大白菜抗干燒心病研究進展
2023-01-11 02:57:16魏小春原玉香龐文星趙艷艷董曉冰楊雙娟蘇賀楠王志勇趙佳琦牛劉靜張曉偉
中國瓜菜 2022年12期
魏小春,原玉香,李 林,龐文星,趙艷艷,董曉冰,楊雙娟,蘇賀楠,王志勇,趙佳琦,牛劉靜,張曉偉
(1.河南省農業科學院園藝研究所 鄭州 450002; 2.沈陽農業大學園藝學院 沈陽 110000)
1926 年,Mcginty 等[1]首次發現生菜干燒心病,隨后干燒心病害在各類蔬菜中越來越常見。其中大白菜干燒心病在世界各地被廣泛報道,其發病程度、面積都在逐步增加,干燒心病害也逐漸成為大白菜的主要病害之一。干燒心病也稱“夾皮爛”(圖1),是一種由缺鈣引起的生理性病害。一般從蓮座期開始發病,發病時葉邊緣干枯,向內傾卷,生長受到抑制;隨著病情的加重,嫩葉邊緣呈水漬狀、半透明,脫水后萎蔫呈白色帶狀;發病后期病株外觀未見異常,但其內葉葉緣變干黃化,葉肉呈干紙狀,有不規則病斑,病健界限清晰。貯藏期發病易引起細菌感染,由干心變腐爛,嚴重影響其商品性。筆者對干燒心病的生理生化、鑒定方法及遺傳育種等方面的研究進行歸納總結。
1 大白菜干燒心病害國內外研究現狀
干燒心病害常出現在結球白菜、甘藍、菜花、萵苣等植物上,在歐美被稱為內部頂燒病(internal tipburn or tipburn)、內腐病(internal rot or internal break down)、內部褐變病(internal browing),日本稱之為心腐病(heart rot)、緣腐病(marginal rot)[2]。1946 年,Shafer 等[3]首次報道甘藍的干燒心病及其發病原因。從此國內外學者相繼開展了大量的研究工作。其中美國Mynard 等[4]認為白菜和甘藍的“干燒心”癥狀、發病原因、發病機制基本相同,都是由缺鈣引起的生理性病害。Bangerth[5]也指出植物生理失調發病機制與其組織中鈣離子的含量有關,提高鈣離子的含量可減少病害的發生。
從20 世紀70 年代以來,干燒心病開始在國內的一些省(市)發生。天津地區在1970—1979 近10年內有6 年干燒心病的發病率在60%以上[6]。據1986 年對全國14 個有關省(市)的調查結果,北自黑龍江、南到福建、東起遼寧大連、西至新疆都有不同程度的大白菜干燒心病[7]。對大白菜的產量和品質造成了巨大的影響,已發展成為嚴重威脅大白菜生產的生理性病害。
鈣是大白菜生長發育所必需的營養元素,以Ca2+的形式存在于細胞中,Ca2+不僅對維持細胞壁和細胞膜結構至關重要,還可以保護細胞膜的完整性。Ca2+作為第二信使成分在調控大白菜生長發育和應對環境脅迫方面發揮著重要作用。植物根系從土壤中吸收Ca2+,經根尖及成熟區根毛的表皮,穿過根的外皮層細胞,進入內皮層,最后進入中柱,經質外體或共質體途徑運輸至地上部分。繆穎等[8-10]認為,隨著缺鈣時間的延長,細胞壁膜上的鈣逐漸釋放到細胞間隙中,葉綠體片層膜上的鈣逐漸釋放到細胞液中。根據Ca2+-ATPase 活性的變化及其與細胞壁結構組分變化的關系,推測干燒心病的發生與細胞膜鈣泵活性變化誘導的鈣第二信使功能有關。
李林章等[11]對寧波地區甘藍干燒心病的研究結果顯示,蓮座期和結球期為該病的主要發生時期,心葉為主要的發病部位,發病早期心葉緣呈水浸狀、半透明,發病后期失水褐變干枯,嚴重影響甘藍的貯藏品質,易腐爛。
