基于高通量測序和組織分離法的虎耳草內生真菌多樣性分析及其抗氧化活性研究
2023-03-21 08:23:34陳美琪黎俊彥譚綺雯李鑒濱姚華雄張玉鑫謝思韻鄧祖軍
中草藥 2023年6期
陳美琪,王 興,黎俊彥,譚綺雯,李鑒濱,姚華雄,張玉鑫,謝思韻,鄧祖軍*
基于高通量測序和組織分離法的虎耳草內生真菌多樣性分析及其抗氧化活性研究
陳美琪1,王 興1,黎俊彥1,譚綺雯2,李鑒濱1,姚華雄1,張玉鑫1,謝思韻1,鄧祖軍1*
1. 廣東藥科大學生命科學與生物制藥學院 廣東省生物活性藥物研究重點實驗室,廣東 廣州 510006 2. 廣州市微生物研究所,廣東 廣州 510663
結合高通量測序技術和傳統可培養技術對虎耳草內生真菌類群及多樣性進行系統分析,并對其抗氧化活性進行初篩,為虎耳草內生真菌資源的開發與利用奠定基礎,也為天然抗氧劑的篩選提供了菌種資源。虎耳草內生真菌多樣性的分析采用Illumina Miseq高通量測序技術和組織分離法,利用化學法檢測虎耳草內生真菌對1,1-二苯基-2-苦基肼自由基(2,2-diphenyl-1-picrylhydrazyl radical,DPPH)、羥基自由基(hydroxyl radical, OH)和2,2′-聯氮-雙-3-乙基苯并噻唑啉-6-磺酸[2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid),ABT] 等自由基的清除率,進而篩選出具有抗氧化活性的菌株。高通量測序分析發現虎耳草內生真菌隸屬于347屬,其中(15.68%)為根部優勢菌屬,莖部的優勢菌屬為(9.57%),而葉部優勢菌屬為炭疽菌屬(5.33%)。利用組織分離法從虎耳草分離到381株內生真菌,隸屬于31個屬,主要類群為炭疽菌屬(19.42%)、籃狀菌屬s(10.76%)和曲霉菌(8.66%)。基于高通量測序的和組織分離法分析的多樣性指數顯示虎耳草根部內生真菌的多樣性明顯高于莖部和葉部,而且虎耳草內生真菌總體多樣性指數均高于多數已報道的其他藥用植物。24株代表性菌株發酵液的抗氧化活性初篩結果顯示:對DPPH、OH和ABTS自由基清除率高于50%的菌株分別占總數的91.67%、79.17%和83.33%,其中菌株sp. HECS10對DPPH、OH和ABTS 3種自由基的清除率均高于75%。虎耳草內生真菌的類群分布具有較高的多樣性,且蘊含著較高比例的抗氧化活性菌種資源,為篩選微生物來源的天然抗氧化劑提供了重要資源庫。
虎耳草;高通量測序;內生真菌;多樣性;抗氧化活性
虎耳草Curt屬虎耳草科虎耳草屬多年生草本藥用植物,其作為一種常用中藥可全草入藥,性微苦、辛、寒,具有祛風清熱,涼血解毒等功效,可用于治療風疹、濕疹、中耳炎、丹毒、肺癰、痔疾等癥[1-2]。已有研究表明虎耳草含有異香豆精類、酚酸類、萜類和黃酮類等多種化合物,具有抑菌、抗炎、抗氧化、保肝、抗腫瘤等藥理活性。例如盧玉棟等[3]發現虎耳草提取物在體外具有良好的抑菌和抗氧化作用。Ji等[4]從虎耳草藥草中分離得到巖白菜素,具有抗腫瘤、抗炎、抗氧化應激和神經保護的特性,可通過激活人磷脂酰肌醇三羥基激酶/蘇氨酸蛋白激酶信號通路改善1-甲基-4-苯基-1,2,3,6-四氫吡啶(1-methyl-4-phenyl- 1,2,3,6-tetrahydropyridine,MPTP)誘發的帕金森氏病。然而,一方面由于人們濫挖亂采,虎耳草野生資源越來越少,難以滿足制藥對藥源的要求[5],另一方面虎耳草在各地區栽培所產的活性提取成分含量及作用效果等存在較大差異從而對后續利用造成了不便[6-7],因此尋找虎耳草藥用資源更穩定有效、可持續的替代利用途徑顯得十分必要。
