中國熱帶-溫帶地區森林群落種序-多度的緯度梯度變化分析
2023-06-15 07:17:38賈珺婷張志明林露湘譚正洪
熱帶生物學報 2023年3期
劉 玥,楊 欣,賈珺婷,張志明,林露湘,譚正洪
(1. 海南大學 生態與環境學院,海口 570228; 2. 云南大學 生態與環境學院,昆明 650504;3. 中國科學院 西雙版納熱帶植物園,昆明 650201)
從地理梯度上看,緯度多樣性梯度格局(Latitudinal Diversity Gradients,LDG)是地球上最廣泛、最重要的生物多樣性分布格局之一[1]。生物多樣性在生態學研究中占據著重要地位,在以往研究中,植物群落多樣性沿地理梯度變化的格局,一直以來都是生物多樣性研究中的一個重要課題[2-3]。在群落生態學研究中,生物多樣性的空間分布格局常常受到各種生態梯度因素制約的影響[4]。其中,物種多度分布格局通常也會隨不同的氣候環境而發生改變。群落多樣性指數可以將群落結構數量化,進而分析出群落中的物種多度格局,在深入討論群落性質上具有進一步的闡釋意義。眾多研究學者對物種豐富度的緯度分布格局做了大量研究,有結果顯示,物種豐富度會在中緯度地區達到峰值,隨緯度梯度變化整體趨勢不明顯[5-6],然而也有研究發現,在從兩極到赤道,即隨緯度下降,物種豐富度呈顯著增加趨勢[7-9],物種多度分布格局也隨之發生相應變化。在熱帶森林群落中,中性理論模擬群落物種相對豐度、物種-面積關系和物種共存模式的效果表現得尤為突出[10-12];至溫帶森林群落,生態位分化過程在群落結構中起到了更為重要的主導作用[13]。一般來說,群落多度研究也能更全面地反映出不同群落之間的差異[14-15],若將群落的物種多度格局與多樣性指數結合分析,達到相輔相成之效,同時也對群落結構的進一步研究具有更深遠的意義。本研究以中國境內溫帶跨越至熱帶的4 個森林群落為研究對象,選取3 種種多度分布模型,對樣地內木本植物群落物種多度曲線進行擬合,試圖闡述木本植物群落物種多度格局隨緯度梯度的變化規律,探討各物種多度分布模型的擬合優度,探討群落結構形成的生態學過程,以期對緯度梯度上的森林物種多樣性維持機制有進一步了解。
1 材料與方法
1.1 研究區概況本研究選擇了位于國內溫帶至熱帶具有代表性的4 個樣地,分別為溫帶地區的長白山(Changbai Mountains, CBS)、中亞熱帶地區的古田山(Gutian Mountain, GTS)、南亞熱帶的鼎湖山(Dinghu Mountain, DHS)和熱帶北緣地區的西雙版納(Xishuangbanna, XSB)20~25 hm2不等的固定監測大樣地,各固定監測樣地的具體情況如下:
