五糧液釀造環(huán)境空氣中霉菌的分離鑒定及產酶初探
2022-12-26 06:31:24雷學俊趙東鄭佳喬宗偉劉多濤張霞李智
食品與發(fā)酵工業(yè) 2022年23期
雷學俊,趙東,2,鄭佳*,喬宗偉,2,劉多濤,張霞,李智
1(宜賓五糧液股份有限公司,四川 宜賓,644007)2(中國輕工業(yè)固態(tài)濃香型白酒發(fā)酵重點實驗室,四川 宜賓,644007)
傳統(tǒng)固態(tài)白酒是我國古代勞動人民經驗智慧的結晶。在獨特的地理環(huán)境條件下,固態(tài)白酒獨特的開放式生產,為釀造高品質白酒網羅富集了空氣中微生物,多元的釀造工藝塑造了獨特的釀造微生物菌群結構并賦予了白酒復雜的香型特征。五糧液作為我國傳統(tǒng)固態(tài)白酒的典型代表,是國家地理標志產品,入選首批中歐地理標志產品互認清單產品[1],其原產地宜賓市位于岷江、金沙江、長江合流之處,被譽為萬里長江第一城,海拔500~2 000 m,獨特的中亞熱帶濕潤季風氣候(年均降雨量1 100 mm以上,年均氣溫18 ℃)以及當地獨特的黃黏土為高品質五糧液酒的釀造奠定了物質基礎[2]。正是因為當地獨特的地理環(huán)境條件,因而十分有必要系統(tǒng)研究空氣中微生物菌群的多樣性。
目前空氣微生物研究主要采用自然沉降法和機器采樣法采集樣品,并結合培養(yǎng)基法及非培養(yǎng)基法進行鑒定,解析白酒釀造環(huán)境微生物群落結構及其作用。朱治宇等[3]利用傳統(tǒng)可培養(yǎng)及18S rRNA序列分析法對醬香型白酒釀造車間的墻面、窗臺等環(huán)境灰塵樣品中霉菌種群結構多樣性進行分析,表明Lichtheimia、Mucor、Aspergillus等霉菌屬是醬香白酒釀造環(huán)境優(yōu)勢菌屬。張亞麗[4]采用安德森法結合傳統(tǒng)微生物培養(yǎng)法及現代分子生物學方法系統(tǒng)性分析了茅臺酒生產環(huán)境中空氣細菌和真菌的組成結構。車路萍等[5]利用機器采樣結合磷脂脂肪酸(phospholipid fatty acid,PLFA)圖譜分析法分析了大曲及曲房空氣微生物PLFA種類數,指出曲房空氣中以革蘭氏陰性細菌為優(yōu)勢菌群。陳澤軍等[6]采用自然沉降法結合真菌分類學方法分析了濃香型酒廠空氣中霉菌組成,指出生產車間內霉菌種類數量高于廠區(qū)和廠區(qū)外,Oedocephalum為優(yōu)勢屬。迄今為止,還未見對五糧液釀造環(huán)境空氣中可培養(yǎng)霉菌多樣性及菌株進行系統(tǒng)的研究。