植物內生菌是指定殖在植物組織內部的微生物群落,它們存在于植物發育的不同階段,且不會對宿主植物產生損害或者感染,包括植物內生真菌、細菌和放線菌[8]。研究表明,植物內生真菌能產生多種結構類型的代謝產物,包括生物堿、芳香類、多肽類、萜類、酚類和黃酮類等成分,具有抗菌、抗病毒、抗炎、抗氧化、免疫抑制劑和抗癌等多種生物學活性[9-10],其中很多內生真菌的代謝產物具有植物宿主提取物相同或相似的結構或活性,如賀鵬飛[11]從中藥烏頭中分離得到的內生菌中發現其次生代謝產物中含有烏頭堿,為烏頭堿的獲得提供了微生物來源;蘇秀麗等[12]從中藥植物黃花倒水蓮內生真菌中發現一株鏈格孢屬HNLF-44菌株具有良好的抑菌和抗氧化活性,與前人對宿主植物化學成分和藥理活性所做的研究結果相符,因此內生真菌是探尋新型活性物質的重要新資源,也為藥用植物資源的持續利用和保護提供了替代途徑。全面了解藥用植物內生真菌的類群與多樣性信息是充分挖掘和利用植物內生真菌資源的前提。近年來,高通量測序技術可直接在宿主組織內檢測內生真菌,具有測序基數大、深度高等優點,能更加全面直觀地分析植物組織內生真菌多樣性[13],有研究者利用高通量測序技術分析了朝鮮淫羊藿葉[14]、百合[15]的內生真菌群落多樣性,獲得了大量的內生真菌類群信息,為后續藥用植物內生真菌的分離和利用奠定了基礎。目前大量研究證實虎耳草提取物具有良好的抗氧化活性[7,16],但關于虎耳草的內生真菌生物多樣性及其抗氧化活性的研究未見報道。因此本研究擬結合高通量測序和傳統可培養技術對虎耳草內生真菌的多樣性進行系統分析,并對其代謝產物的抗氧化活性進行篩選,從而為虎耳草內生真菌資源的后續利用奠定了基礎,也為微生物來源的天然抗氧化劑篩選提供菌種資源,同時為解決虎耳草資源緊缺提供了替代利用途徑。
1 材料與儀器
1.1 材料
供試植株為新鮮、健康(無明顯病癥)的虎耳草,于2021年7月7日采集自湖南省張家界慈利縣(29°23′90.2N,111°07′77.6E),株高約16 cm,經華南植物園陳貽竹研究員鑒定為虎耳草Curt。將植株完整采摘后送至實驗室并于4 ℃冰箱保存備用。
1.2 主要試劑及儀器設備
高效植物基因組DNA提取試劑盒,天根生化科技有限公司;真菌DNA小量提取試劑盒,Magen公司;引物(ITS1和ITS4),生工生物工程(上海)有限公司;馬鈴薯葡萄糖固體培養基(PDA)和液體培養基(PDB),廣東環凱生物技術有限公司;1,1-二苯基-2-苦基肼自由基(DPPH),購自Sigma公司;抗壞血酸(Vc)、水楊酸、硫酸亞鐵、2,2′-聯氮-雙-3-乙基苯并噻唑啉-6-磺酸(ABTS)、過硫酸鉀為分析純,上海麥克林有限公司;SW-CJ-1F型單面雙人潔凈工作臺,蘇州蘇潔凈化設備有限公司;ETC811基因擴增儀,北京東勝創新生物科技有限公司;JY-C電泳儀,北京君意東方電泳設備有限公司;SpectraMax i3x酶標儀,MD美谷分子公司。
2 方法
2.1 虎耳草植株樣品的表面消毒
將新鮮健康虎耳草植株在流水下沖洗以去除土壤顆粒等表面雜質,用無菌剪刀剪切至適量大小,無菌水清洗3~5遍并用無菌濾紙吸干,依次在75%的酒精中浸泡1 min,含3%~5%有效氯的NaClO溶液中浸泡根部1 min、莖部2 min、葉部1.5 min,再用75%酒精浸泡30 s,最后用無菌水漂洗3遍,并收集最后一次漂洗樣品的無菌蒸餾水,經平板涂布檢查無菌后進行后續實驗,樣品則靜置于無菌濾紙上吸去多余水分,晾干。
2.2 虎耳草植株總DNA的提取及PCR擴增
將按“2.1”項消毒好的虎耳草不同部位通過高效提取試劑盒提取樣品基因組總DNA,通過瓊脂糖凝膠電泳法對提取的DNA純度及濃度進行驗證,確認合格后送至北京諾禾致源科技股份有限公司進行檢測。
2.3 高通量測序及數據處理