(1)吉林省長白山闊葉紅松林25 hm2樣地,位于42°N,植物群落是以紅松(Pinus koraiensis)為主的針闊混交林,偶混有亞熱帶和亞寒帶的植物成分,群落整體具有明顯的成層現象和優勢樹種。該區域內主要受季風影響,會形成溫帶大陸性氣候,年平均氣溫 3.6℃,年平均降水量700 mm,降水多集中在夏季,夏季短暫且較溫暖,冬季漫長且寒冷。該植物群落內土壤分布多為山地森林土,顏色多呈暗棕色[16-17]。
(2)浙江省古田山亞熱帶常綠闊葉林24 hm2樣地,位于29°N,植物群落是以黃山松(Pinus taiwanensis)針闊混交林為主,屬于亞熱帶季風氣候區。年平均氣溫為 15.3℃,最高氣溫常出現在 7月,為 38.1℃,最低氣溫出現在 1 月,為-6.8℃,年均降雨量可達 1 963.7 mm。在夏季因受季風影響,降水充沛,冬季季風因被山體遮擋,氣候會稍顯干燥,故容易形成冬季溫暖、夏季涼爽濕潤的氣候特點。該植物群落中土壤因地形環境較復雜,沿海拔從下至上主要包括紅壤、黃紅壤、紅黃壤和沼澤土4 類,其中,紅壤、黃紅壤、紅黃壤垂直分布于海拔 500~1 000 m,而沼澤土則多分布于地形低洼地帶[18 - 19]。
(3)廣東省鼎湖山南亞熱帶常綠闊葉林20 hm2樣地,位于24°N,植物群落垂直結構比較復雜,植被具有較為顯著的地上成層現象,是北回歸線附近保存較為完整的地帶性植被。該區域屬于亞熱帶向熱帶的過渡區域,為南亞熱帶季風濕潤型氣候,夏季和冬季氣候交替明顯,年平均氣溫為20.9℃,最熱月常為7 月,平均氣溫為28.0℃,最冷月為1 月,平均氣溫為12.6℃,年降雨量為1 985 mm,且多集中于4—9 月,多伴有臺風。在海拔300 m 以下的大部分丘陵地帶多分布著赤紅壤,并且土層厚度可達到40~80 cm;黃壤多見于海拔300~900 m 地區,土層厚度40~90 cm,當海拔高度超過900 m,多為山地灌叢草甸土,其厚度較薄,約20~30 cm[20]。
(4)云南省西雙版納熱帶季節雨林20 hm2樣地,位于21°N,森林類型屬熱帶季節雨林植被亞型,植物群落類型中多以望天樹(Parashorea chinensis)為優勢,屬于北熱帶季風氣候,常年受印度季風影響,氣候溫暖濕潤,具有較強的季節性,且干濕季明顯,年平均氣溫為 21.7℃,最高溫常出現在6 月,最低溫出現在1 月,年平均降雨量達1 531.9 mm,5—10 月為雨季,11 月至翌年4 月為干季。群落內的土壤分布類型多為磚紅壤[21-24]。
1.2 研究方法本研究所選樣地均嚴格按照CTFS(Centre for Tropical Forest Science)[25]的技術標準建設,并對樣地內 DBH ≥ 1 cm 的木本植物進行統計分析,計算各樣地群落內的α多樣性和β多樣性,分析各樣地中整體群落的物種多度分布格局;采用頻度定義的方法[26-27],將各群落中頻度大于25%的物種定義為常見種,而頻度小于5%的物種定義為稀有種,并分析群落中常見種和稀有種各自的種多度分布格局。