因此,本研究采用空氣浮游菌采集器采集五糧液核心釀造區(qū)域內空氣微生物,結合可培養(yǎng)法及內源轉錄間隔區(qū)(internally transcribed spacer,ITS)序列分析技術,系統(tǒng)分析了五糧液白酒釀造環(huán)境空氣中可培養(yǎng)霉菌種群多樣性,并初步探索分離菌株的產酶功能,為認識了解環(huán)境微生物對五糧液釀造的貢獻提供理論支撐。
1 材料與方法
1.1 樣品采集
樣品地點位于宜賓五糧液股份有限公司主生產基地釀酒車間外圍、攤晾場和窖池區(qū)域等釀酒環(huán)境(28°45′02″~28°52′00″N,104°34′01″~104°39′03″E),采樣時間為2020年5~6月。
采用浮游菌采樣器收集空氣樣本,采樣器距離地面80 cm,將直徑為9 cm的PDA平板置于采樣器內,空氣流量100 L/min,采樣時間1 min;采集結束后,將平板置入無菌塑料封袋中,30 min內運回實驗室進行霉菌分離。每個取樣地點以半徑為2 m的圓上進行3點取樣,每個點平行取樣3份,共計135份樣品。
1.2 樣品采集
葡萄糖(分析純)、蛋白胨、牛肉膏,國藥集團化學試劑有限公司;脫脂牛奶,蒙牛集團股份有限公司;脫氧膽酸鈉,北京奧博星生物技術有限責任公司;氯霉素,廣州賽國生物科技有限公司;Ezup柱式真菌基因組DNA抽提試劑盒,生工生物工程(上海)股份有限公司;FKC-1型浮游菌采樣器,江蘇蘇凈凈化設備有限公司;離心機,Eppendorf公司;光學顯微鏡,日本Olympus公司;生化培養(yǎng)箱,上海一恒科技儀器有限公司;UV-1800紫外可見分光光度計,日本島津公司。
PDA 分離培養(yǎng)基(g/L):馬鈴薯浸粉6.0,葡萄糖20.0,瓊脂20.0,氯霉素1.0,脫氧膽酸鈉2.0,pH 5.6。
產酶活定性培養(yǎng)基[7]:(1)產淀粉酶培養(yǎng)基(g/L):蛋白胨10.0,NaCl 5.0,牛肉膏5.0,可溶性淀粉2.0,瓊脂20.0,pH 4.0。(2)產蛋白酶培養(yǎng)基(g/L):脫脂牛奶20.0,瓊脂20.0,pH 6.5。(3)產纖維素酶培養(yǎng)基(g/L):羧甲基纖維素鈉(CMC-Na) 10.0,KH2PO42.0,MgSO4·7H2O 0.5,NaNO31.0,剛果紅0.1,瓊脂20.0。(4)產果膠酶培養(yǎng)基(g/L):K2HPO41.0,MgSO40.5,NaNO33.0,果膠2.0,FeSO4·7H2O 0.01,瓊脂20.0,pH 5.5。
發(fā)酵培養(yǎng)基[8]:麩皮培養(yǎng)基:麩皮10.0 g,面粉3.0 g,水10.0 mL,混合均勻,121 ℃滅菌20 min。
1.3 菌株分離純化、保藏及形態(tài)描述
