鹽酸藥根堿對(duì)HPV16 E6/E7-HaCaT細(xì)胞中NF-κB及C2TA蛋白的影響
2023-01-18 13:37:28戴任金銘時(shí)晨李秀商婷婷任青玲
中醫(yī)藥信息 2023年1期
戴任金銘,時(shí)晨,李秀,商婷婷,任青玲?
(1.南京中醫(yī)藥大學(xué)附屬江蘇省中醫(yī)院,江蘇 南京 210000;2.南京中醫(yī)藥大學(xué)中醫(yī)兒科研究所,江蘇 南京 210000)
研究發(fā)現(xiàn),宮頸癌的明確病因是高危型人乳頭瘤病毒(human papilloma virus,HPV)的持續(xù)感染[1],由HPV 感染導(dǎo)致的宮頸癌約占70%~80%[2]。盡管液基細(xì)胞學(xué)檢測(cè)等宮頸篩查手段以及HPV 疫苗的問世降低了疾病發(fā)生率及惡性進(jìn)展率,但此類疾病發(fā)病率在我國仍然處于上升階段[3]。由于病毒E6/E7 蛋白介導(dǎo)的免疫逃逸現(xiàn)象,導(dǎo)致HPV 持續(xù)感染(HR-HPV)并進(jìn)展成為宮頸鱗狀細(xì)胞癌的風(fēng)險(xiǎn)大幅度增加[4],目前臨床尚未有特異性治療宮頸HPV 感染的藥物。
HPV 是小型、無包膜DNA 病毒,通過多種途徑進(jìn)行免疫逃逸,包括E6、E7 蛋白抑制細(xì)胞因子及趨化因子分泌,妨礙抗原提呈功能等。上皮細(xì)胞是病原體感染后的第一道防線,HPV 具有強(qiáng)烈的嗜上皮屬性,角質(zhì)形成細(xì)胞是其天然的宿主細(xì)胞[5],作為表皮的主要成分,角質(zhì)形成細(xì)胞上表達(dá)多種模式識(shí)別受體(pattern recognition receptor,PRRS),此類受體可以識(shí)別包括病毒、脂多糖、肽聚糖、鞭毛蛋白在內(nèi)的病原相關(guān)模式,導(dǎo)致產(chǎn)生促炎細(xì)胞因子和趨化因子,如IL-1β、IL-8 和CCL20 分泌,隨后將白細(xì)胞募集到損傷部位,發(fā)揮免疫殺傷作用[6?8]。角質(zhì)形成細(xì)胞上還存在主要組織相容性復(fù)合體(Major histocompatibility complex,MHC)Ⅰ、Ⅱ類分子,識(shí)別處理抗原并分別將病毒抗原特異性呈遞給CD4+、CD8+T 細(xì)胞,激活效應(yīng)T細(xì)胞,產(chǎn)生大量促炎細(xì)胞因子[9?11]。然而HPV E6、E7 蛋白作為非分泌性蛋白在上皮細(xì)胞中低表達(dá),借此逃避抗原提呈細(xì)胞的識(shí)別,使其能順利整合到宿主基因組以促進(jìn)疾病惡性進(jìn)展。目前關(guān)于宮頸HPV感染導(dǎo)致宮頸鱗狀細(xì)胞癌的機(jī)制研究主要集中于病毒E6、E7 蛋白,大量研究數(shù)據(jù)表明E6、E7 蛋白顯著抑制核轉(zhuǎn)錄因子NF-κB激活,IL-6、IL-1β等細(xì)胞因子的釋放,這些均使白細(xì)胞無法被正常募集到感染部位進(jìn)行免疫應(yīng)答,最終幫助HPV 實(shí)現(xiàn)免疫逃逸[4,12],為腫瘤的發(fā)展提供有益微環(huán)境。C2TA 作為NLRs 家族NLRA 的唯一成員,主要負(fù)責(zé)調(diào)控MHC Ⅱ類分子的表達(dá),與CD4+CTL免疫殺傷機(jī)制密切相關(guān)。