1.2.1 群落多樣性指數(1)多樣性指數常作為群落結構種的一種量化指標,可用于反映群落中的組成結構,也可為研究群落中各物種或個體分布之間相關關系提供系統性理解。基于各群落種實測所得的樹木個體數據,α多樣性指數的計算公式如下:
Shannon-Wiener 多樣性指數[28]:
Simpson 優勢度指數[29]:
式中,Pi為群落中各植物種的重要值,S為群落中的物種數。
(2)Bray-curtis 相異度常用于衡量不同群落間物種組成的差異度,可基于群落中不同物種組成的數量特征如多度、重要值等進行計算,公式如下:
式中,k是物種數,y表示物種多度,i和j表示相比較的2 個樣地[30]。
1.2.2 種多度分布模型(1)純統計模型對數級數模型:
式中,S(n)為群落中多度為n的物種數,α代表群落內物種多樣性;X為常數(0<X≤1)[31]。
對數正態模型:
式中,Ai為第i個物種的多度,μ和δ分別表示正態分布的均值和方差,Φ為正態偏差[32]。
(2)生態位理論模型
生態位優先占領模型:
式中,模型假定種 1 先占用群落總生態位的k份,種 2 占用剩下的k(1-k)份,種 3 再繼續占據余下的k(1-k)2份,依次類推,直至剩余資源無法繼續維持另一個物種的生存[33]。
斷棍模型:
式中,該模型假設某一群落內的總生態位是長度為 1 的一條短棍,若將其分割為S段,則代表生態位被S個物種所占有,模型假定S個物種的分類地位和競爭能力均相似,且在群落中同時出現。J表示群落內物種的個體總數,Ai表示第Ai物種的多度[34]。
(3)中性理論模型
復合群落零和多項式模型:
式中,S(n)為群落中多度為n的物種數,θ代表群落內物種多樣性;J為取樣點內個體數[35]。該模型假定某一群落的物種多度分布由自中性復合群落的隨機漂變過程主導。
Volkov 模型:
式中,相較于復合群落零和多項式模型增加了遷移系數m,S(n)為群落中多度為n的物種數,γ表示遷移到局域群落內的個體數[36]。
1.2.3 模型擬合優度檢驗本研究將采用K-S檢驗[37-38]對模型的擬合效果進行驗證,同時還利用赤池信息準則(Akaike information criterion, AIC)來比較模型擬合的優劣。K-S檢驗是一種常用的非參數檢驗方法,它通過計算2 個樣本經驗分布函數(Empirical distribution function)之間的距離(統計量D),以此來判斷二者是否存在顯著差異。為了避免K-S檢驗中常發生的數據連結問題,在處理數據時通過優化 bootstrap,之后再進行K-S檢驗(其中,bootstrap 的次數為 1 000 次,在所得結果中,若當P<0.05 時,則表明該模型被拒絕)[39]。同時還會利用赤池信息準則(Akaike information criterion,AIC)對各模型的擬合優度進行判斷,在AIC檢驗中,若AIC的數值越小,則表示該模型擬合效果越好。