將采集樣品后的平板倒置于28 ℃恒溫培養(yǎng)箱3~4 d,待菌落長出后,根據菌落顏色、形態(tài)、大小等表型差異,挑取所有形態(tài)、顏色不同的單菌落,在新平板上進行劃線純化、編號并記錄菌落形態(tài)特征。將純化后的菌株制作凍干備份,置于-80 ℃超低溫冰箱保存。通過觀察培養(yǎng)基上的純培養(yǎng)菌落記錄其菌落形態(tài),通過光學顯微鏡觀察顯微形態(tài)。
1.4 ITS序列PCR擴增及序列測定
采用Ezup柱式真菌基因組DNA抽提試劑盒提取基因組DNA后,采用真菌通用引物ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4 (5′-TCCTCCGCTTATTGATATGC-3′)擴增霉菌的rRNA基因ITS。PCR反應體系參考文獻[9],PCR反應條件為:95 ℃預變性3 min后,進入以下循環(huán):94 ℃變性30 s,55 ℃退火25 s,72 ℃延伸1 min,35個循環(huán);72 ℃延伸5 min。經質量分數2%瓊脂糖凝膠電泳檢測,將PCR擴增產物送至生工生物工程(上海)股份有限公司進行測序,序列提交至NCBI GenBank (https://www.ncbi.nlm.nih.gov/genbank/)進行比對分析。
1.5 序列比對及系統(tǒng)發(fā)育樹的構建
參考文獻[10]略有改動,將獲得的霉菌rRNA基因ITS序列在GenBank 數據庫中進行同源序列分析(BLAST, https://blast.ncbi.nlm.nih.gov/),下載最相似序列。利用MEGA 5.0軟件進行多序列比對、p-distances模型計算進化距離、最后采用鄰接法(neighbor-joining)構建系統(tǒng)發(fā)育樹,并用Bootstrap檢驗進化樹分支聚類的置信度,重復1 000次。
1.6 產酶霉菌的篩選
產淀粉酶、蛋白酶、纖維素酶、果膠酶菌株的篩選均參照文獻[11-13],略有改動。產淀粉酶菌株的篩選:在含有可溶性淀粉的產淀粉酶培養(yǎng)基中,加入盧氏碘液,以菌落周圍出現明顯的水解圈,判斷其水解淀粉的能力,28 ℃培養(yǎng)96 h;產蛋白酶菌株的篩選:在以脫脂牛奶為底物的產蛋白酶培養(yǎng)基中進行蛋白酶活性檢測,以菌落周圍是否產生透明圈來表示蛋白酶活性,28 ℃培養(yǎng)96 h;產纖維素酶的篩選:在含有CMC-Na的產纖維素酶培養(yǎng)基中加入剛果紅染液,以菌落周圍是否產生透明圈來表示纖維素酶活性,28 ℃培養(yǎng)96 h;產果膠酶的篩選:在含有果膠的產果膠酶培養(yǎng)基中加入盧氏碘液,以菌落周圍是否產生透明圈來表示果膠酶活性,28 ℃培養(yǎng)96 h。