張燁等[12]通過對山東省惠民縣大白菜干燒心病嚴重發生地塊土壤成分分析,發現其主要原因是缺錳。山東省農業科學院高建偉[13]團隊研究發現,大白菜干燒心病也有細菌性干燒心類型,首次報道了菠蘿泛菌(Pantoea ananatis)可引起大白菜細菌性干燒心病,并對分離得到的P.ananatisY2 菌株進行生物學特征、全基因組測序及其引起大白菜干燒心病發病規律調查,共鑒定出29 個強致病相關基因。然而,目前尚未見所報道的29 個基因在大白菜葉片致病機制方面的研究,對其致病力的研究將有利于今后對葉片細菌性干燒心致病機制及大白菜對其抗性作用機制的解析[14]。
2 大白菜抗干燒心病生理生化研究
Kuo 等[15]研究表明,促進根部鈣離子的運轉及吸收是避免干燒心的有效措施。Aloni 等[16]研究表明,大白菜干燒心病的發生與幼嫩葉片中的鈣素含量呈顯著正相關。Saure[17]認為大白菜干燒心病與其說是缺鈣造成的生理性病害,不如說是一種因逆境脅迫造成的生理失調。大多學者都認為大白菜干燒心病是一種生理性缺鈣病害[18-21],所謂生理缺鈣并非只有植株生長環境缺鈣才會導致,直接的病因是植株對鈣素的吸收利用受阻導致植株內可利用的鈣含量滿足不了植株生長所需。
20 世紀80 年代,南開大學趙素娥等[22]從田間著手調查,開始了對大白菜干燒心病的研究工作,先后分析了氣候、土壤中鹽分與發病的關系,以及病株和健株不同部位鈣化物含量的比較,指出病株含鈣量明顯低于健株,尤其是內葉。安志信等[18]通過心葉補鈣的方法證明此病是由缺鈣引起的生理性病害。王莉莉[21]研究結果表明,大白菜干燒心病是由土壤中缺少水溶性鈣和營養失調引起的,過量施氮也會嚴重影響植株對鈣的吸收,但也有個別學者認為干燒心病是由缺錳引起的[23-24]。根據農業農村部環境保護科研監測所測定,盡管典型鈣質土中有效錳含量較低,石灰性土壤中的交換態錳含量和還原態錳含量可以保證作物所需,從化驗分析結果來看,病健株大白菜中的含鈣量并無明顯規律性,但其含錳量差異顯著,患病白菜含錳量較低。
張純胄等[25]通過葉面噴施的方法,證明在大白菜上噴施錳肥和硼肥比噴施鈣肥防治干燒心病的效果更好,他認為干燒心病在一定程度上與缺錳有關,但不是主要病因,其主要原因是缺水生長和營養吸收不暢引起的代謝失調。
繆穎等[9]研究了大白菜干燒心病發生過程中酶活性的變化情況,發現在缺鈣條件下細胞膜Ca2+-ATPase 的活性發生了變化。擬南芥Ca2+-ATPase 基因ECA的敲除可導致與大白菜干燒心病發病相似的表型。這些結果表明ECA可能參與了大白菜干燒心病的發病進程,進一步的Real-time PCR(熒光定量PCR)檢測結果證實,大白菜ECA和CAS基因的表達受鈣脅迫的影響,說明這2 個基因與大白菜干燒心病的發生密切相關。
對于植物來說,通過外質體和陽離子通道從土壤中吸收Ca2+,在木質部隨蒸騰作用向上運輸[26]。Ca2+在植物細胞中主要存儲在質膜、液泡和內質網(ER),但胞質Ca2+通過濃度的變化在宿主響應各種壓力信號。細胞質Ca2+水平的調節提供了對環境刺激的快速反應,是通過Ca2+的運輸和存儲途徑系統實現的,外源鈣離子的缺失是干燒心病發生的誘因。
3 大白菜干燒心病抗性鑒定方法