以上數據處理分析及優度檢驗均基于 R-4.2.1軟件(R Core Team, 2022),其中模型擬合使用 sads軟件包完成;基于 bootstrap 的K-S檢驗利用Matching 軟件包完成。
2 結果與分析
2.1 不同緯度各群落多樣性分布格局從圖1中可以看出,隨緯度的上升,森林群落中α多樣性呈現逐漸減小的趨勢,在熱帶森林群落中,α多樣性最高,在溫帶森林群落中則最小。其中Shannon-Wiener 指數與Simpson 指數的變化趨勢基本保持一致。在β多樣性方面,從圖2 可以看出,沿緯度梯度,Bray-curtis 相異度逐漸增大,且在位于低緯度森林群落中,熱帶森林群落與緯度相鄰的亞熱帶森林相異度小于其余緯度梯度群落之間的相異度。
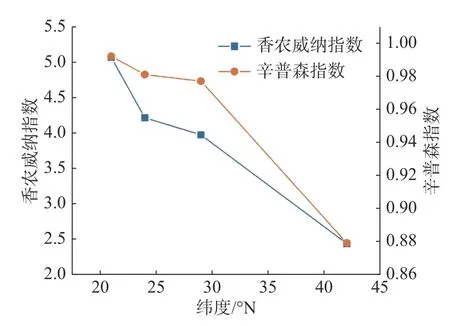
圖1 緯度梯度群落α 多樣性變化特征
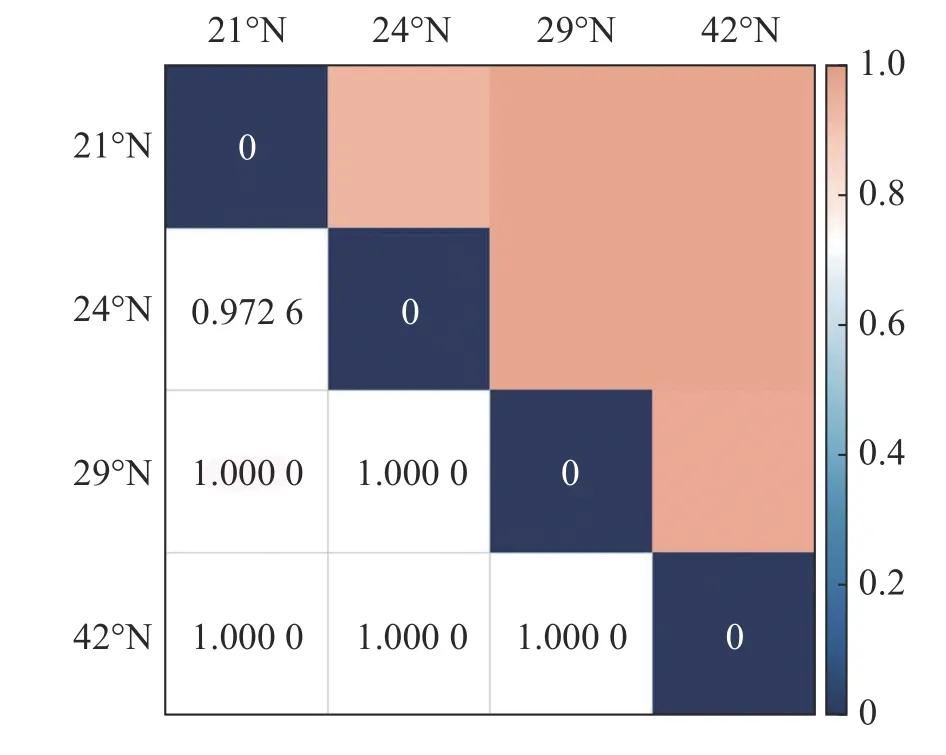
圖2 緯度梯度各群落 Bray-curtis 相異性
2.2 沿緯度梯度常見種、稀有種變化情況從統計結果可以看出(圖3,表1),常見種物種數變化趨勢與群落內物種總數變化趨勢基本相符,即緯度越高,常見種物種數逐漸變少。但不同緯度群落中常見種種數相比較可以看出,隨緯度從赤道向北增加,常見種占總樹種數分別為 14.5%、24.6%、31.5%、44.2%,呈增長趨勢。從稀有種變化情況可知,沿緯度梯度,稀有種數目變化趨勢與群落內物種總數變化趨勢一致,即緯度越高,稀有種數目變少;從不同緯度群落中稀有種種數相比較可看出,隨緯度從赤道向北增加,稀有種占總樹種數分別為 60.6%、56.4%、52.8%、28.8%,也呈減少趨勢。
表1 緯度梯度各群落樹種數情況

圖3 各群落中常見種和稀有種所占百分比沿緯度梯度變化情況
2.3 不同緯度下整體群落種多度分布擬合優度檢驗結果通過統計分析4 個樣地實測數據(表1)可發現,緯度從南至北,群落內物種數有逐漸減少的趨勢,熱帶西雙版納森林群落中的物種數最多,有 469 種,物種數最少為溫帶長白山森林群落,只有 59 種,利用6 個物種多度分布模型擬合結果表明(圖4,表2~3):在較低緯度的西雙版納(21°N)和鼎湖山(24°N)群落中,生態位優先占領模型無法通過K-S檢驗(P< 0.05),西雙版納和鼎湖山群落的物種多度分布可由對數級數模型、對數正態模型、斷棍模型、復合群落零和多項式模型和 Volkov 模型擬合,其中均顯示出復合群落零和多項式模型和對數級數模型的擬合效果最優,西雙版納群落種多度模型擬合效果排序為:復合群落零和多項式模型 > 對數級數模型 > 對數正態模型 > Volkov 模型 > 斷棍模型,鼎湖山群落種多度模型擬合效果排序為:復合群落零和多項式模型 > 對數級數模型 > Volkov 模型 > 對數正態模型 > 斷棍模型。