1.7 酶活力測定
酶液的制備:將篩選得到透明圈較大的霉菌接種于麩皮培養(yǎng)基中,28 ℃培養(yǎng)24 h后扣瓶,繼續(xù)培養(yǎng)72 h。稱取麩曲5.0 g,加入30 mL磷酸緩沖液,40 ℃下保溫1 h,經5 000 r/min離心10 min去沉淀,所得粗酶液用于后續(xù)酶活力測定。
α-淀粉酶活力和蛋白酶活力[14]、纖維素酶活力[13]、果膠酶活力[11]的測定均參照文獻所述方法進行。
2 結果與分析
2.1 可培養(yǎng)霉菌菌株菌落形態(tài)和結構
采用傳統(tǒng)培養(yǎng)法,挑取平板上大小、顏色以及顯微細胞形態(tài)等方面有差異的菌落。菌落形態(tài)如圖1所示,部分菌株編號為1-17、1-57、1-48、1-62、1-47、1-104、1-83、1-79、1-107和1-45 d,顏色種類豐富,包括黃色、綠色、白色、紫黑色、銹橘色、黑色;菌落直徑15~70 mm。
顯微形態(tài)如圖2所示,霉菌的分生孢子梗呈有隔或無隔,分生孢子形態(tài)有圓形、橢圓形、球形或輻射狀等,分生孢子單生或鏈生;毛霉屬無囊托,有球形孢子囊,內含大量的孢子囊孢子;青霉屬有分生孢子梗和近圓形的分生孢子,整體呈典型的“帚狀枝”形態(tài);曲霉屬分生孢子梗長而粗,頂端產生瓶狀或近球形球囊,附著生長球形分生孢子;Quambalaria無分生孢子梗,菌絲頂端或其短分枝產橢圓形、卵形分生孢子;Cunninghamella無囊軸和囊托,孢子囊梗的分枝頂端形成球形孢子囊。
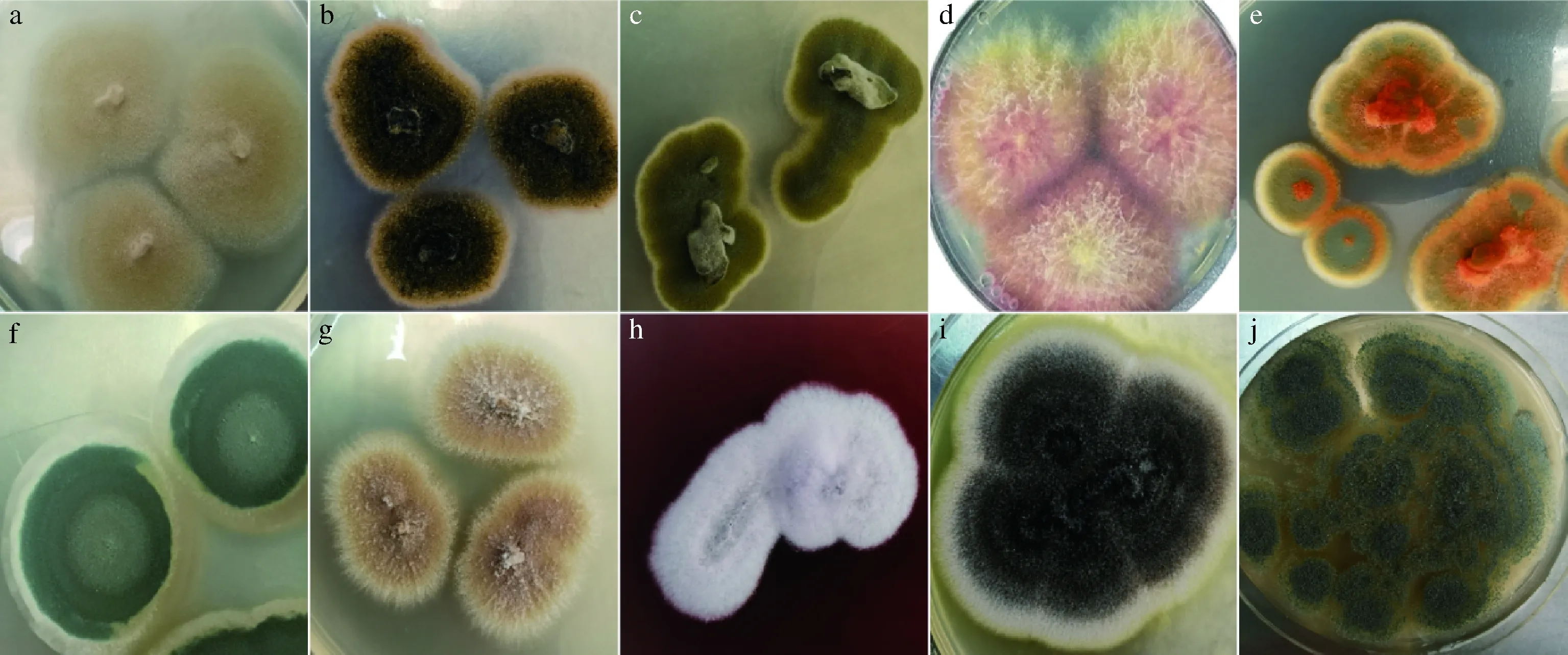
a:1-17;b:1-57;c:1-48;d:1-62;e:1-47;f:1-104;g:1-83;h:1-79;i:1-107;j:1-45 d(下同)圖1 部分霉菌單菌落形態(tài)Fig.1 The single colony morphology of some molds

圖2 部分霉菌的顯微結構Fig.2 Optical microscope observation of cells of some molds
2.2 基于ITS序列系統(tǒng)發(fā)育分析
擴增從3個不同釀酒場地空氣中共得到可培養(yǎng)霉菌的ITS rDNA序列并測序,通過NCBI 中的 BLAST 比對工具在GenBank數據庫中進行同源性對比,其ITS與GenBank中對應菌株的相似性均在99%以上,構建系統(tǒng)發(fā)育樹如圖3所示。