根據Barcode序列和PCR擴增引物序列從下機數據中拆分出各樣本數據,截去Barcode和引物序列后進行拼接、過濾、質量控制、與物種注釋數據庫進行比對檢測并去除嵌合體序列,最終得到有效數據。以97%的一致性[17]將聚類成為可操作分類單元(operational taxonomic units,OTUs),用Qiime軟件(V 1.9.1)對OTUs中出現頻數最高的序列進行物種注釋分析[18],并分別在門綱目科屬種各個分類水平統計各樣本的群落組成。使用MUSCLE(V 3.8.31)軟件進行快速多序列比對,得到所有OTUs代表序列的系統發生關系,再對各樣本的數據進行均一化處理后進行Alpha多樣性分析和Beta多樣性分析,最后使用R軟件(V 2.15.3)繪制稀釋曲線,Rank abundance曲線等。
2.4 虎耳草內生真菌可分離培養、純化及其多樣性分析
將虎耳草植株按“2.1”項步驟進行表面消毒驗證無菌后,采用文獻[19]中的組織塊分離法進行虎耳草內生真菌的分離,將晾干后的樣品剪切至適當小片,接種至PDA培養板在28 ℃下培養1~2周,每日觀察,若有菌絲從植物組織切口長出,及時挑出轉移至新的平板上,純化培養后接種至斜面培養基,4 ℃保存備用。參考文獻[20-22]中的方法根據菌落的菌絲和孢子等形態學特征及ITS測序進行內生真菌的分類鑒定。然后,按照真菌基因組DNA提取試劑盒的說明書提取真菌DNA,引物為ITS1和ITS4,再將經瓊脂糖凝膠電泳檢測合格后的PCR產物送至生工生物工程(上海)股份有限公司測序鑒定,利用BLAST(http://blast.ncbi.nlm.nih.gov/Blast. cgi)在GenBank中獲取與鑒定菌株序列相似的相似序列信息。
2.5 虎耳草內生真菌抗氧化活性菌株篩選
選取虎耳草不同部位代表性菌株進行DPPH、OH和ABTS自由基抗氧化活性篩選。接種適量菌餅至PDB培養基中,28 ℃、180 r/min培養5~7 d,然后將發酵液依次通過紗布、濾紙進行濾過,再用0.22 μm微孔濾膜對發酵液進行過濾除菌,收集得到內生真菌發酵液,于4 ℃冰箱保存備用。
參考Subbiah等[23]的方法并加以改進后進行測定。采用無水乙醇溶液將DPPH配制成0.05~0.1 mmol/L的溶液。以Vc為陽性對照,在96孔板中分別以A為100 μL不同內生真菌發酵液或者陽性對照品與100 μL的DPPH混合溶液,c為100 μL內生真菌發酵液與100 μL的無水乙醇溶液混合,0取100 μL無水乙醇溶液與100 μL的DPPH溶液,即以無水乙醇作空白對照,震蕩搖勻,在25 ℃避光反應30 min,最后在517 nm下檢測值,每個樣品平行測3次,DPPH自由基清除率計算如公式(1)所示。
DPPH自由基清除率=1-(A-c)/0(1)
OH自由基測定采用水楊酸法,參照文獻中Hu等[24]的方法并加以改進后進行測定。稱量試劑,配制0.1%的H2O2溶液,濃度為6 mmol/L的FeSO4溶液和水楊酸乙醇溶液。以Vc為陽性對照,在96孔板中依次加入6 mmol/L的FeSO4溶液、水楊酸乙醇溶液各50 μL,不同內生真菌發酵液或者陽性對照品50 μL,然后加入0.1%的H2O2溶液50 μL混合均勻,設為B,其中,對照管c以B為基礎,用無水乙醇替代水楊酸乙醇溶液,0以B為基礎,用無菌蒸餾水替代內生真菌發酵液,總體積為200 μL,搖勻后在37 ℃下孵育30 min,510 nm處測定值,每組實驗平行操作測定3次,OH自由基清除率計算見公式(2)。
OH自由基清除率=1-(B-c)/0(2)
參考文獻中Zou等[25]方法并加以改進后進行測定。首先,配制ABTS溶液,將7 mmol/L ABTS和140 mmol/L過硫酸鉀溶液混合制備ABTS儲備溶液,在黑暗環境中孵育16 h,再用無水乙醇溶液將其在734 nm下的值調整至0.70±0.02。以Vc為陽性對照,在96孔板中加入50 μL的不同內生真菌發酵液或者陽性對照品(Y)和200 μL ABTS溶液(Y),混合搖勻10 s以充分混合,避光靜置6 min,用無水乙醇作為空白對照(Y),在734 nm波長下檢測值,每個樣品平行測3次,ABTS自由基清除率計算見公式(3)。
ABTS自由基清除率=1-(Y-c)/0(3)