由于大白菜自然生長周期較長,使得鑒定時期所需周期更長,干燒心病一般在內葉發生比較嚴重,從外觀上并不能直觀地觀察到發病癥狀。抗性鑒定有利于快速、直觀地觀察干燒心病與抗干燒心病情況。大部分鑒定模式在幼苗期間進行,對于干燒心發生的情況可控,相對占用空間小,方便試驗管理,減少經濟損失,有利于進一步的分子研究。目前,白菜干燒心病鑒定方法主要有傳統田間鑒定法、離體葉片扦插法、室內苗期鑒定法3 種。
3.1 傳統田間鑒定法
將試驗材料育苗后定植在露天大田環境中,統一實行常規管理,結球期縱切剖開調查球心發病情況。優點是田間鑒定較為準確,不需要溫室設施,降低成本。缺點是該鑒定方法費時費工,鑒定周期長,不宜用于大量材料的篩選,發病受環境條件影響大,不同年份之間發病情況差異較大,很難用于抗病育種實踐中。
3.2 室內苗期鑒定法
張鳳蘭等[27]在大白菜育種工作中發現,不同品種和材料對大白菜干燒心病的抗性差異較大,初步建立了大白菜抗干燒心病的室內人工鑒定技術,為培育大白菜抗干燒心病品種提供了有效手段。優點是利用營養液進行育苗可以更準確地控制營養元素的含量,而且相比傳統模式,苗期鑒定法節省空間,節省成本,周期較傳統田間鑒定省時,更適合試驗研究。缺點是白菜幼苗期間需要適宜的溫度、光照、濕度環境條件,所以需要在培養室進行試驗。
3.3 離體葉片扦插法
日本學者吉川宏昭等[28]創建了離體葉片扦插法。優點是能夠簡單、快速、省時地鑒定干燒心病發病情況。缺點是發病差異不明顯,適于大量材料粗劣的鑒定。
目前采用較多的方法是室內苗期干燒心病鑒定方法,又稱為干燒心病(缺鈣)抗性室內鑒定方法。張鳳蘭等[27]根據過施氮會嚴重影響植株鈣吸收的原理,使用了配方中去除了Ca2+、且含N 成分增加1 倍的Hoagland 營養液進行室內培養,初步確立了大白菜對干燒心病抗性的室內苗期人工鑒定技術。隨后不斷完善,大白菜干燒心病苗期鑒定法能排除許多外界干擾,重復性好,能夠觀察到植株的整個發病過程,是一種相對準確的鑒定方法,但在培養時間的長短與始培養苗形態方面并未達成一致。開始處理的苗形態差異很大,有的采用種子播種直接缺鈣培養[29],也有采用2 片[30]、8 片[7]、12 片真葉[6]幼苗培養,處理時間也各有不同。石姜超等[31]在玻璃溫室內采用營養缽栽培,以珍珠巖為栽培基質在使用前先用蒸餾水沖洗2 次,以避免鈣離子的干擾。采用缺鈣的Hongland 營養液配方,即除去原配方中的Ca(NO3)2,使其他含N 成分比原營養液增加約1 倍,總N 量和原營養液基本平衡。采用室內苗期干燒心病鑒定方法的研究報道有很多,但仍存在幼苗黃化、干燒心癥狀不典型以及操作繁瑣等問題。
苗期鑒定法不受環境條件限制,準確率較高,適合少量材料精選。但在育苗和鑒定過程中會受到營養缽珍珠巖中鈣元素的干擾,因此苗期鑒定法還有改良的空間。隨著無土栽培技術的發展,改良苗期鑒定法-營養液水培鑒定法也成為一種創新的鑒定方式。雷娟利等[32]調查大白菜小黑點病時將帶有苗子的水培海綿定植到穴盤中,再統一放入盛有營養液的托盤里進行鑒定。王鑫[33]在調查白菜干燒心病時采用阿夫道寧營養液配方,將材料定植在營養液槽中進行苗期水培鑒定。營養液水培鑒定過程中避免了鈣元素的干擾,環境條件控制更加精確,無土栽培的興起為干燒心病鑒定模式提供了新的思路。
4 大白菜干燒心病抗性遺傳規律研究
大白菜在抗干燒心病和鈣吸收特性上存在基因型差異,但其發病受多種環境因素的誘導。對大白菜干燒心病害抗性遺傳規律的研究結果表明,此病害并不是簡單的顯性與隱性之間的關系,而是屬于非單一基因控制的數量性狀遺傳(表1)。