66株可培養(yǎng)霉菌分屬3門6綱9目15科18屬41種,曲霉屬(Aspergillus)13個種、青霉屬(Penicillium) 8個種;毛霉屬(Mucor)3個種;木霉屬(Trichoderma)、交鏈孢屬(Alternaria)、枝孢霉屬(Cladosporium)各2個種;黃絲曲霉屬(Talaromyces)、鐮刀菌屬(Fusarium)、Purpureocillium、Pestalotiopsis、帚霉屬(Scopulariopsis)、附球霉屬(Epicoccum)、Didymella、Peyronellaea、Stagonosporopsis、Botryotinia、Sclerotinia和Quambalaria各1個種。其中曲霉屬(Aspergillus)19株和青霉屬(Penicillium)15株占菌株總數的51.52%,表明Aspergillus和Penicillium等屬霉菌是濃香型白酒釀造核心區(qū)域的優(yōu)勢菌屬。
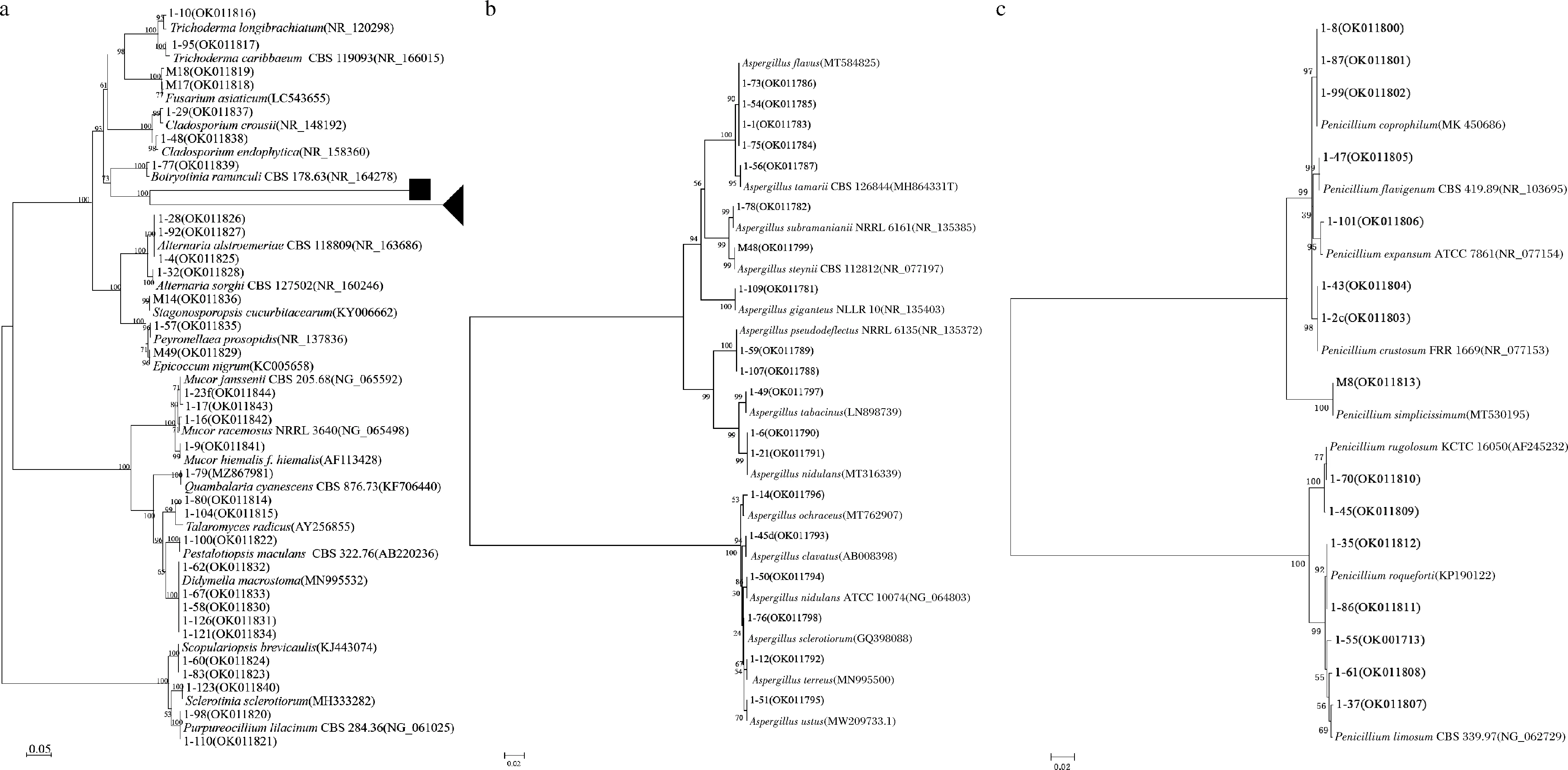
a-所有菌株發(fā)育樹圖;b-a中黑色正方形圖標所代表的曲霉屬菌株發(fā)育樹圖;c-a中黑色三角形圖標所代表的青霉屬菌株發(fā)育樹圖圖3 基于ITS rRNA基因序列構建的釀造環(huán)境空氣中霉菌系統(tǒng)發(fā)育樹Fig.3 Phylogenetic tree based on ITS rRNA gene sequences of the molds isolated from the air of Wuliangye Baijiu-making workshop
2.3 不同位置空氣中可培養(yǎng)霉菌分布
根據釀酒車間的工藝操作流程,車間內大致分為帶有攤晾床的前處理區(qū)域和窖池發(fā)酵區(qū)域。比較車間外環(huán)境,車間內攤晾區(qū)域和窖池區(qū)域的可培養(yǎng)霉菌多樣性分布,各個釀酒場地得到的分布情況結果如圖4所示。從3個釀酒場地空氣中分離的菌株隸屬于18個不同屬。其中,車間外圍空氣中獲得可培養(yǎng)霉菌最多,為28株,分屬2門5綱7目13科16屬;攤晾場共得到26株,分屬2門4綱6目8科9屬;窖池區(qū)域共得到12株,分屬3門4綱5目5科6屬。
在屬水平,對比分析3個不同釀造場地的可培養(yǎng)霉菌相對豐度(圖4-a),發(fā)現不同場地的菌群差異明顯,其中曲霉屬(Aspergillus)、青霉屬(Penicillium)、交鏈孢屬(Alternaria)在3個采樣點中均有檢出。