課題組前期研究發(fā)現(xiàn),鹽酸藥根堿是加味二妙顆粒君藥黃柏主要成分之一[13],目前對(duì)其研究主要集中在抗菌及抗腫瘤等藥理學(xué)作用上[14],但該藥物在治療人乳頭瘤病毒導(dǎo)致的宮頸鱗狀細(xì)胞癌中具體作用機(jī)制尚不明確。本研究通過生物信息學(xué)手段篩選宮頸鱗狀細(xì)胞癌組織與正常組織中C2TA 表達(dá)差異(C2TA 的低表達(dá)不利于宮頸鱗狀細(xì)胞癌患者生存),隨后通過體外實(shí)驗(yàn)使用慢病毒構(gòu)建HaCaT-HPV16 E6/E7 穩(wěn)定表達(dá)細(xì)胞模型,旨在研究HPV16 E6/E7 蛋白引起的免疫逃逸過程中鹽酸藥根堿的藥理學(xué)作用以及與C2TA 及NF-κB通路相關(guān)的生物學(xué)機(jī)制。
1 儀器與試劑
1.1 儀器
細(xì)胞培養(yǎng)箱(美國,ThermoFisher);高速冷凍離心機(jī)(德國,Eppendorf,5810R);蛋白核酸分析儀(德國,Eppendorf,Bio Photometer);垂直流超凈工作臺(tái)(德國,ESCO);7500 實(shí)時(shí) 定量PCR 儀(美國,ABAppliedBiosystems,QUANTSTUDIO 7 FLEX QUANTSTUDIO);多功能酶標(biāo)儀(瑞士,TECAN,M200 PRO);電泳槽(美國,Bio-Rad,Mini PROTEAN Tera Cell);濕轉(zhuǎn)轉(zhuǎn)膜儀(美國,Bio-Rad);蛋白印跡-多功能成像系統(tǒng)(美國,Bio-Rad,ChemiDoc MP);轉(zhuǎn)移搖床(中國,海門市其林貝爾儀器有限公司)。
1.2 試劑
鹽酸藥根堿(南京良緯生物科技有限公司,批號(hào):B21451);DMEM 完全培養(yǎng)基(江蘇凱基生物有限公司,批號(hào):KGM12500H-500);0.25% 胰蛋白酶-EDTA酶(美國Gibico公司,批號(hào):25200-056);胎牛血清(美國Corning 公司,批號(hào):35-081-CV);CCK-8(上海翌圣生物科技有限公司,批號(hào):40203ES60);NF-κB、IKB-α(武漢三鷹生物技術(shù)有限公司,批號(hào):10745-1-AP、10268-1-AP);C2TA(上海艾比瑪特生物醫(yī)藥公司,批號(hào):TP71691);p-NF-κB、羊抗鼠/兔二抗(美國CST 公司,批號(hào):3033T、7074);β-actin(武漢三鷹生物技術(shù)有限公司,批號(hào):66009-1-Igs);SuperECLPlus 超敏發(fā)光液(上海天能公司,批號(hào):180-501);慢病毒載體pLenti-Ⅲ-HPV16E6-IRESE7Vector(Puro)(鎮(zhèn)江愛必夢(mèng)生物科技有限公司,ABM);Viral Plus Transduction Enhancer(鎮(zhèn)江愛必夢(mèng)生物科技有限公司,批號(hào):No.G698);Polybrene(鎮(zhèn)江愛必夢(mèng)生物科技有限公司,批號(hào):No.G062);HPV16 E6、E7 及GAPDH 引物(鎮(zhèn)江愛必夢(mèng)生物科技有限公司,ABM);RNA 抽提試劑盒(南京諾唯贊生物科技股份有限公司,批號(hào):RC-101);逆轉(zhuǎn)錄試劑盒(南京諾唯贊生物科技股份有限公司,批號(hào):R312-01);Taq Pro Universal SYBR qPCR Master Mix 試劑盒(南京諾唯贊生物科技股份有限公司,批號(hào):Q712-02);蛋白裂解液RIPA(上海碧云天生物技術(shù)有限公司,批號(hào):P0013B);BCA 試劑盒(上海碧云天生物技術(shù)有限公司,批號(hào):P0010S)。