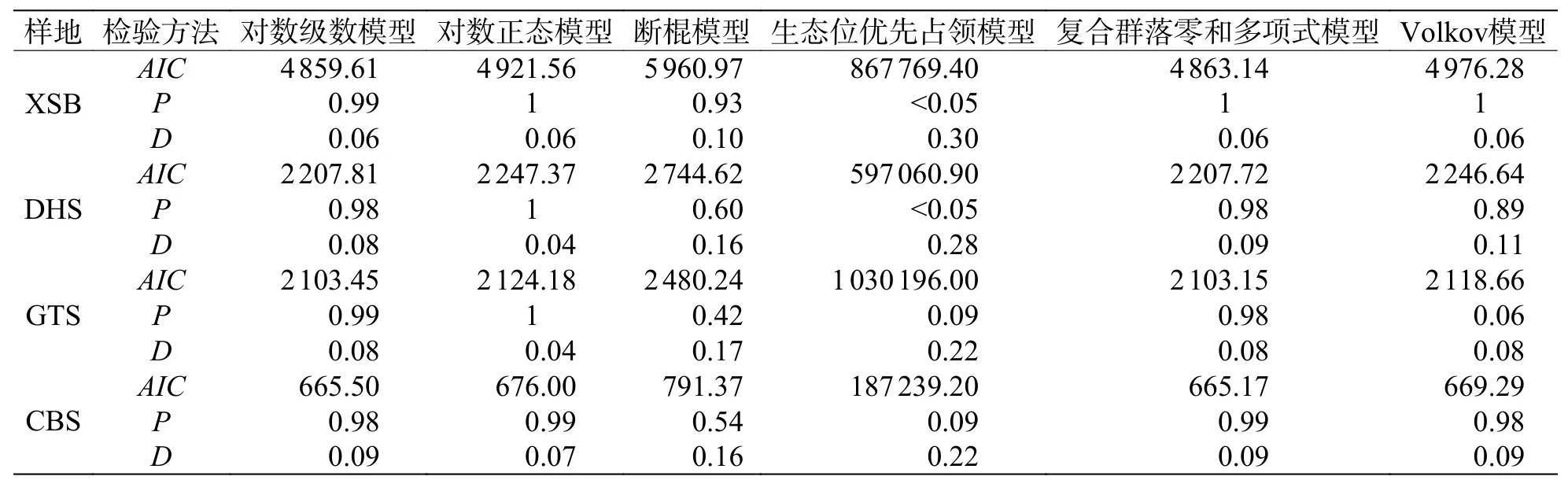
表2 緯度梯度上群落整體物種多度模型擬合優度檢驗結果
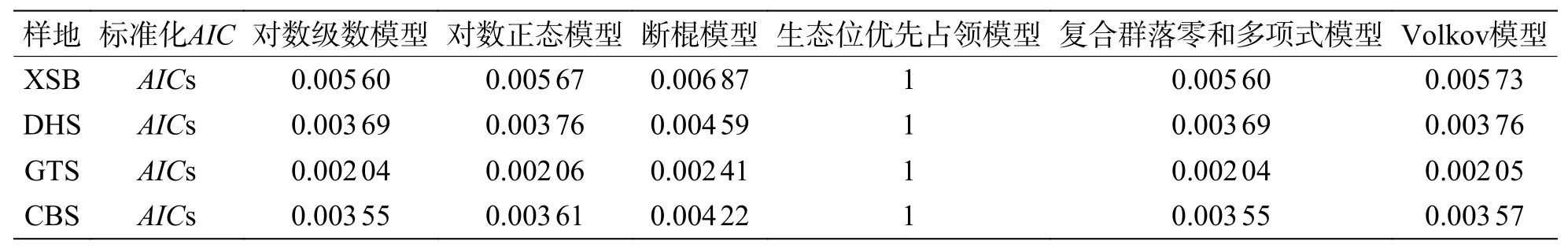
表3 整體物種種多度分布模型AIC 值標準化
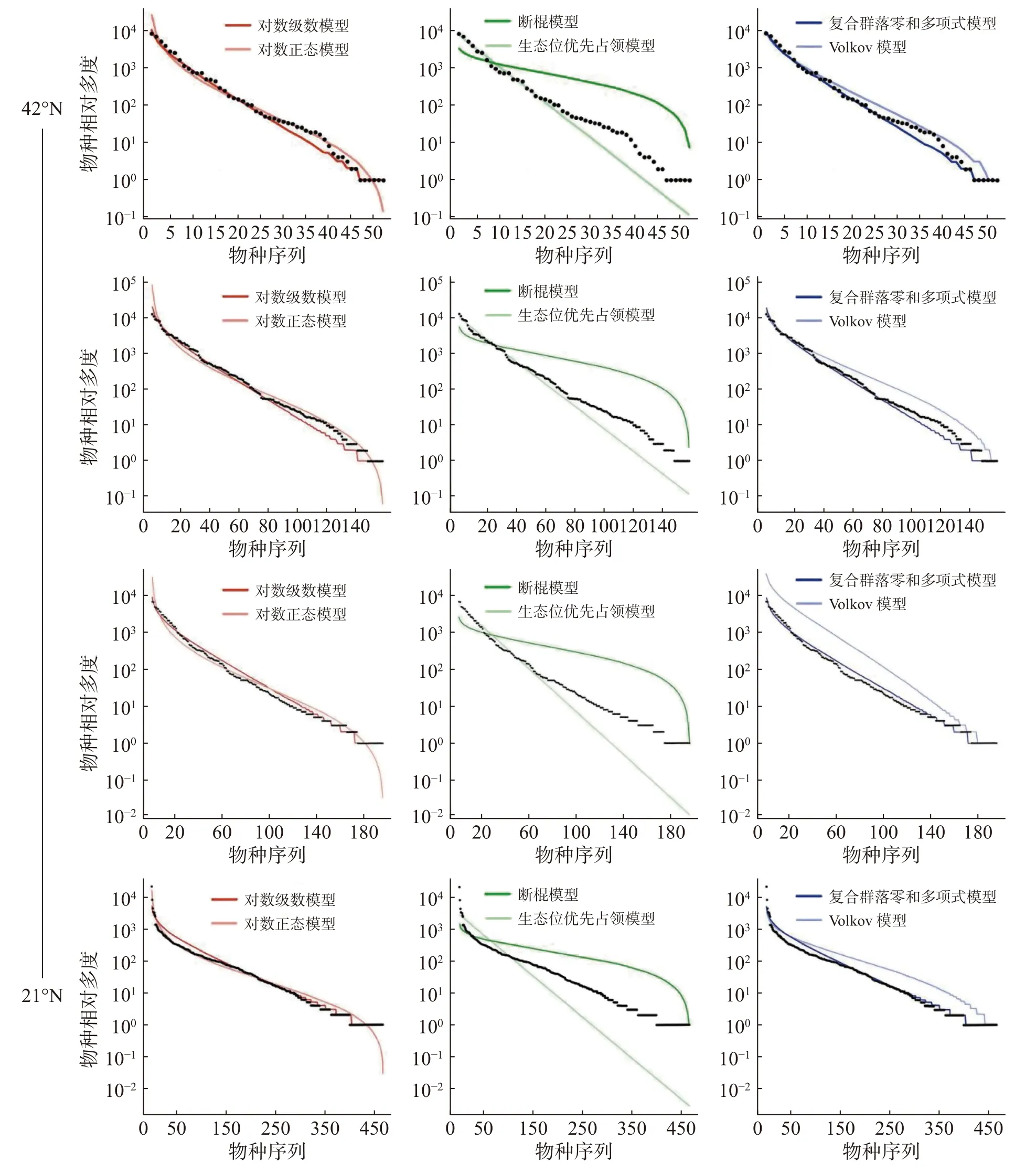
圖4 緯度梯度整體群落物種多度分布模型擬合結果從上至下依次為:長白山,古田山,鼎湖山,西雙版納。
古田山(29°N)和長白山(41°N)群落中物種多度擬合結果顯示所有模型都被接受(P> 0.05),種多度分布模型擬合優度排序為:復合群落零和多項式模型 > 對數級數模型 > Volkov 模型 > 對數正態模型 > 斷棍模型 > 生態位優先占領模型,其中生態位優先占領模型的擬合效果依然最差。
沿緯度梯度,6 個模型的擬合優度在各樣地中的檢驗結果均顯示中性理論模型對物種多度分布格局的擬合效果優于生態位理論模型,且其中復合群落零和多項式模型擬合結果均為最佳,除生態位優先占領模型之外,其他5 種模型均可以擬合不同緯度下的物種多度分布,且擬合效果會隨緯度上升變得更好。
2.4 不同緯度下植物群落常見種的模型擬合優度比較在常見種的擬合效果上(圖5,表4),所有模型均被接受(P> 0.05),其中生態位理論模型中的生態位優先占領模型的擬合效果較差,所有群落中6 個模型擬合效果排序均為:對數正態模型 >斷棍模型 > Volkov 模型 > 復合群落零和多項式模型 > 對數級數模型 > 生態位優先占領模型。
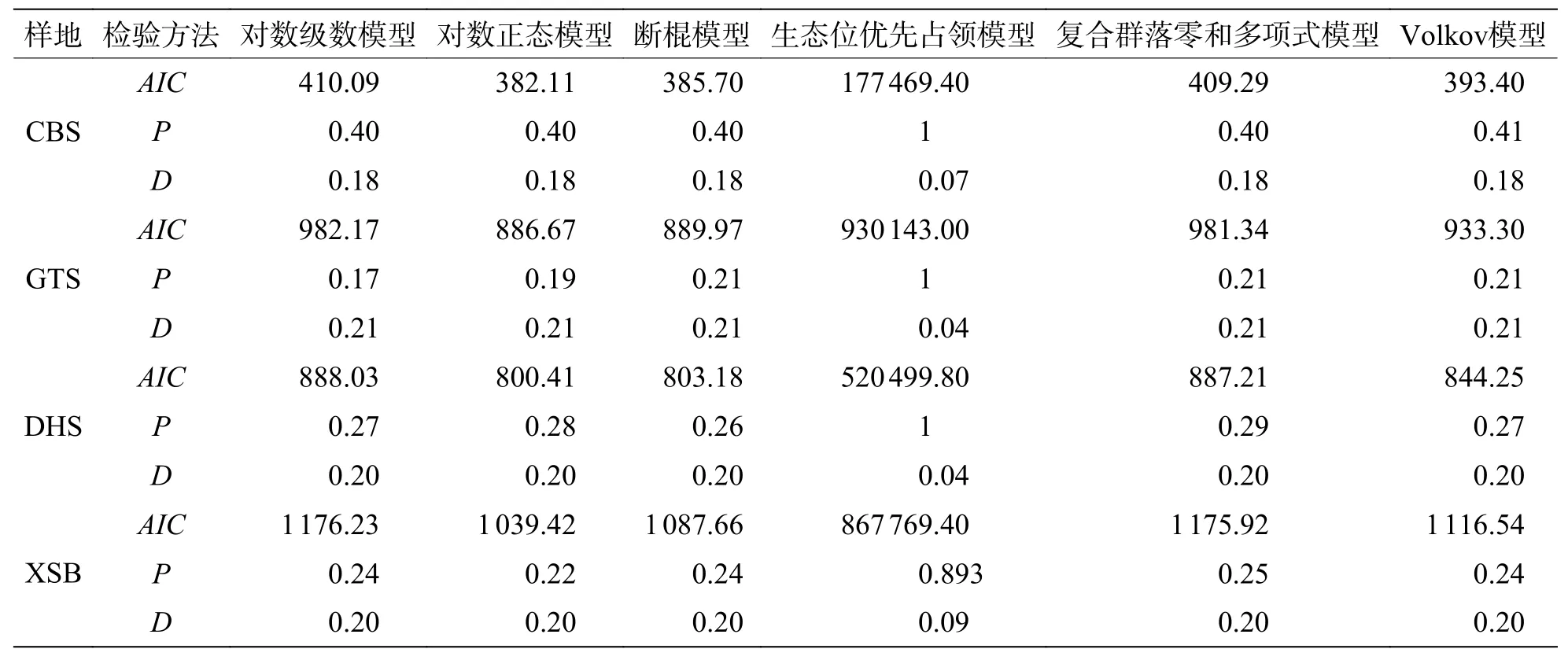
表4 緯度梯度上群落中常見種物種多度模型擬合優度檢驗
通過標準化之后的AIC可更直觀地發現(表5),在常見種的種多度分布擬合中,生態位理論模型中的斷棍模型較中性理論模型擬合效果更優,且對數正態模型擬合效果優于對數級數模型,沿緯度從赤道向北增加,斷棍模型對群落中常見種種多度分布格局的擬合效果均表現最好,除古田山群落(29°N)之外,斷棍模型的擬合優度也呈逐漸增加趨勢。
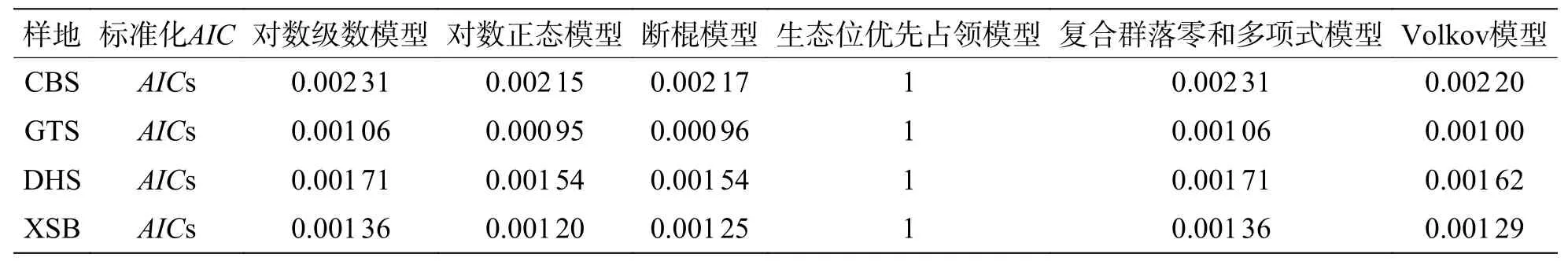
表5 常見種模型擬合AIC 值標準化