在車間外環(huán)境、攤晾床區(qū)域和窖池發(fā)酵區(qū)域分離得到菌株總數中,曲霉屬菌株數分別占25.00%、26.92%和41.67%,青霉屬菌株數分別占21.43%、23.08%和25.00%,交鏈孢屬菌株數分別占3.57%、7.69%和8.33%;毛霉屬(Mucor)、帚霉屬(Scopulariopsis)是攤晾場和窖池區(qū)域共有分布屬;Botryotinia、Peyronellaea、Epicoccum、Fusarium等8個屬僅在釀造車間外圍分離得到;木霉屬(Trichoderma)、枝孢霉屬(Cladosporium)、黃絲曲霉屬(Talaromyces)僅在攤晾場分離得到;Quambalaria屬僅在窖池區(qū)域分離得到。在種水平(圖4-b),A.flavus和A.nidulans在3個采樣點中均被分離得到。
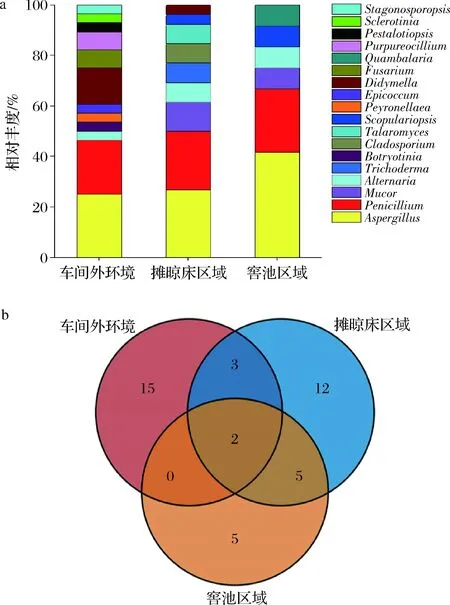
a-屬水平;b-種水平的韋恩圖分布圖4 釀酒場地空氣中霉菌的多樣性Fig.4 Diversity of molds in the air of Wuliangye baijiu-making workshop
2.4 霉菌產酶情況初探
本研究中,66株霉菌產酶特性表明(圖5),產α-淀粉酶、中性蛋白酶、果膠酶、纖維素酶分別為21株、19株、18株、26株。Penicilliumcrustosum、Penicilliumexpansum、Aspergillussteynii、Aspergillusclavatus能同時產4種酶;Stagonosporopsiscucurbitacearum、Epicoccumnigrum、Aspergillusochraceus、Aspergillustabacinus能同時產3種酶。其余菌株產1種或2種酶或不產酶。
進一步對產透明圈強的菌株進行產酶能力的測試,各選取5株、7株、4株、8株進行α-淀粉酶、中性蛋白酶、果膠酶和纖維素酶活力檢測。由圖6可知,5株霉菌產α-淀粉酶活力為(7.69±0.89)~(13.34±0.89) U/g,以1-49(A.tabacinus)最高;7株霉菌產蛋白酶活力為(1 900.00±213.72)~(5 391.76±63.15) U/g,以1-37(Penicilliumlimosum)最高;4株霉菌產果膠酶活力為(4.82±0.08)~(10.19±0.30) U/g,以1-123(Sclerotiniasclerotiorum)最高;8株霉菌產纖維素酶活力為(1 928.15±105.01)~(3 194.33±152.91) U/g,以1-123(Sclerotiniasclerotiorum)最高。
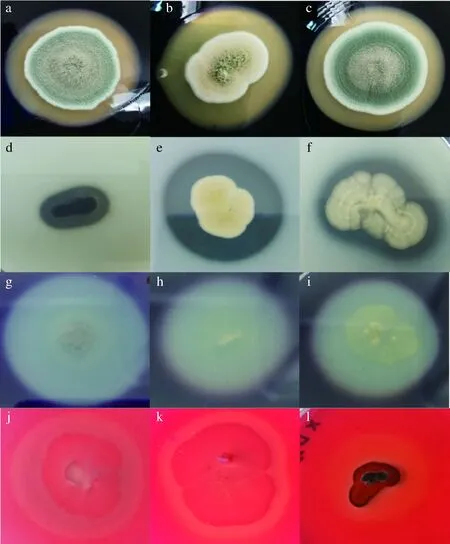