2.6 數據分析
虎耳草內生真菌的定殖率(colonization rate,CR)、分離率(isolation frequency,IF)、相對分離頻率(relative frequency,RF)、Shannon-Wiener多樣性指數(′)、均勻度指數()以及相似性指數(s)參考孟祥才等[26]的方法進行計算,不同部位間采用Duncan′s多重比較法進行分析,統計顯著水平為<0.05。
3 結果與分析
3.1 高通量測序分析真菌多樣性分析
3.1.1 虎耳草高通量測序樣本數據統計與Alpha多樣性分析 利用Illumina高通量測序技術,從虎耳草根、莖、葉3個部位的樣品中共得到有效序列660 231條,其中根部244 780條,莖部188 596條,葉部226 855條(表1)。基于97%的有效性進行OTU聚類分析從根、莖、葉分別獲得507、377、550個OTUs(圖1)。樣品的稀釋曲線顯示其隨著測序數量的增加趨向平坦(圖2),且3個部位的OTU覆蓋指數均為1(表1),這表明測序結果能夠較好地反映虎耳草各部位間微生物的真實情況。葉部的Chao1和ACE指數最高,其次為根部,莖部最低,這表明虎耳草葉部的內生真菌物種豐富度最高,而莖部最低。根部的Shannon和Simpson指數要明顯高于莖部和葉部,這表明虎耳草根部的內生真菌物種多樣性最高,而莖部和葉部之間差異不大(表1)。虎耳草的根、莖、葉3部位共有的OTU為96個,根部和莖部之間共有194個OTU,莖部與葉部共有130個OTU,葉部與根部之間共有158個OTU,其中根部和莖部之間共有OTU數量最多,這表明這2個部位含有較多相似或是相同的物種(圖1)。

表1 虎耳草根、莖、葉中內生真菌的Alpha多樣性指數

圖1 基于OTU的虎耳草根莖葉各部位Venn圖

圖2 虎耳草根莖葉各樣品的稀釋曲線
3.1.2 虎耳草高通量測序分析內生真菌的群落組成分析 基于高通量測序分析,虎耳草內生真菌隸屬于11門、45綱、100目、205科和347屬。在門的分類水平上(圖3-a),子囊菌門(Ascomycota)是虎耳草根(84.81%)、莖(39.82%)、葉(27.09%)的共有優勢菌門,其次為擔子菌門(Basidiomycota),分別占根、莖、葉的8.97%、7.23%和7.43%。在綱分類水平上,虎耳草根部的主要類群有糞殼菌綱(Sordariomycetes)、錘舌菌綱(Leotiomycetes)、散囊菌綱(Eurotiomycetes)和傘菌綱(Agaricomycetes),分別占比49.14%、19.39%、11.18%和8.74%;莖部中糞殼菌綱(24.41%)、座囊菌綱(Dothideomycetes,7.66%)和散囊菌綱(5.19%)的相對豐富較高,而在葉部的主要類群為糞殼菌綱、座囊菌綱和節擔菌綱(Wallemiomycetes),分別占15.73%、4.74%和4.46%(圖3-b)。在目分類水平上,虎耳草根部優勢類群為肉座菌目(Hypocreales,26.84%)和柔膜菌目(Helotiales,18.69%);莖部為糞殼菌目(Sordariales)、肉座菌目和格孢腔菌目(Pleosporales),分別占比為10.75%、7.16%和6.76%;另外,肉座菌目(6.98%)、Glomerellales(6.69%)和節擔菌目(Wallemiales,4.46%)是葉部的優勢類群(圖3-c)。在科分類水平上,赤殼科(Nectriaceae)為虎耳草根、莖、葉的共同優勢菌科,分別占26.19%、9.69%、6.88%(圖3-d)。在屬分類水平上,、和為根部優勢菌屬,分別占比15.68%、7.23%和6.03%;莖部的優勢菌屬為(9.57%)、鏈格孢屬(4.15%)和(2.13%);炭疽菌屬、麗赤殼菌屬和屬為葉部優勢菌屬,分別占比5.33%、4.63%和4.46%(圖4-b)。