2.5 不同緯度下植物群落稀有種的模型擬合優度比較在稀有種的擬合效果上(圖6,表6),所有模型均被接受(P> 0.05),生態位理論模型中的生態位優先占領模型擬合效果較差,西雙版納群落模型擬合效果排序為:復合群落零和多項式模型 >對數級數模型 > Volkov 模型 > 對數正態模型 > 斷棍模型 > 生態位優先占領模型;鼎湖山群落模型擬合效果排序為:Volkov 模型 > 復合群落零和多項式模型 > 對數級數模型 > 對數正態模型 > 斷棍模型 > 生態位優先占領模型,古田山群落模型擬合效果排序為:對數級數模型 > Volkov 模型 > 斷棍模型 > 復合群落零和多項式模型 > 對數正態模型 > 生態位優先占領模型,長白山群落模型擬合效果排序為:復合群落零和多項式模型 > 對數級數模型 > Volkov 模型 > 斷棍模型 > 對數正態模型 > 生態位優先占領模型,總體上,中性理論模型對稀有種的擬合效果更佳,且對數級數模型擬合效果優于對數正態模型。通過標準化AIC也可看出(表7),沿緯度梯度,中性理論模型對群落中稀有種種多度分布的擬合效果均更好。
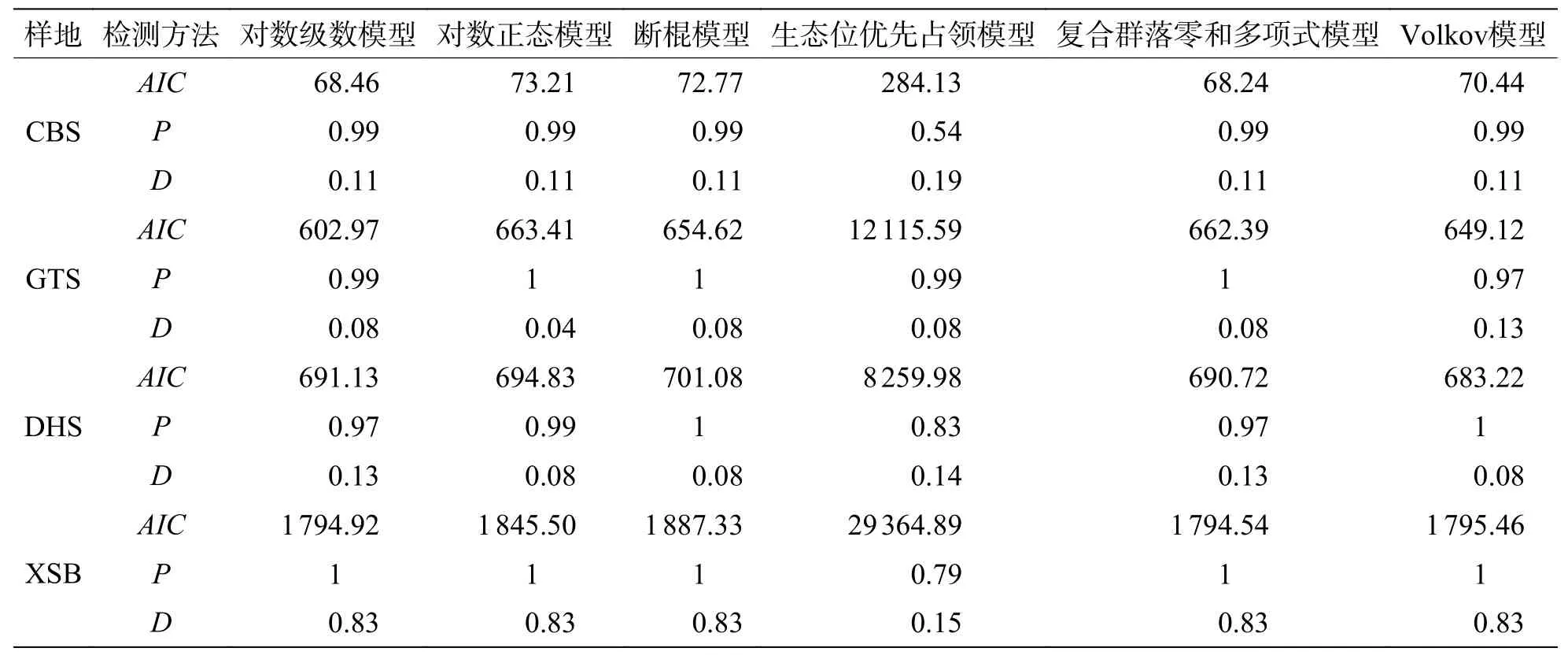
表6 緯度梯度上群落中稀有種物種多度模型擬合優度檢驗
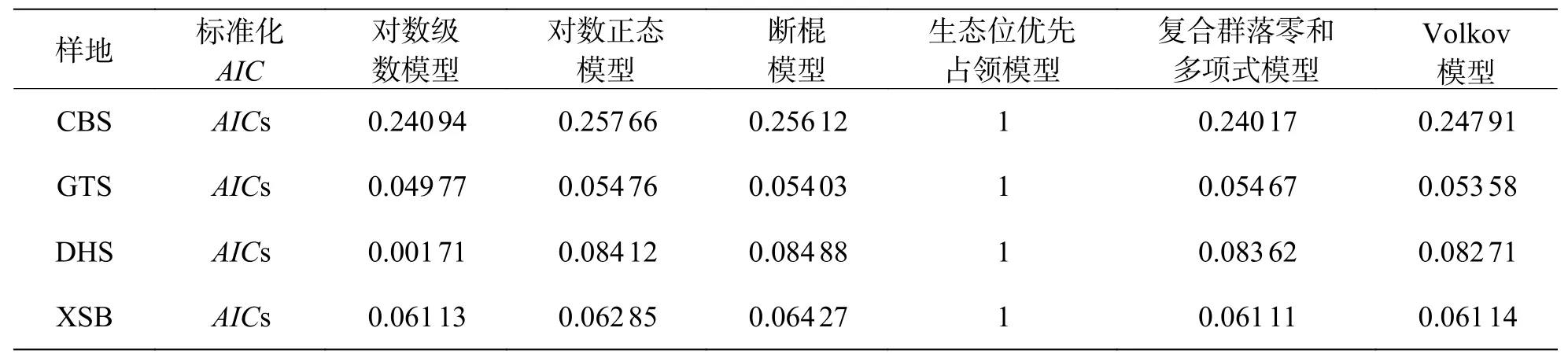
表7 稀有種模型擬合AIC 值標準化
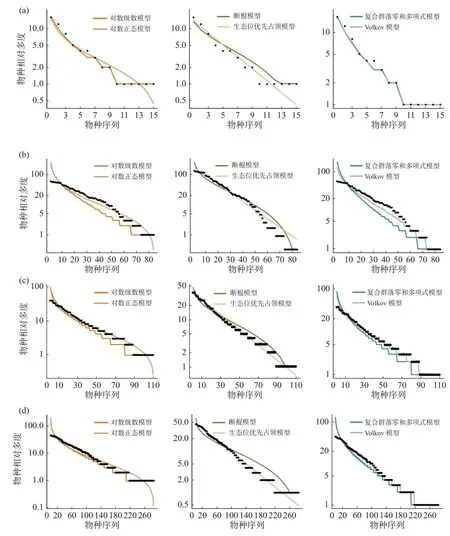
圖6 緯度梯度上群落中稀有種物種多度分布擬合結果
3 討 論
3.1 沿緯度梯度群落多樣性的變化特征從熱帶跨越至溫帶,物種之間的相互作用,以及環境因子的影響都會使群落向不同的結構組成發展進化,使得植物群落的組成結構出現差異化。眾多研究表明沿赤道向更高緯度發展,植物物種豐富度和多樣性通常被認為呈逐漸下降趨勢[40],在本研究結果中也出現了群落中的物種豐富度向赤道方向逐漸增加的相似變化趨勢,在離赤道較近的低緯度西雙版納森林群落因為氣候季節間和年際間變化較小,群落物種多樣性較高,亦或因其群落進化時間較長,環境條件更加穩定,故物種豐富度也會更高;鼎湖山森林群落因緯度位置處于亞熱帶與熱帶過渡區[20],熱帶物種豐富,所以物種多樣性會略高于同為亞熱帶群落的古田山植物群落,同樣也因緯度與西雙版納森林群落較接近,具有一定數量的共有種,故群落之間的相異度較低;而緯度位置較高的長白山森林群落氣候環境較特殊,自然選擇主要受物理因素控制,且具有廣適性的植物更容易生存,故群落內物種數下降且多樣性較小。
關于物種多樣性沿緯度梯度分布格局有很多假說試圖解釋[41],如“競爭假說”[42-43]、“環境異質性假說”[44]、“生態位假說”[45]等,其中眾多假說中所涉及到的生態位分化理論和群落中性理論是目前從物種相似度的視角來闡述物種共存和物種多樣性維持的兩種主要理論,大部分的研究結果表明,在低緯度熱帶地區,群落中的物種多樣性較高,無法通過生態位分化來形成相應的群落結構,故群落中物種多度的變化更傾向于受隨機過程的可能性較大[46-48];沿緯度升高,群落中物種多樣性較低緯度群落低,可供分配的資源充足,生態位分化得更明顯。