2 方法
2.1 數(shù)據(jù)庫及單基因生物信息學(xué)分析
Timer 數(shù)據(jù)庫是常用的癌癥免疫浸潤標(biāo)志物數(shù)據(jù)庫之一,使用基因模塊分析C2TA 表達(dá)水平與腫瘤組織中浸潤的B 細(xì)胞、CD4+T 細(xì)胞、CD8+T 細(xì)胞、巨噬細(xì)胞、樹突狀細(xì)胞和嗜中性粒細(xì)胞間的關(guān)系;利用GEPIA數(shù)據(jù)庫在相關(guān)性分析模塊中分析C2TA 表達(dá)在宮頸鱗狀細(xì)胞癌組織中的表達(dá)及其與臨床患者預(yù)后間的關(guān)系,以及免疫細(xì)胞標(biāo)志物間是否具有相關(guān)性。
2.2 細(xì)胞培養(yǎng)
HaCaT細(xì)胞在10%胎牛血清+DMEM 完全培養(yǎng)基條件下生長。HaCaT 購自江蘇凱基生物技術(shù)有限公司,HEK293T細(xì)胞由ABM公司提供。
2.3 HaCaT 細(xì)胞系慢病毒感染復(fù)數(shù)(MOI)值確定、Puromycin(Puro)篩選濃度摸索及內(nèi)源性E6/E7基因檢測(cè)
添加Viral Plus Transduction Enhancer 和Polybrene作為感染增強(qiáng)劑,慢病毒感染HaCaT 細(xì)胞72 h 記錄HaCaT 細(xì)胞圖片。設(shè)置MOI 梯度分別為0、5、10、20、50、100,pLenti-CMV-CBH-GFP-2A-Puro-BlankVector 慢病毒感染HEK293T 作為陽性對(duì)照組。HaCaT 細(xì)胞以20%密度培養(yǎng)在含Puromycin 的培養(yǎng)基中,培養(yǎng)72 h 后記錄HaCaT 細(xì)胞形態(tài),Puromycin 濃度范圍為0~5 μg/mL。使用5XAll-In-OneRT Maste rmix with EXCellent CTLysis Kit 提取HaCaT 細(xì) 胞RNA并進(jìn)行cDNA 合成,對(duì)cDNA 樣品分別使用人源HPV16 E6、E7和內(nèi)參GAPDH 的特異性引物進(jìn)行RTPCR。Lenti-Ⅲ-HPV-16 E6/E7(Puro)載體質(zhì)粒轉(zhuǎn)染的293T 細(xì)胞(293T-HPV)和野生型293T 細(xì)胞(293T-WT)為對(duì)照。
2.4 慢病毒包裝
適當(dāng)完全培養(yǎng)基平鋪細(xì)胞于10 cm2培養(yǎng)皿中,將細(xì)胞置于含5% CO2的37 ℃溫箱中孵育24 h后開始轉(zhuǎn)染。將pLenti-Ⅲ-HPV-16E6-IRES-E7Vector(Puro)和 Lenti-ComboPackingMix、lentifectin 共轉(zhuǎn)染 至HEK293T中,完成轉(zhuǎn)染后置于含5% CO2的37 ℃溫箱孵育。8 h 后吸去培養(yǎng)基,加入37 ℃預(yù)熱的完全培養(yǎng)基,繼續(xù)將細(xì)胞放置溫箱孵育,72 h后收集細(xì)胞上清,此時(shí)為未純化的病毒液,濃縮純化處理以獲取高滴度慢病毒保存液,置于?80 ℃冰箱保存。所獲得的慢病毒定名為Lenti-Ⅲ-HPV-16E6-IRES-E7Virus(Puro)。
