏈活性增強(qiáng),氧化磷酸化強(qiáng)化,ATP及ROS生成增加[63]。糖代謝狀態(tài)改變誘發(fā)或伴隨顯著免疫學(xué)功能變化。一方面,LPS等促炎因素刺激將磷酸化單核細(xì)胞內(nèi)mTOR,繼而誘發(fā)同步的葡萄糖攝取/酵解上調(diào)及以CD11b+為特征的炎癥表型獲得[64]。RA背景下,mTOR激活可能與氨基酸轉(zhuǎn)運(yùn)蛋白SLC7A5控制的亮氨酸內(nèi)流有關(guān)。RA患者單核細(xì)胞中,SLC7A5表達(dá)顯著升高,糖酵解代謝隨之上調(diào);阻斷SLC7A5將導(dǎo)致mTOR信號抑制,IL-1β產(chǎn)率降低[65]。另一方面,葡萄糖過度利用將導(dǎo)致ROS蓄積,從而促進(jìn)胞質(zhì)中丙酮酸激酶M2(pyruvate kinase M2,PKM2)二聚化及入核轉(zhuǎn)移。后者作為蛋白激酶直接激活核轉(zhuǎn)錄因子STAT3,促進(jìn)IL-6和IL-1β等轉(zhuǎn)錄表達(dá);同時(shí)通過干預(yù)HIF-1α信號誘導(dǎo)IL-1β生成[66]。除糖代謝外,脂代謝模式也發(fā)生了同步改變。受缺氧環(huán)境中滑膜細(xì)胞誘導(dǎo),RA患者關(guān)節(jié)處單核細(xì)胞中肉堿代謝物增加,促進(jìn)CCL20釋放,單核細(xì)胞向破骨細(xì)胞分化的潛能及對Th17細(xì)胞的募集能力增強(qiáng)[67]。此外,關(guān)節(jié)炎模型小鼠單核細(xì)胞膽固醇外排基因Abca1、Apoe表達(dá)下調(diào),對干細(xì)胞分化和髓外髓細(xì)胞生成發(fā)育與增殖具有深遠(yuǎn)影響[68]。
4 靶向單核細(xì)胞的RA治療
4.1 細(xì)胞因子阻斷 促炎細(xì)胞因子是RA發(fā)生發(fā)展的核心驅(qū)動(dòng)因素。其中大量關(guān)鍵細(xì)胞因子如IL-1、IL-6、TNF-α的主要來源包括炎癥單核細(xì)胞,而另一些細(xì)胞因子則可干預(yù)單核細(xì)胞分化和功能,從而影響后者免疫功能。因此,多種經(jīng)典RA治療藥物具有單核細(xì)胞分泌干預(yù)活性。如MTX可有效抑制RA患者單核細(xì)胞中細(xì)胞因子產(chǎn)生及膜受體表達(dá),首診患者外周血單核細(xì)胞總數(shù)、CMs、IMs偏高,對MTX的治療敏感性相對較低[69]。如青風(fēng)藤堿已被證明可降低RA患者外周IMs比例,減少炎癥細(xì)胞因子如粒細(xì)胞巨噬細(xì)胞集落刺激因子(granulocyte macrophage colony stimulating factor,GM-CSF)、TNF-α、IL-1β、CXCL1、巨噬細(xì)胞集落刺激因子(macrophage colony stimulating factor,M-CSF)等合成,進(jìn)而緩解單核-巨噬細(xì)胞主導(dǎo)的局部炎癥與組織破壞[70]。
上述現(xiàn)象對靶向制劑開發(fā)極具啟發(fā)。已明確在TNF-α等刺激下,CD11b+單核細(xì)胞更易分化為破骨細(xì)胞,且促炎免疫能力增強(qiáng)。與這一結(jié)論對應(yīng),TNF抑制劑(tumor necrosis factor inhibitors,TNFi)可使循環(huán)中CMs亞群顯著減少,并通過下調(diào)其表面CXCR4、CCR2表達(dá)抑制其免疫活性[71];單核細(xì)胞干預(yù)可進(jìn)一步實(shí)現(xiàn)Treg分化增強(qiáng)及破骨細(xì)胞功能抑制[72]。此外,TNFi還可誘導(dǎo)幼稚單核細(xì)胞向NCMs分化成熟,避免高致病性IMs過度增加;下調(diào)NCMs中CD11b表達(dá),增加其穿越內(nèi)皮的難度,導(dǎo)致炎癥部位單核細(xì)胞浸潤減少[73];此外,作用于跨模型TNF-α可進(jìn)一步強(qiáng)化對單核細(xì)胞炎癥免疫功能的影響,并誘導(dǎo)其凋亡[74]。
其他單核細(xì)胞相關(guān)細(xì)胞因子的選擇性阻斷表現(xiàn)出類似治療潛力。靶向IL-6阻斷可誘導(dǎo)單核細(xì)胞凋亡,從而間接促進(jìn)Treg分化,提高其分布比例。GM-CSF可刺激單核細(xì)胞表達(dá)HLA-DR和CD86,合成TNF-α和IL-1β,從而具備啟動(dòng)適應(yīng)性免疫的能力;GM-CSF信號阻斷原理上可逆轉(zhuǎn)上述過程,同時(shí)可刺激單核細(xì)胞分泌趨化因子CXCL-11,間接抑制T細(xì)胞增殖[75]。M-CSF也可通過影響單核細(xì)胞分化干預(yù)RA進(jìn)程;基于兩例活動(dòng)期RA患者的臨床試驗(yàn)表明,M-CSF拮抗抗體可降低外周血IMs和NCMs分布比例,改善臨床癥狀[76]。