3.2 基于可培養方法的內生真菌多樣性分析
3.2.1 虎耳草內生真菌的分離率和定殖率 表面消毒最后一次漂洗用無菌水經平板涂布培養7 d后,檢查無菌生長,這表明分離得到的菌株均屬虎耳草的內生真菌。從430塊虎耳草不同部位組織塊中共分離得到381株內生真菌,其中根部103株,莖部136株,葉部142株(表2)。虎耳草葉、根、莖部內生真菌的定殖率分別為89.33%、79.23%和78.63%,其內生真菌分離率分別為94.67%、90.67%和79.23%,3個部位中葉部內生真菌的分離率和定殖率均為最高(表2)。
3.2.2 虎耳草內生真菌的類群組成 利用組織分離法共從虎耳草分離到381株內生真菌,經形態學鑒定和ITS-rDNA序列分析,其屬于31個屬,其主要類群為炭疽菌屬(19.42%)、籃狀菌屬(,10.76%)、曲霉菌(,8.66%)、青霉菌屬(7.87%)、麗赤殼屬(7.61%)和間座殼屬(7.35%)(表3)。虎耳草內生真菌不同部位之間內生真菌類群組成存在一定差異:根部內生真菌有20個屬,其優勢菌屬為炭疽菌屬、籃狀菌屬和,相對分離頻率分別為10.68%、10.68%和9.71%;莖部共分離得到12個屬,優勢屬為炭疽菌屬(15.44%)、籃狀菌屬(12.50%)、間座殼屬(11.76%)和曲霉菌(11.03%);另外,葉部分離到12個屬,炭疽菌屬(29.58%)、麗赤殼屬(11.97%)和青霉菌屬(10.56%)為主要類群。

a-門分類水平 b-綱分類水平 c-目分類水平 d-科分類水平
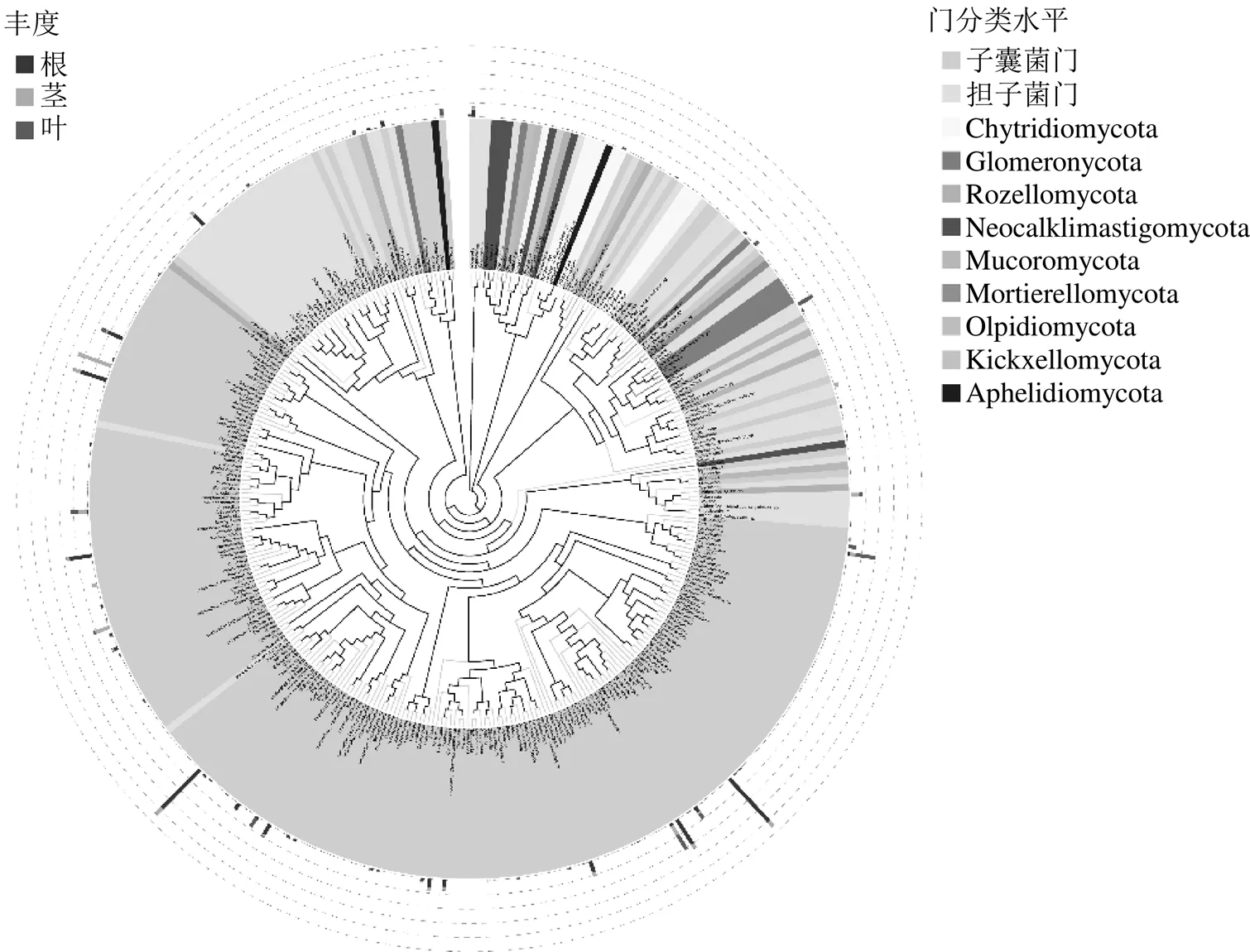
圖4 虎耳草根莖葉中內生真菌在屬分類水平上的相對豐度

表2 虎耳草根、莖、葉內生真菌的分離率和定殖率
同列的不同小寫英文字母表示根、莖葉不同部位之間具有顯著差異(<0.05)
Different lowercase letters in the same column indicate significant differences between different parts of roots, stems and leaves (< 0.05)
3.2.3 虎耳草不同部位內生真菌的多樣性和相似性比較 虎耳草內生真菌的為3.01,其中根部值最高(2.81),其次是莖部(2.36),葉部(2.18)最低。虎耳草莖部(0.95)和根部(0.94)的內生真菌均勻度指數明顯高于葉部(0.88)(<0.05)。莖部與葉部、根部與莖部、根部與葉部之間的內生真菌組成的相似性指數介于0.25~0.50,均為中等不相似(表4)。
3.3 虎耳草內生真菌抗氧化菌株篩選結果
挑選出24株代表性內生真菌菌株,其分子鑒定結果見表5,并對它們的抗氧化活性進行了篩選,結果發現,DPPH自由基清除率大于50%的菌株有22株,占測試菌株的91.67%,高于80%的菌株有4株(占16.67%),分別為sp. HECR12、sp. HECS10、sp. HECL35和sp. HECS27,其中菌株sp. HECS27(91.67%)的清除率最高;OH自由基清除率大于50%的菌株有19株(占79.17%),高于80%的菌株有sp. HECS27和sp. HECL23,其中菌株sp. HECL23的清除率(98.88%)高于陽性對照組(Vc),具有很強的OH清除能力;ABTS自由基清除率大于50%的有20株,占測試菌株的83.33%,其中高于80%的有sp. HECS30和sp. HECS31,其清除率為別為81.27%和88.70%。菌株sp. HECS27對DPPH、OH自由基清除率均高于80%,菌株sp. HECS10對DPPH、OH、ABTS 3種自由基的清除率均高于75%(表6)。

表3 虎耳草內生真菌的種類組成

表4 虎耳草根、莖、葉中內生真菌的多樣性
第4~5列中不同小寫字母表示不同部位的多樣性指數和均勻度指數具有顯著性差異(<0.05)