2.5 穩(wěn)定表達(dá)HPV16 E6/E7 基因HaCaT 細(xì)胞系的建立
在6 孔板中接種密度約為20%~30%的HaCaT 細(xì)胞,24 h 后用慢病毒感染HaCaT 細(xì)胞,在5%的CO2,37 ℃條件下孵育48 h,將感染后的HaCaT 細(xì)胞鋪板96 孔板進(jìn)行穩(wěn)轉(zhuǎn)株抗性篩選,篩選培養(yǎng)基為添加終濃度0.2 μg/mL Puromycin 的完全培養(yǎng)基。篩選3 周后,選擇合適克隆進(jìn)行擴(kuò)增,RT-qPCR法篩選陽性克隆。
2.6 Cell Counting Kit?8(CCK-8)篩選最佳給藥濃度
將100 μL 各組細(xì)胞(5 × 104/mL)接種至96 孔板,每組設(shè)置6 個(gè)復(fù)孔,置于5% CO2、37 ℃、飽和濕度培養(yǎng)箱 中培養(yǎng),過 夜后分別 加入0.001、0.05、0.2、1、5 μmol/L 鹽酸藥根堿,接種后第24、48 h 分別終止培養(yǎng)。向各孔中加入10 μL 的CCK-8 溶液,繼續(xù)培養(yǎng)4 h,用酶標(biāo)儀在450 nm波長下檢測(cè)各組細(xì)胞的OD值。
2.7 實(shí)時(shí)熒光定量聚合酶鏈?zhǔn)椒磻?yīng)分析轉(zhuǎn)錄水平變化
參照RNA抽提試劑盒說明書要求抽提總RNA。應(yīng)用紫外分光光度儀測(cè)定A260/A280 比值及濃度,A260/A280 比值在1.8~2.0 之間為符合純度要求。由上海生工生物科技有限公司依據(jù)熒光定量PCR引物設(shè)計(jì)原則設(shè)計(jì)并合成引物,引物序列見表1。參照逆轉(zhuǎn)錄試劑盒說明書操作,進(jìn)行逆轉(zhuǎn)錄獲得cDNA產(chǎn)物,置于?20 ℃冰箱中保存。按照試劑盒說明書進(jìn)行操作擴(kuò)增。采用2?ΔΔCT法對(duì)RT-qPCR結(jié)果進(jìn)行數(shù)據(jù)處理。

表1 引物序列
2.8 蛋白免疫印跡(Western blot)法檢測(cè)C2TA 及NF-κB信號(hào)通路相關(guān)蛋白
Western blot 法檢測(cè)細(xì)胞中C2TA、NF-κB、p-NF-κB、IKB-α 蛋白表達(dá)。6 孔板中細(xì)胞每孔加入80 μL RIPA 裂解液,裂解液中按50∶1的比例加入蛋白酶抑制劑與磷酸酶抑制劑,冰上裂解,靜置10 min,4 ℃,12 000 r/min 離心10 min,取上清液,BCA 蛋白定量試劑盒檢測(cè)總蛋白濃度,再取適量蛋白加5 × Loading buffer 100 ℃加熱10 min,獲得樣品,?20 ℃保存。通過10% SDS-PAGE80V,30 min,110 V,60 min分離,使用濕轉(zhuǎn)轉(zhuǎn)膜系統(tǒng)400 mA,90 min 轉(zhuǎn)移到PDVF膜,1 × TBST洗滌3次,每次10 min,后封閉液封閉2 h。加入稀釋的一抗(稀釋倍數(shù)均為1∶1 000),搖床4 ℃ 孵育,次日1 × TBST 洗膜3 次,每次10 min。加入二抗(1∶5 000),搖床室溫孵育2 h。再用1 × TBST 洗膜3 次,每次10 min。使用超敏發(fā)光液試劑檢測(cè)。用Image Lab 軟件處理各條帶的灰度值,以目的蛋白與β-actin的灰度比值作為目的蛋白的相對(duì)含量。