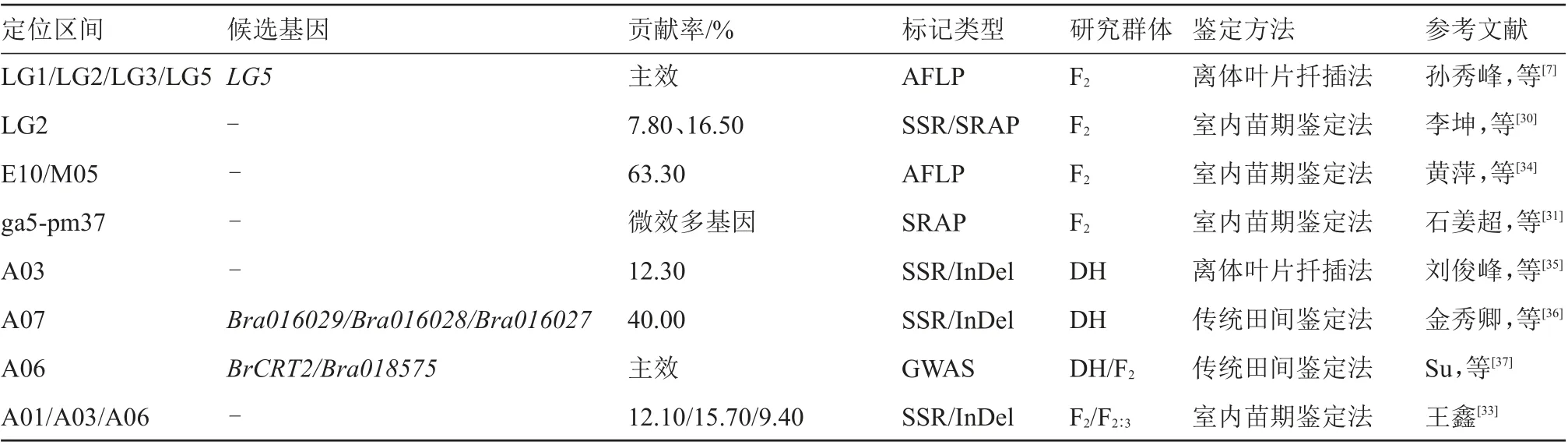
表1 大白菜干燒心病抗性遺傳規律研究進展
孫秀峰等[7]首次將大白菜干燒心病的研究深入到分子方面,構建了AFLP 分子遺傳圖譜,其中包含105 個標記位點、11 個連鎖群、覆蓋長度669.7 cM。在此基礎上檢測到分布在4 個連鎖群上的與抗干燒心病有關的4 個QTL 位點,解釋的遺傳變異范圍在11.0%~58.9%之間,符合正態分布,表明大白菜干燒心病抗病性受微效多基因控制,遺傳表現符合數量遺傳性狀特點。但偏度系數為負數表示該曲線呈負偏態,說明干燒心病可能由不等效多基因控制。將大白菜干燒心病的研究深入到分子水平,這為從分子角度揭示大白菜干燒心病發病機制和闡明合適的鑒定方法提供了理論依據,并為利用分子標記輔助選育大白菜抗干燒心病品種做出了開創性的工作。
李坤等[30]利用SSR 和SRAP 分子標記技術,篩選出2 個與干燒心性狀相關的QTL 位點和3 個與其緊密連鎖的分子標記,通過對QTL 位點加性效應分析可知提高干燒心病抗病性為增效位點,貢獻率分別為7.80%和16.50%,對F2代干燒心性狀進行正態分布檢驗,結果表明干燒心病抗病性遺傳不是簡單的顯隱性的關系,而是符合數量性狀遺傳的特征。黃萍等[34]利用AFLP 和BSA 相結合的方法,檢測到1 個與大白菜干燒心病基因緊密連鎖的AFLP標記E10/M05,該標記與干燒心病病情級數的相關系數為0.633,為極顯著正相關。石姜超等[31]采用營養液培養法來觀察干燒心病發病過程,發現不同時期、不同植株的干燒心病表現不同,初步明確了不同病狀發病特點,F2代植株的抗病性呈現連續分布,干燒心病病害級別的次數分布呈現單峰曲線,符合正態分布,說明干燒心病抗病性受微效多基因控制,符合數量遺傳特征。劉俊峰等[35]以DH 群體為研究材料,構建了一張包含12 個連鎖群、108 個標記位點、覆蓋基因組長度1 004.7 cM、平均圖距9.30 cM 的InDel/SSR 標記連鎖圖,定位了1 個與干燒心性狀相關的QTL 位點,位于3 號染色體上,其貢獻率為12.30%,為增效位點,同時也對大白菜其他農藝性狀的基因定位進行了研究,例如株高、株幅、外葉長、外葉寬、葉形指數等,為大白菜新品種的分子輔助育種提供了理論基礎。金秀卿等[36]報道了2 個與大白菜干燒心抗病性基因緊密連鎖的In-Del 標記,都位于7 號染色體,貢獻率都達到40.00%以上。王鑫[33]利用SSR 和InDel 分子標記構建了一個包含10 個連鎖群、64 個標記位點、總長度893.9 cM 的InDel/SSR 遺傳連鎖圖,共檢測出3個與大白菜干燒心抗病性性狀相關的QTL 位點,位于A01、A03、和A06 連鎖群上,貢獻率分別為12.10%、15.70%和9.40%。通過對每個群體的發病率和發病系數分析,峰值走向呈正態分布,符合數量性狀遺傳特征。以上研究結果均表明,大白菜干燒心病抗性符合數量遺傳規律,部分貢獻率高的位點可以開發為分子標記,為分子標記輔助選育和抗病機制研究提供了理論依據。