α-淀粉酶活力:a-Aspergillus flavus;b-Aspergillus tabacinus;c-Penicillium coprophilum;中性蛋白酶活力:d-Botryotinia ranunculi;e-Aspergillus tabacinus;f-Aspergillus ochraceus;果膠酶活力:g-Stagonosporopsis cucurbitacearum;h-Aspergillus sclerotiorum;i-Aspergillus clavatus;纖維素酶活力:j-Scopulariopsis brevicaulis;k-Alternaria sorghi;l-Botryotinia ranunculi圖5 部分霉菌株產酶情況Fig.5 Enzymes production of some molds
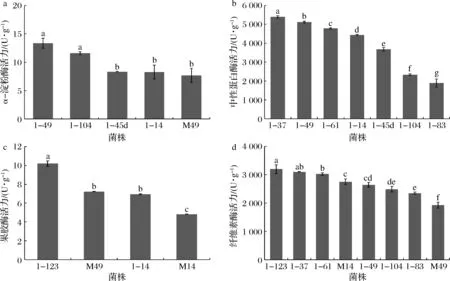
1-49: A. tabacinus;1-104:Talaromyces radicus;1-45d:A. clavatus;1-14:Aspergillus ostianus;M49:Epicoccum nigrum;1-37:P. limosum;1-61:P. limosum;1-83:Scopulariopsis brevicaulis;1-123:S. sclerotiorum;M14:S. cucurbitacearum圖6 部分霉菌菌株的產4種酶能力Fig.6 Four enzymes producing ability of some molds
3 結論與討論
白酒釀造環(huán)境涵蓋生產工藝操作中釀造原料直接或間接接觸的一切環(huán)境,包括空氣、操作工具、地面粉塵等環(huán)境因素。目前濃香型[15]、醬香型[16]、清香型[17]白酒釀造環(huán)境中可培養(yǎng)微生物的研究方法,是對每個釀酒場地的空氣、工具、粉塵等環(huán)境采樣進行平板培養(yǎng)篩選分離,然后對不同場地微生物組成進行比較。本研究對優(yōu)質濃香型白酒釀造環(huán)境空氣中可培養(yǎng)霉菌進行篩選分離,通過ITS rDNA基因序列分析得出曲霉屬(Aspergillus)和青霉屬(Penicillus)為優(yōu)勢菌屬,也是大曲和糟醅中優(yōu)勢菌屬[18-19],在一定程度上反映了不同釀造區(qū)域微生物區(qū)系的變化。