2.9 統(tǒng)計(jì)學(xué)方法
數(shù)據(jù)處理采用GraphPad Prism 9.00統(tǒng)計(jì)軟件。數(shù)據(jù)以±s表示。多組間比較,如符合正態(tài)分布且方差齊,用多組間比較采用單因素方差分析,多重比較采用多因素方差分析法,兩兩比較采用單因素方差分析;若數(shù)據(jù)不符合正態(tài)分布或方差不齊時(shí)則用非參數(shù)檢驗(yàn)。P< 0.05表示差異具有統(tǒng)計(jì)學(xué)意義。
3 結(jié)果
3.1 C2TA表達(dá)水平與宮頸鱗狀細(xì)胞癌患者預(yù)后的相關(guān)性
在線數(shù)據(jù)庫GEPIA 對(duì)C2TA 進(jìn)行相關(guān)生存分析,Kaplan-Meier 生存分析發(fā)現(xiàn)C2TA 表達(dá)水平降低的宮頸鱗狀細(xì)胞癌患者OS較差。結(jié)果見圖1。

圖1 GEPIA數(shù)據(jù)庫中C2TA與宮頸鱗狀細(xì)胞癌患者預(yù)后不良的相關(guān)性
3.2 C2TA表達(dá)水平與宮頸鱗狀細(xì)胞癌免疫浸潤細(xì)胞及標(biāo)志物的相關(guān)性
C2TA 表達(dá)水平與宮頸鱗狀細(xì)胞癌純度、免疫B 細(xì)胞、CD4+T 細(xì)胞、CD8+T 細(xì)胞、巨噬細(xì)胞、樹突狀細(xì)胞和嗜中性粒細(xì)胞相關(guān)。結(jié)果見圖2。在此基礎(chǔ)上,探究 C2TA在宮頸鱗狀細(xì)胞癌免疫調(diào)節(jié)中的作用,進(jìn)一步使用 GEPIA 數(shù)據(jù)庫發(fā)現(xiàn)在宮頸鱗狀細(xì)胞癌組織中C2TA的表達(dá)水平與HLA-DMA,HLA-DOB、HLA-DPA1、HLA-DRA、HLA-DPB1、CD74、HLA-DOA、RP11-876、HLA-DQA1等相關(guān)。結(jié)果見表2。

圖2 C2TA表達(dá)水平與宮頸鱗狀細(xì)胞癌純度和免疫細(xì)胞的相關(guān)性
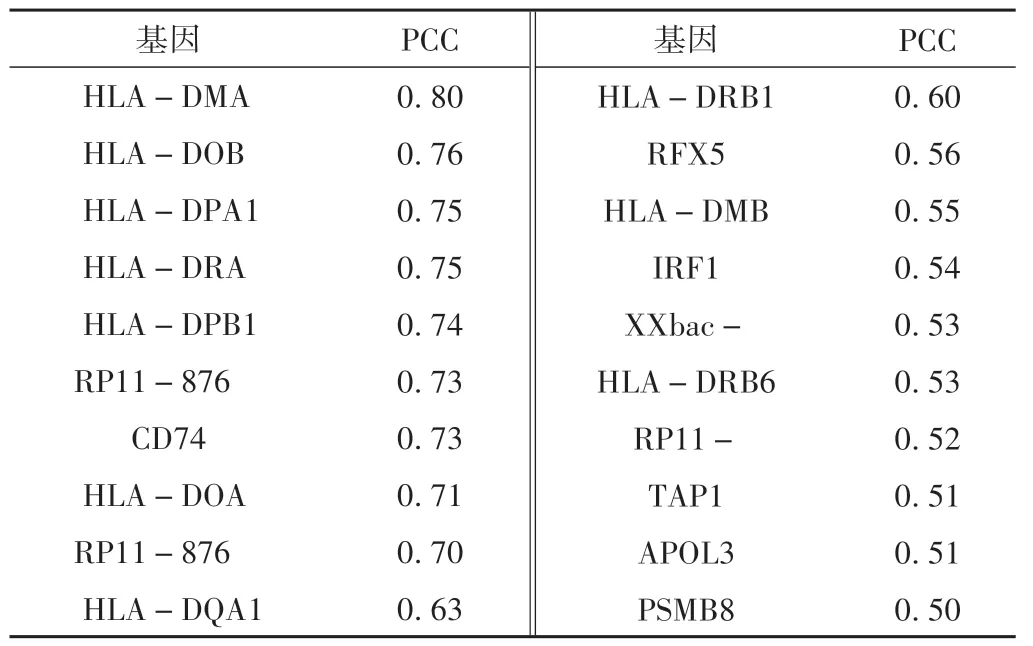
表2 TIMER數(shù)據(jù)庫中宮頸鱗狀細(xì)胞癌組織中的表達(dá)水平與C2TA表達(dá)水平之間的相關(guān)性
3.3 HaCaT-HPV16 E6/E7過表達(dá)模型的建立
挑取HPV16 E6和E7基因及陰性對(duì)照慢病毒轉(zhuǎn)化后的陽性重組克隆,抽提質(zhì)粒DNA 后行酶切鑒定。結(jié)果顯示,pLenti-Ⅲ-HPV16E6-IRES-E7Vector(Puro)載體轉(zhuǎn)化菌落的酶切鑒定產(chǎn)物分別為487 bp和307 bp,片段大小和預(yù)期一致。所獲慢病毒載體重組克隆測(cè)序結(jié)果顯示HPV16 E6/E7基因序列與基因庫人編碼HPV16 E6/E7基因的序列完全吻合。在熒光顯微鏡下可觀察到在陽性對(duì)照慢病毒感染后HaCaT細(xì)胞中大量綠色熒光蛋白表達(dá),提示慢病毒包裝成功,且MOI=20 感染效率80%~90%,細(xì)胞存活率80%(見圖3a),抗性篩選得知Puromycin濃度達(dá)到0.2 μg/mL時(shí)HaCaT 細(xì)胞死亡率為99%,以此濃度進(jìn)行穩(wěn)轉(zhuǎn)株的抗性篩選(見圖3b)。