Different lowercase letters in columns 4—5 indicate significant differences in diversity index and uniformity index of different parts (< 0.05)

表5 虎耳草內生真菌的分子鑒定
4 討論
藥用植物具有豐富的內生菌資源[26],對其群落結構及多樣性進行分析,進而指導篩選功能性菌株可以更好地實現藥用植物資源可持續利用,也為藥用微生物資源的開發提供更可靠的菌種信息。本研究采用Illumina Miseq高通量測序技術并結合傳統培養方法對中藥虎耳草植株進行了內生真菌類群結構及多樣性分析。基于Illumina Miseq高通量測序共從虎耳草檢測出347屬的內生真菌,明顯高于可培養分離得到的31個屬,這說明高通量測序技術獲得內生真菌類群和多樣性信息更為豐富和全面。高通量測序結果顯示虎耳草根、莖、葉3個部位中的優勢菌門為子囊菌門和擔子菌門,這與青海云杉針葉內生真菌類群相似[27]。另外,其優勢菌屬有、、、、鏈格孢屬、、炭疽菌屬和麗赤殼菌屬等,其中鏈格孢屬、炭疽菌屬在其他植物內生真菌中也較為常見[28-19]。組織分離法得到的相對豐度較高優勢菌屬如炭疽菌屬、和麗赤殼菌屬均在高通量測序中檢測到,但存在一部分類群在高通量測序中并不是優勢菌,這可能與其在分離培養基中更易生長有關[31]。虎耳草內生真菌基于高通量分析的α多樣性Shannon指數(根5.16、莖2.96、葉2.88)與基于可培養方法分析的Shannon-Wiener指數(總體多樣性3.01)均高于多數已報道的其他藥用植物[27,31-32],這表明虎耳草蘊含較為豐富的內生真菌資源,這可能與植物的類型及其特定的生存環境、氣候條件等因素有關[33]。高通量測序分析和組織分離法均發現虎耳草根部內生真菌多樣性明顯高于莖部和葉部,這可能與根部長期接觸蘊含豐富微生物資源的土壤有關[34]。結合高通量測序和傳統可培養技術可以獲得較為全面的虎耳草內生真菌群落信息和菌株資源,這為虎耳草內生真菌藥用資源的后續開發與利用奠定了基礎[35]。

表6 虎耳草內生真菌發酵液對3種自由基的抗氧化活性
當前,氧化應激已被證明在許多情況下會加速疾病的發展,包括糖尿病、心血管疾病、阿爾茨海默癥和帕金森癥、急性肺損傷、放射性損傷、癌癥,甚至衰老等[36]。幾乎所有生物體都存在內源性抗氧化防御和修復系統以防止機體氧化損傷,但這些系統通常不足以完全防止損傷[37]。因此,尋找天然抗氧化劑以減少機體的氧化損傷在食品工程和醫藥等領域都具有重要的應用價值。現有研究表明虎耳草具有良好的抗氧化活性,關于虎耳草內生真菌的抗氧化活性未見報道。本研究對24株虎耳草內生真菌代表性菌株發酵液進行DPPH、OH和ABTS自由基抗氧化活性篩選,結果顯示受測試菌株均具有不同程度的抗氧化活性,其中對DPPH、OH和ABTS自由基具有50%以上活性的菌株分別占受測總數的91.67%、79.17%和83.33%。菌株sp. HECS27對DPPH(91.67%)和OH自由基(86.57%)都具有較高的清除活性,菌株sp. HECL23對OH自由基的清除率達到98.88%;菌株sp. HECS30(81.27%)和sp. HECS31(88.70%)對ABTS自由基具有很好的清除效果,而菌株sp. HECS10對DPPH、OH和ABTS 3種自由基的清除率均高于75%。以上結果表明虎耳草內生真菌中蘊含中豐富的具抗氧化活性的菌株,它們為篩選微生物來源的天然抗氧化劑提供重要資源庫,也為虎耳草藥用資源的可持續利用提供了替代途徑,其中活性優良菌株有待于后續進一步深入研究。
利益沖突 所有作者均聲明不存在利益沖突
[1] 何紅梅, 戴岳, 夏玉鳳. 虎耳草的研究概況 [J]. 中國野生植物資源, 2017, 36(2): 75-78.