Su 等[37]解析了鈣網蛋白BrCRT2(calreticulin 2)在大白菜抗干燒心中的作用,闡述了BrCRT2參與植物胞質Ca2+濃度調控的新機制,該基因編碼的蛋白在其關鍵結構域中有17 個氨基酸的缺失,通過轉基因分析、Ca2+熒光標記和內質網分離體的鈣離子結合/外排等試驗證明,這17 個氨基酸的缺失會導致該蛋白鈣離子結合能力下降,從而降低植物應對缺鈣脅迫的能力,最終導致干燒心病的發生。通過對抗、感2 種材料中的鈣濃度進行分析,結合先前報道的與鈣吸收和運輸相關的差異表達基因進行功能注釋和KEGG 途徑富集分析,發現鈣缺乏不是感病大白菜J95822 發生干燒心病的主要原因,而可能是由后期細菌感染引起的[38]。目前,用于生產的大白菜主栽品種大多不具備抗病性,此外,環境、栽培管理不當等不利因素均可導致干燒心病發生,嚴重影響大白菜的產量和品質。因此,挖掘植物自身的抗干燒心基因、培育抗病新品種是解決白菜干燒心病的最佳途徑。
5 展 望
蔬菜干燒心病是由缺鈣引起的生理性病害,自1946 年Shafer 等[3]首次在甘藍上報道以來,國內外專家對此病的誘因和致病機制進行了大量研究,并取得了一些進展。大白菜原產于我國,是廣大人民喜愛的一種傳統蔬菜,其種植歷史悠久、種類繁多。我國各種類型的白菜為抗病蟲害育種研究提供了極為豐富的種質資源。干燒心病是大白菜生產上發生最普遍的一種生理病害,主要發生在白菜幼嫩葉片上,使大白菜失去食用價值和商品價值,造成嚴重產量損失。我國學者結合生理生化特征與遺傳規律方面進行深入系統的研究之后發現,大白菜在抗干燒心病和鈣吸收特性上存在基因型的差異,但是其發病受到多種環境因素的誘導。
國內外在大白菜干燒心病的發生和防治方面已經取得了很大的成績,比如在生產栽培方面合理安排播期避開高溫天氣,深耕改善土壤環境。生長期加強水肥管理,及時補施鈣素和葉面追肥進行防控等。但在不同基因型中篩選抗干燒心病材料方面仍缺乏簡便有效的方法。目前采用較多的方法是室內苗期干燒心病鑒定方法[27],種子播種直接缺鈣培養[29],離體葉片數為2 片真葉[30]、8 片真葉[7]、12片真葉時[6]幼苗培養。關于苗期干燒心病鑒定方法研究報道有很多,但仍存在幼苗黃化、干燒心病癥狀不典型以及操作繁瑣等問題。雷娟利等[32]將帶有苗子的水培海綿定植到穴盤中,再統一放入盛有營養液的托盤里進行鑒定。王鑫[33]采用阿夫道寧營養液配方,將材料定植在營養液槽中進行苗期水培鑒定。營養液水培鑒定過程中避免了鈣元素的干擾,環境控制更加精確,為干燒心病鑒定模式提供了新的思路。
分子標記技術已被用于大白菜干燒心病的定位和分析,并輔助蔬菜育種。盡管如此,目前對大白菜干燒心的遺傳規律研究還不夠系統和深入。孫秀峰等[7]、李坤等[30]、黃萍等[34]、石姜超等[31]、劉俊峰等[35]、金秀卿等[36]、王鑫[33]等的研究結果均表明,大白菜干燒心病抗性符合數量遺傳規律,部分貢獻率高的位點可以開發為分子標記,為分子標記輔助選育和抗病機制研究提供理論依據。Su 等[37]解析了鈣網蛋白BrCRT2(calreticulin 2)在大白菜抗干燒心中的作用,闡述了BrCRT2參與植物胞質Ca2+濃度調控的新機制,該基因編碼的蛋白在其關鍵結構域中有17 個氨基酸的缺失。
目前,用于生產的大白菜主栽品種大多不抗干燒心病。此外,環境不佳、栽培管理不當等不利因素均可導致干燒心病發生,嚴重影響大白菜的產量和品質,因此,挖掘植物自身的抗干燒心基因、培育抗病新品種是防治白菜干燒心病的最佳途徑。利用現代生物技術的手段、結合改良的干燒心病鑒定方法,是植物營養學、分子生物學以及遺傳育種學之間的有機結合及交叉滲透,也是針對植物營養性狀基因在育種實踐中應用的探討,學科交叉融合也將使大白菜抗干燒心病育種取得更大的成就。