RT-PCR 結(jié)果表明,實(shí)驗(yàn)細(xì)胞HaCaT 中HPV16 E6、HPV16 E7 基因均無表達(dá)。經(jīng)過抗性篩選的HaCaT 穩(wěn)轉(zhuǎn)株RNA 經(jīng)RT-PCR 后條帶大小與預(yù)期符合,HPV16 E6/E7 基因經(jīng)慢病毒介導(dǎo)后成功整合至宿主細(xì)胞基因組且被成功轉(zhuǎn)錄成mRNA(見圖3c),表明穩(wěn)定表達(dá)HPV16 E6/E7 的HaCaT 細(xì)胞株建立。

圖3 慢病毒感染HaCaT細(xì)胞實(shí)驗(yàn)
3.4 鹽酸藥根堿對(duì)HPV16 E6/E7 導(dǎo)致的促炎細(xì)胞因子的影響
研究使用CCK-8 確定給藥濃度,HaCaTHPV16 E6/E7 細(xì)胞經(jīng)鹽酸藥根堿給藥(0.001、0.05、0.2、1、5 μmol/L)后細(xì)胞活力均受影響(P< 0.05),且呈明顯的時(shí)間和劑量依賴關(guān)系,在給藥5 μmol/L 時(shí)細(xì)胞存活率約50%。結(jié)果見表3。因此,本試驗(yàn)選定5 μmol/L 作用24 h 作為鹽酸藥根堿給藥組。C2TA、核轉(zhuǎn)錄因子NF-κB 及IL-6、IL-8、IL-1β、TNF-α 等促炎細(xì)胞因子mRNA 水平明顯受到抑制,隨后給予鹽酸藥根堿后得到恢復(fù),與模型組相比差異具有統(tǒng)計(jì)學(xué)意義。結(jié)果見表4。
表3 各組HaCaT-HPV16 E6/E7細(xì)胞活力的比較(±s,n =3)

表3 各組HaCaT-HPV16 E6/E7細(xì)胞活力的比較(±s,n =3)
注:與24 h存活率比較,1)P < 0.05。
表4 各組HaCaT-HPV16 E6/E7細(xì)胞 NF-κB、IL-6、IL-8、TNF-α mRNA表達(dá)的比較(±s,n =3)

表4 各組HaCaT-HPV16 E6/E7細(xì)胞 NF-κB、IL-6、IL-8、TNF-α mRNA表達(dá)的比較(±s,n =3)
注:與對(duì)照組比較,1)P < 0.05;與模型組比較,2)P < 0.01。
3.5 鹽酸藥根堿對(duì)C2TA蛋白及NF-κB通路的影響
與模型組相比,鹽酸藥根堿處理組中C2TA的蛋白表達(dá)均呈上升趨勢(shì)(P< 0.01)。NF-κB、P-NF-κB、IKB-α是NF-κB通路中的關(guān)鍵蛋白,其激活促進(jìn)細(xì)胞因子IL-6等表達(dá),有利于將白細(xì)胞募集到受感染部位傳遞免疫信號(hào),激活固有免疫應(yīng)答清除病原體。與對(duì)照組相比,NF-κB、P-NF-κB、IKB-α 表達(dá)明顯下降(P< 0.01),而鹽酸藥根堿干預(yù)后呈現(xiàn)回調(diào)趨勢(shì),這說鹽酸藥根堿的作用機(jī)制可能與NF-κB通路以及C2TA蛋白有關(guān),通過激活C2TA蛋白、NF-κB通路及細(xì)胞因子釋放提高上皮細(xì)胞識(shí)別HPV 癌蛋白E6/E7 的敏感性。結(jié)果見圖4和表5。
表5 各組HaCaT-HPV16 E6/E7細(xì)胞中C2TA、p-NF-κB、NF-κB、IKB-α蛋白表達(dá)的比較(±s,n =3)