[2] 張慧, 李秋月, 賀尚文, 等. 虎耳草化學成分及藥理活性研究進展 [J]. 動物醫學進展, 2021, 42(1): 94-99.
[3] 盧玉棟, 范敏, 覃貝妮, 等. 虎耳草有效成分的提取及其生物活性的研究 [J]. 福建師范大學學報: 自然科學版, 2018, 34(4): 68-74.
[4] Ji Y F, Wang D, Zhang B A,. Bergenin ameliorates MPTP-induced parkinson′s disease by activating PI3K/Akt signaling pathway [J]., 2019, 72(3): 823-833.
[5] 蒙先舉, 陶文丞, 韋景楓, 等. 虎耳草組培快繁技術試驗研究 [J]. 中國園藝文摘, 2010, 26(12): 27-28.
[6] 馬曉彤, 周蓉蓉, 張晶云, 等. 虎耳草科植物指紋圖譜及抗氧化活性研究進展 [J]. 廣州化工, 2022, 50(1): 27-28.
[7] 文培華, 顏怡冰, 王文君, 等. 虎耳草總黃酮含量測定及其抗氧化性 [J]. 食品工業, 2022, 43(2): 315-319.
[8] Gouda S, Das G, Sen S K,. Endophytes: A treasure house of bioactive compounds of medicinal importance [J]., 2016, 7: 1538.
[9] Manganyi M C, Ateba C N. Untapped potentials of endophytic fungi: A review of novel bioactive compounds with biological applications [J]., 2020, 8(12): 1934.
[10] Wu W, Chen W H, Liu S Y,. Beneficial relationships between endophytic bacteria and medicinal plants [J]., 2021, 12: 646146.
[11] 賀鵬飛. 烏頭內生菌的分離鑒定及其次生代謝產物研究 [D]. 沈陽: 沈陽農業大學, 2020.
[12] 蘇秀麗, 李惠敏, 唐輝, 等. 黃花倒水蓮內生真菌生物活性評價及HNLF-44菌株鑒定 [J].廣西植物, 16: 1-10.
[13] Tamo?iūn? I, Andriūnait? E, Stanys V,. Exploring diversity of bacterial endophyte communities using advanced sequencing technology [A] //[M]. Singapore: Springer Singapore, 2019: 447-481.
[14] 吳媛, 肖井雷, 姜大成, 等. 基于高通量測序技術的朝鮮淫羊藿內生真菌多樣性分析 [J]. 分子植物育種, 1-10.
[15] 王安萍, 馮關萍, 曾建忠, 等. Illumina高通量測序分析百合內生真菌群落多樣性 [J]. 井岡山大學學報: 自然科學版, 2022, 43(4): 30-36.
[16] 張巖, 陳懷慶, 陶金巧, 等. 虎耳草提取物對酪氨酸酶的激活及抗氧化作用 [J]. 食品工業, 2019, 40(11): 174-177.
[17] Edgar R C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads [J]., 2013, 10(10): 996-998.
[18] K?ljalg U, Nilsson R H, Abarenkov K,. Towards a unified paradigm for sequence-based identification of fungi [J]., 2013, 22(21): 5271-5277.
[19] Deng Z J, Zhang R D, Shi Y,. Characterization of Cd-, Pb-, Zn-resistant endophyticsp. MXSF31 from metal accumulatingand its potential in promoting the growth of rape in metal-contaminated soils [J]., 2014, 21(3): 2346-2357.
[20] Dhayanithy G, Subban K, Chelliah J. Diversity and biological activities of endophytic fungi associated with[J]., 2019, 19(1): 22.
[21] Rocha P D S D, Paula V M B, Olinto S C F,. Diversity, chemical constituents and biological activities of endophytic fungi isolated fromraddi [J]., 2020, 8(6): 859.
[22] 吳可欣, 唐詩雨, 朱奕儒, 等. 野生型斜莖黃芪內生真菌分離鑒定及苦馬豆素分析 [J]. 草地學報, 2022, 30(7): 1692-1700.
[23] Subbiah V, Zhong B M, Nawaz M A,. Screening of phenolic compounds in Australian grown berries by LC-ESI-QTOF-MS/MS and determination of their antioxidant potential [J].(), 2020, 10(1): 26.
[24] Hu T Y, Subbiah V, Wu H J,. Determination and characterization of phenolic compounds from Australia-grown sweet cherries (L.) and their potential antioxidant properties [J]., 2021, 50(6): 34687-34699.
[25] Zou X D, Bk A, Rauf A,. Screening of polyphenols in tobacco () and determination of their antioxidant activity in different tobacco varieties [J]., 2021, 39(6): 25361-25371.