3.2 整體群落物種多度分布模型擬合優度隨緯度變化特征物種多度模型擬合效果方面,在本研究緯度梯度上,模型擬合結果顯示4 個緯度梯度群落中均為中性理論模型擬合效果最佳,其次為對數級數模型。中性理論包括了2 種物種相對多度的分布情況,一是在集合群落中,通過點突變形成新物種的對數級數分布;其二是局域群落中,因受擴散限制而隨機產生新物種的零和多項式分布,且此分布模式的解釋度在熱帶雨林群落中較高[49]。從此結果來看,零和多項式模型與其他眾多模型相比,具有擬合曲線的尾部較長的特點,結合熱帶群落中物種多度情況,也能更好地說明該模型在低緯度的熱帶群落中擬合優度更高。
物種多度模型擬合的結果通常具有一定的尺度效應[50],不同尺度的群落中主導的生態學過程可能不同,故模型擬合的結果也會存在差異[51]。以往研究發現,隨著研究群落樣地尺度的增大,最優的擬合模型可能會發生變化,當選樣尺度越小(40 m×40 m 或 60 m×60 m 以下),生態位理論模型擬合效果越好[52],而中性理論模型在大尺度(100 m×100 m)群落的物種多度格局的擬合方面往往會優于生態位模型,能更好地預測群落植物物種多度分布[53-54]。綜上,也能更好地解釋在此森林群落中中性理論模型擬合效果更優。在本研究中較高緯度的長白山群落物種多樣性顯著下降,但依然顯示出復合群落零和多項式模型和對數級數模型擬合效果較好。
3.3 常見種、稀有種物種多度分布模型擬合優度隨緯度變化特征過往研究中,常見種與稀有種的多度分布格局同群落整體多度分布的模式一直存在爭議:Vazquez 等[55-56]發現,常見種的多度分布通常與群落整體多度分布模式保持一致;但也有學者在研究中發現植物群落中的稀有種對整體物種多樣性及多度的貢獻度高于非稀有種[57]。本研究得出的結果與此相似,緯度沿赤道向北增加,常見種和稀有種種樹同群落中總樹種數的變化趨勢一致,均為逐漸減少,但其各自占總樹種數的百分比有所差異:稀有種所占百分比隨緯度增大而減少,而常見種所占百分比卻隨緯度增大而增大;從模型擬合效果來看,低緯度至高緯度地區,在常見種群落中斷棍模型均具有較好的擬合效果,而對稀有種物種多度的擬合則表現出中性理論模型效果更佳,且稀有種種多度分布最優擬合模型與群落整體種多度分布最優模型一致,均為中性理論模型。由此可說明,常見種種多度分布格局與群落中的資源分配有關,而稀有種的種多度分布格局形成過程更傾向于生態位等價下,具有隨機性的物種生態漂變[58],且稀有種的種多度分布可能是群落整體物種多度分布格局形成的主導因素。