表5 各組HaCaT-HPV16 E6/E7細(xì)胞中C2TA、p-NF-κB、NF-κB、IKB-α蛋白表達(dá)的比較(±s,n =3)
注:與對(duì)照組比較,1)P < 0.01;與模型組比較,2)P < 0.01。
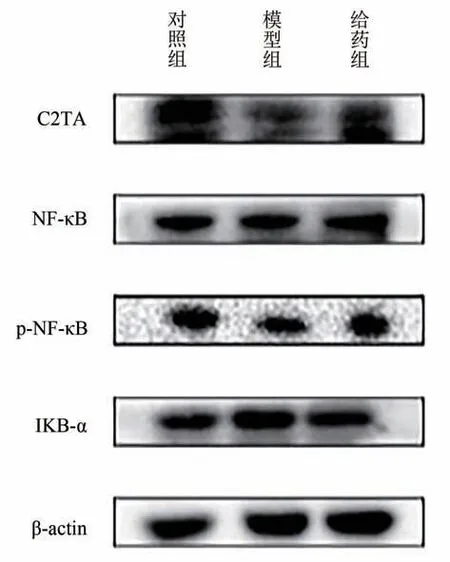
圖4 各組HaCaT-HPV16 E6/E7細(xì)胞C2TA、p-NF-κB、NF-κB、IKB-α蛋白表達(dá)電泳圖
4 討論
人乳頭狀瘤病毒是常見的生殖道病原體,HRHPV 為宮頸鱗狀細(xì)胞癌的發(fā)展提供有利條件[15],E6/E7 是目前已知的兩類存在于宮頸鱗狀細(xì)胞癌中的病毒蛋白[16]。在上皮細(xì)胞的分化過程中E6/E7 通過影響促炎細(xì)胞因子分泌、上皮細(xì)胞黏附能力在內(nèi)的多種途徑幫助病毒基因組順利整合到宿主內(nèi)[17]。研究表明,E6、E7 的表達(dá)影響NF-KB 通路以及下游促炎細(xì)胞因子的激活,而細(xì)胞因子的分泌對(duì)于宿主免疫識(shí)別至關(guān)重要[18]。研究認(rèn)為,由于細(xì)胞因子的分泌對(duì)T 淋巴細(xì)胞受體(T cell receptor,TCR)識(shí)別病毒抗原,激活CTL 中的關(guān)鍵作用,HPV 感染后微環(huán)境中低水平表達(dá)的細(xì)胞因子與宮頸上皮瘤變進(jìn)展相關(guān)[3]。MHC Ⅱ類分子的丟失對(duì)于免疫逃逸并不是必需的。但最近的研究數(shù)據(jù)表明,C2TA 可以調(diào)節(jié)Fas 配體或IL-4 等基因的表達(dá),可能是參與免疫逃逸的關(guān)鍵步驟[19]。因此,推測(cè)C2TA 可通過間接效應(yīng)改變腫瘤生長、腫瘤進(jìn)展或免疫逃逸[20?21]。
HPV E7蛋白通過多種途徑干擾遞呈抗原,其中包括直接干擾高爾基復(fù)合體的內(nèi)環(huán)境,導(dǎo)致病毒抗原肽難以被MHC 類分子識(shí)別或提呈至T 細(xì)胞受體[22]。此外,還包括病毒蛋白的低表達(dá)、影響干擾素作用,抑制細(xì)胞因子[23]、趨化因子[24]、黏附分子[25]表達(dá)等途徑,通過這些機(jī)制逃避免疫識(shí)別、殺傷。近期C2TA 在病毒感染中的作用備受矚目[26?27],C2TA是NLRs家族NLRA的唯一成員,對(duì)MHC Ⅱ類的轉(zhuǎn)錄激活物發(fā)揮主要調(diào)控作用[28?29],低表達(dá)的C2TA 限制MHC Ⅱ分子將病毒蛋白肽正常提呈給CD4+T細(xì)胞,一方面導(dǎo)致下游NF-κB信號(hào)通路活化受限,另一方面導(dǎo)致細(xì)胞因子及趨化因子分泌異常,以上均能幫助病毒逃避CTL 的免疫殺傷[30]。在一項(xiàng)對(duì)埃博拉病毒和SARS 樣冠狀病毒的研究中,研究人員利用轉(zhuǎn)座子介導(dǎo)的基因激活篩選方法發(fā)現(xiàn)C2TA 通過激活第二個(gè)基因CD74 的表達(dá),誘導(dǎo)人細(xì)胞系對(duì)病毒產(chǎn)生抵抗力[9]。有研究表明[9,31?32],C2TA的低表達(dá)導(dǎo)致MHC Ⅱ類分子表達(dá)下調(diào),從而干擾CD4+CTL,使上皮細(xì)胞對(duì)病毒蛋白抗原多肽處于無應(yīng)答狀態(tài),具體表現(xiàn)為NF-κB信號(hào)通路失活以及IL-6、IL-8、TNF-α 等細(xì)胞因子的表達(dá)受限。