本研究在濃香型白酒核心釀造區(qū)域中檢出交鏈孢屬、帚霉屬,這2個屬主要在醬香型白酒的大曲和酒醅中檢出[3]。此外,與過去研究相比[18, 20-21],本研究在五糧液核心釀造區(qū)域中篩選獲得了Peyronellaea、Quambalaria、Sclerotinia和Stagonosporopsis等4個屬,除Quambalaria來源于窖池區(qū)域,其余屬均來自于釀酒車間外圍環(huán)境中,極大地豐富了核心釀造區(qū)域霉菌的種屬多樣性。Peyronellaeaprosopidi、S.sclerotiorum、S.cucurbitacearum、Quambalariacyanescens主要分布于植物界中,在大曲、酒醅、窖泥等白酒釀造微生態(tài)中未見報道[22],這可能是由于某些霉菌雖然在白酒開放式生產環(huán)境中廣泛存在,但是進入白酒釀造環(huán)節(jié)后,由于基質(大曲、糟醅等)的理化因素,如酸度、酒精度、水分等致使這些霉菌無法生存,進而未能檢出。
過去研究認為,霉菌主要起糖化作用,為制曲和酒醅發(fā)酵提供豐富的酶類,它代謝產生的淀粉酶、蛋白酶、纖維素酶等水解酶系,能將原料中蛋白質、糖類等大分子物質降解為氨基酸、寡糖等小分子營養(yǎng)物質,為其他微生物生長代謝提供能源;另一方面推動著酒體中呈香呈味物質的形成[21]。本研究中P.limosum產中性蛋白酶活力(5 391.76±63.15) U/g,高于劉瑩瑩等[23]優(yōu)化毛霉產中性蛋白酶發(fā)酵培養(yǎng)基,得到中性蛋白酶活力能達到1 310.56 U/g。與本實驗其他菌株相比,S.sclerotiorum產果膠酶活力和纖維素酶活力的能力最高,分別為(10.19±0.30)、(3 194.33±152.91) U/g。KASHYAP等[24]指出Sclerotinia在酸性環(huán)境下能代謝產生多聚半乳糖醛酸酶等類型的果膠酶。A.tabacinus產α-淀粉酶活力(13.34±0.89) U/g最高,但相比鄧加聰等[25]篩選的黑曲霉發(fā)酵產淀粉酶活力(452.23±33.05) U/g性能相對較弱。此外,產α-淀粉酶、蛋白酶、果膠酶、纖維素酶主要集中于青霉屬和曲霉屬中,均占本屬種菌株總數50%以上。在白酒釀造過程中,曲霉屬被廣泛認為主要產淀粉酶、蛋白酶等酶類,青霉屬主要代謝產纖維素酶,這些酶水解產生的糖、多肽或氨基酸可用作酵母菌生長的營養(yǎng)物質和風味化合物形成的前體物質。說明在參與白酒釀造過程中,青霉屬和曲霉屬對原料利用與降解方面發(fā)揮著重要作用,也再次印證青霉屬和曲霉屬在白酒核心釀造區(qū)域中的重要性。
本研究以濃香型白酒核心釀造區(qū)域空氣中可培養(yǎng)霉菌和其所產酶活性種類為對象,比較不同釀造場地空氣中霉菌種群結構差異性,有助于后期系統(tǒng)地開展?jié)庀阈桶拙漆勗煳⑸鷳B(tài)及代謝調控研究。同時,通過研究霉菌所產幾種與實際生產相關聯的功能酶的情況,對釀造微生態(tài)中真菌資源的開發(fā)利用具有積極的指導作用。