[26] 孟祥才, 于鵬程, 何錄文, 等. 從社會發展角度探討中藥資源的歷史、現在與未來 [J]. 中草藥, 2022, 53(16): 5235-5244.
[27] Strobel G. The emergence of endophytic microbes and their biological promise [J].(), 2018, 4(2): 57.
[28] 方泰軍, 白露超. 青海云杉針葉內生真菌群落結構及多樣性分析 [J]. 分子植物育種, 2022, 20(15): 5198-5204.
[29] 陳少如, 許曉晨, 趙文寶, 等. 閩楠內生真菌高通量分離及種類多樣性分析 [J]. 森林與環境學報, 2022, 42(5): 544-551.
[30] 顧美英, 張志東, 唐光木, 等. 黑果枸杞不同組織內生真菌群落組成及生態功能分析 [J]. 菌物學報, 2022, 41(8): 1254-1267.
[31] 周楠, 姜成英, 劉雙江. 從環境中分離培養微生物: 培養基營養水平至關重要 [J]. 微生物學通報, 2016, 43(5): 1075-1081.
[32] 張秫華, 方千, 賈紅梅, 等. 川芎非根際、根際及根莖內生真菌群落差異分析 [J]. 生物技術通報, 2021, 37(4): 56-69.
[33] 彭黎明, 蔣顯鋒. 紅棕杜鵑根系真菌群落組成與多樣性分析 [J]. 西南農業學報, 2022, 35(4): 957-963.
[34] 馬廣強, 徐紅波, 朱金華, 等. 植物內生菌對道地藥材道地性的影響 [J]. 江西中醫藥大學學報, 2014, 26(1): 56-58.
[35] 文才藝, 吳元華, 田秀玲. 植物內生菌研究進展及其存在的問題 [J]. 生態學雜志, 2004, 23(2): 86-91.
[36] Munteanu I G, Apetrei C. Analytical methods used in determining antioxidant activity: A review [J]., 2021, 22(7): 3380.
[37] SIMIC M G. Mechanisms of inhibition of free-radical processes in mutagenesis and carcinogenesis [J]., 1988, 202(2): 377-86.
Study of endophytic fungi diversity ofbased on high-throughput sequencing method and tissue isolation method and their antioxidant activities
CHEN Mei-qi1, WANG Xing1, LI Jun-yan1, TAN Qi-wen2, LI Jian-bin1, YAO Hua-xiong1, ZHANG Yu-xin1, XIE Si-yun1, DENG Zu-jun1
1. School of Life Sciences and Biopharmaceutics, Guangdong Pharmaceutical University / Guangdong Provincial Key Laboratory of Pharmaceutical Bioactive Substances, Guangzhou 510006, China 2. Guangzhou Institute of Microbiology, Guangzhou 510663, China
To analyze the taxonomic structure and diversity of endophytic fungi of Huercao () by high-throughput sequencing technology and traditional culturing techniques, and screen the endophytic fungi strains with antioxidant activity, which laid the foundation for the utilization of the endophytic fungi ofand also provided microbial resources for screening of natural antioxidants.The diversity of endophytic fungi inwas analyzed using Illumina Miseq high-throughput sequencing technology and tissue isolation method. The scavenging rate of endophytic fungi on DPPH, OH and ABTS was detected by chemical method for screening the endophytic strains with antioxidant activity.The diversity of endophytic fungi inbased on high-throughput sequencing showed that the endophytic fungi ofbelonged to 347 genera. The(15.68%) was the dominant genera in roots, and the dominant taxa in stems was(9.57%), while the dominant genera in leave was(5.33%). A total of 381 endophytic fungi belonging to 31 genera were isolated fromusing the tissue isolation method and the main taxa were(19.42%),(10.76%) and(8.66%). Diversity indices analyzed based on high-throughput sequencing and the tissue isolation found that the diversity of endophytic fungi inroots was significantly higher than those in stems and leaves, and that the total diversity index of fungal endophytes inwas higher than those in most other medicinal plants that have been reported. The results of the antioxidant activity of 24 representative endophytic strains showed that stains with free radical scavenging rates higher than 50% for DPPH, OH and ABTS accounted for 91.67%, 79.17% and 83.33%, respectively. Strainsp. HECS10 had scavenging rates of over 75% for all of DPPH, OH and ABTS.The endophytic fungi ofhad a high diversity and contained a high proportion of antioxidant active bacterial resources, which provides an important resource library for screening natural antioxidants derived from microorganisms.
Curt.; high-throughput sequencing technology; endophytic fungi; diversity; antioxidant activities
R286.2
A
0253 - 2670(2023)06 - 1924 - 11
10.7501/j.issn.0253-2670.2023.06.025
2022-09-21
國家自然科學基金項目(31971384)
陳美琪,碩士研究生,研究方向為藥用植物微生物資源開發。E-mail: 846825414@qq.com
鄧祖軍,教授,研究方向為藥用植物內生菌多樣性與功能。E-mail: dengzujun66@163.com
[責任編輯 時圣明]