我們推測(cè)在HPV16 E6、E7 蛋白干擾細(xì)胞因子分泌過程中C2TA 發(fā)揮關(guān)鍵作用,通過恢復(fù)C2TA 的表達(dá)或許能夠逆轉(zhuǎn)這一趨勢(shì),增強(qiáng)免疫應(yīng)答反應(yīng)。藥根堿是從黃連等植物中分離出來的一種四氫異喹啉生物堿,其結(jié)構(gòu)與小檗堿相似,具有解毒殺菌降血糖的作用。在對(duì)黃連中4 種生物堿成分進(jìn)行分析時(shí)發(fā)現(xiàn)藥根堿的抑菌作用最為明顯[33],研究發(fā)現(xiàn)宮頸HPV 的感染與患者陰道菌群失衡密切相關(guān)[34],陰道菌群失衡導(dǎo)致上皮細(xì)胞屏障功能受限,無法正常對(duì)病毒抗原肽進(jìn)行免疫應(yīng)答,也有利于病毒實(shí)現(xiàn)免疫逃逸[35]。
本實(shí)驗(yàn)通過生物信息學(xué)手段發(fā)現(xiàn)C2TA 的低表達(dá)與宮頸鱗狀細(xì)胞癌患者的不良生存情況之間存在聯(lián)系。隨后進(jìn)行體外實(shí)驗(yàn)驗(yàn)證,利用慢病毒載體在角質(zhì)形成細(xì)胞HaCaT上成功構(gòu)建HPV16 E6/E7共同穩(wěn)定表達(dá)的細(xì)胞株,旨在研究HPV16 E6、E7 蛋白協(xié)同致病機(jī)制,鹽酸藥根堿給藥實(shí)驗(yàn)試圖闡明鹽酸藥根堿在HaCaT 細(xì)胞中恢復(fù)免疫激活通路NF-κB 相關(guān)促炎細(xì)胞因子IL-6、IL-8等表達(dá)的作用。通常,宿主一旦檢測(cè)到病原相關(guān)分子模式(pathogen-associated molecular patterns,PAMPs),將會(huì)觸發(fā)細(xì)胞內(nèi)抗病毒信號(hào)級(jí)聯(lián),誘導(dǎo)NF-κB信號(hào)通路激活和各種促炎細(xì)胞因子例如IL-6、IL-8、IL-1β 及TNF- α 等的表達(dá)[18,28,36],然而RT-PCR 結(jié)果顯示,IL-6、IL-8 等細(xì)胞因子在HPV16 E6/E7 過表達(dá)后受到的抑制,這提示E6、E7蛋白干擾上皮細(xì)胞免疫調(diào)節(jié)的能力可能與細(xì)胞因子分泌受限有關(guān)。為了探索E6、E7蛋白引起細(xì)胞因子分泌減少的具體機(jī)制,通過Western blot 法驗(yàn)證的C2TA 蛋白的表達(dá),發(fā)現(xiàn)C2TA 在E6、E7 蛋白過表達(dá)后呈下降趨勢(shì),以上說明細(xì)胞因子分泌減少確與C2TA之間存在聯(lián)系。此外,實(shí)驗(yàn)發(fā)現(xiàn)HPV16 E6/E7 蛋白不僅抑制了細(xì)胞模型中C2TA 蛋白的表達(dá),還干擾了NF-κB 通路的激活,這一現(xiàn)象說明HPV16 E6、E7 蛋白可能通過限制C2TA 的表達(dá)以及干擾NF-κB 通路激活影響細(xì)胞因子的分泌,抑制上皮細(xì)胞免疫反應(yīng)的敏感性。
綜上所述,鹽酸藥根堿可能通過上調(diào)HaCaTHPV16 E6/E7 細(xì)胞中C2TA 表達(dá)及激活NF-κB 調(diào)節(jié)人乳頭瘤病毒16 型E6/E7 蛋白抑制促炎細(xì)胞因子分泌,恢復(fù)上皮細(xì)胞分泌促炎細(xì)胞因子。