4.2 表面抗原拮抗 單核細(xì)胞表面標(biāo)志物CD11b在髓系細(xì)胞分化的過程中發(fā)揮重要作用。RA患者體內(nèi)CD11b表達(dá)顯著高于健康對照組;糖皮質(zhì)激素有效治療后其表達(dá)趨于下降[77]。這一現(xiàn)象初步提示CD11b可能是RA的治療靶點(diǎn)。抗CD11b單克隆抗體可減少CIA小鼠體內(nèi)單核細(xì)胞分布,抑制關(guān)節(jié)炎癥部位對單核細(xì)胞的招募,及相關(guān)破骨細(xì)胞分化。雷帕霉素通過作用于其受體mTOR能夠調(diào)控前體單核細(xì)胞內(nèi)STAT5-IRF8信號通路,從而下調(diào)表面CD115表達(dá)。鑒于CD115可驅(qū)動(dòng)單核細(xì)胞分化為巨噬細(xì)胞或破骨細(xì)胞,上述變化最終將有效緩解RA滑膜炎癥和關(guān)節(jié)破壞[78]。
4.3 信號通路調(diào)控Janus激酶(Janus kinase,JAK)作為重要的酪氨酸激酶,銜接了諸多細(xì)胞因子受體與下游信號轉(zhuǎn)導(dǎo)通路;同時(shí)JAK-STAT途徑又參與了細(xì)胞因子合成過程,被確定為RA炎癥性病變的關(guān)鍵信號傳導(dǎo)途徑之一。Peficitinib與Baricitinib可通過抑制滑膜成纖維樣滑膜細(xì)胞JAK-STAT通路,抑制多種促炎細(xì)胞因子分泌,阻斷單核細(xì)胞向炎癥部位趨化及后續(xù)炎癥分化過程[79-80]。FcγR通過胞內(nèi)免疫受體酪氨酸活化基序激活酪氨酸激酶,啟動(dòng)后續(xù)磷酸偶聯(lián)級聯(lián)反應(yīng),激活單核-巨噬細(xì)胞免疫功能;布魯頓酪氨酸激酶(Bruton's tyrosine kinase,BTK)通過結(jié)合FcγR介導(dǎo)免疫復(fù)合物免疫調(diào)控活性。此外,BTK還參與單核細(xì)胞來源破骨細(xì)胞分化過程,其選擇性抑制劑HM71224可有效抑制BTK介導(dǎo)的單核細(xì)胞激活,并抑制破骨細(xì)胞主導(dǎo)的骨結(jié)構(gòu)破壞,有效治療實(shí)驗(yàn)性小鼠關(guān)節(jié)炎[81]。已批準(zhǔn)和正在開發(fā)的作用于單核細(xì)胞的RA治療藥物見表2。

表2 已批準(zhǔn)和正在開發(fā)的作用于單核細(xì)胞的RA治療藥物Tab.2 Drugs approved and under development applies to monocytes for RA treatment
5 展望
現(xiàn)有常規(guī)RA治療藥物雖能緩解癥狀,但無法遏制疾病進(jìn)程,且毒副作用明顯。近年興起的生物療法作用機(jī)制和靶點(diǎn)明確,短期臨床效果顯著,但存在安全風(fēng)險(xiǎn)。因此闡明RA發(fā)生發(fā)展過程不同階段免疫細(xì)胞的動(dòng)態(tài)變化具有重要意義。識別主導(dǎo)RA單核細(xì)胞病理變化的分子機(jī)制,有利于開發(fā)RA精準(zhǔn)治療新方案。早期診斷和適當(dāng)治療可迅速控制RA進(jìn)展,顯著改善肢體功能和關(guān)節(jié)結(jié)構(gòu)。本課題組近期研究發(fā)現(xiàn),煙酰胺磷酸核糖轉(zhuǎn)移酶(nicotinamide phosphoribosyltransferase,NAMPT)介導(dǎo)的單核細(xì)胞炎癥極化對早期RA發(fā)展具有關(guān)鍵推動(dòng)作用。考慮到NAMPT作為炎癥表型單核-巨噬細(xì)胞共有的特征性糖代謝重編程核心操控因素,及其靶向抑制對RA的顯著治療潛力,這一發(fā)現(xiàn)表明代謝水平調(diào)控亦可能是實(shí)現(xiàn)單核細(xì)胞靶向治療的策略[82-83]。

- 中國免疫學(xué)雜志的其它文章
- SHP2特性及其變構(gòu)抑制劑SHP099腫瘤免疫治療的研究進(jìn)展
- 調(diào)節(jié)性B細(xì)胞在妊娠合并系統(tǒng)性紅斑狼瘡中的研究進(jìn)展①
- PBL+CBL+TBL混合式教學(xué)模式應(yīng)用于我國基礎(chǔ)醫(yī)學(xué)教學(xué)效果的Meta分析①
- 線上線下混合式教學(xué)在醫(yī)學(xué)免疫學(xué)教學(xué)中的應(yīng)用與反思①
- 貝利尤單抗治療系統(tǒng)性紅斑狼瘡的效果分析①
- 復(fù)方甘草酸苷片聯(lián)合第二代抗組胺藥物治療慢性蕁麻疹有效性及安全性的Meta分析①