對于群落中的稀有種來說,不同緯度上中性理論模型擬合的效果有所差別。復合群落零和多項式模型對低緯度的西雙版納群落和較高緯度的長白山群落中稀有種的種多度分布格局擬合效果更好,而增加了密度限制因子的 Volkov 模型卻對位于中等緯度的古田山森林群落和鼎湖山森林群落中稀有種種多度分布格局擬合更優。因為不同群落的分布格局往往會有不同的形成機制,如遺傳漂變和物種遷移等基本生態過程[59]均會導致向中性理論模型分布,且在群落的不同的生活史階段,生態位與中性過程的相對重要性是在不斷變化的[60],同時也有研究發現,稀有種在群落中會具有較高的喪失率和遷移率[51]。基于此,可猜測在本研究中不同緯度群落中稀有種的種多度格局形成過程存在差異。
沿緯度上升,常見種多度分布模型擬合結果呈現出對數正態模型和斷棍模型效果最優,雖然與群落整體物種多度分布結果不一致,但群落中常見種占群落總樹種數百分比卻呈增加趨勢。一般來說,在群落中常見種會比稀有種具有更高的穩定性[61],在低緯度的群落中,物種多樣性高,在資源分配方面,常見種個體在群落中往往占據的資源更多,故導致常見種種樹數會低于稀有種;而在高緯度地區中的常見種因能適應嚴苛的環境,逐漸成為了緯度梯度上的特化種,在高緯度的環境中占據較高的生態位,故群落中物種種樹會高于稀有種,且占比高于稀有種。故結合中性理論模型的擬合特點分析,在高緯度的群落中,起主導作用的生態學過程不能單從稀有種的種多度模型擬合效果看,關于常見種和稀有種在群落中物種多度分布方面的研究還亟待更深入的探究。
本研究對緯度21°~40°N 范圍內森林群落種多度分布格局進行研究,結果發現:緯度沿赤道至北,群落內物種數有逐漸減少的趨勢,低緯度群落中植物物種數最多,高緯度群落物種數最少。且沿緯度梯度的未發現物種多度格局分布的明顯差異,但不代表各群落中的生態過程相同,結果表明在各森林群落中種多度分布模型擬合優度檢驗結果均顯示出中性理論模型對現階段物種多度分布格局的擬合效果優于生態位理論模型,除生態位優先占領模型之外,其余模型均可以擬合不同緯度下的物種多度分布,且擬合效果隨緯度上升變得更好;森林群落中常見種和稀有種物種數變化趨勢與群落內物種總數變化趨勢基本符合,即緯度越高,物種數逐漸變少,但常見種在總樹種數中的占比呈增長趨勢,斷棍模型對群落中常見種種多度分布格局的擬合效果均表現最好;而稀有種數占總樹種數比值呈逐漸減小趨勢,中性理論模型對稀有種種多度分布的擬合效果均更好。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-11-30 02:58:01
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
攝影之友(影像視覺)(2019年2期)2019-03-05 08:27:14
汽車觀察(2018年10期)2018-11-06 07:05:26
中華詩詞(2018年11期)2018-03-26 06:41:34
Coco薇(2016年8期)2016-10-09 02:11:50
