種齡及接種量協同作用對纖維素酶活性的影響
2023-07-04 04:46:50賈洪柏許超劉軍何凡丁宏云
安徽農業科學 2023年6期
賈洪柏 許超 劉軍 何凡 丁宏云

摘要[目的]分析菌種種齡和接種量的協同作用對纖維素酶活性的影響,為生產纖維素酶奠定理論基礎。[方法]分別對單菌種(DQ1)和混合菌種(DQ1+DQ2)不同種齡和接種量進行發酵培養試驗,測定各處理的纖維素酶活力,分析不同處理纖維素的變化趨勢及差異。[結果]在培養過程中,不同種齡的纖維素酶活力都在第8天達到最大;單菌種接種量為1%和2%時種齡4的纖維素酶活力最高,分別達到19.57和20.50 IU/mL;混合菌種接種量為1%和2%時種齡1的纖維素酶活力最高,分別為21.21和20.82 IU/mL。在第8天時,相同菌種和接種量的不同種齡的纖維素酶活力有一定差異。[結論]采用該研究的單菌或混合菌生產纖維素酶時,最優接種條件是種齡4、接種量2%,可以獲得較高的纖維素酶活力,并且生產成本較低。
關鍵詞 種齡;接種量;協同作用;纖維素酶;活性
中圖分類號 X172 文獻標識碼 A 文章編號 0517-6611(2023)06-0001-03
doi:10.3969/j.issn.0517-6611.2023.06.001
Synergistic Effects of Seeding Age and Inoculum Size on Cellulase Activity
JIA Hong-bai1,2,XU Chao1,LIU Jun1 et al
(1.Anji Guoqian Environmental Technology Co., Ltd., Huzhou, Zhejiang 313300;2.College of Life, Northeast Forestry University, Harbin, Heilongjiang 150040)
Abstract [Objective]To analyze the effect of the synergistic effect of strain seeding age and inoculation size on cellulase activity and lay a theoretical foundation for cellulase production.[Method]The fermentation experiments of single strain (DQ1) and mixed strain (DQ1 + DQ2) with different seeding ages and inoculum sizes were carried out, and the change trend and difference of cellulase activities were analyzed.[Result] During the culture process, the cellulase activity of different ages reached its maximum on the 8th day.The cellulase activity of seeding age 4th reached the highest at 1% and 2% inoculation of single strain, they were 19.57 and 20.50 IU/mL, respectively. And the cellulase activity of seeding age 1st reached the highest at 1% and 2% inoculation of single strain, they were 21.21 and 20.82 IU/mL. At the 8th day, the cellulase activity of different seeding ages of the same strain and inoculation size had certain differences.[Conclusion] When using the single or mixed strain in this study to produce cellulase, the optimal inoculation conditions are seeding age 4th, inoculation size 2%, which can obtain higher cellulase activity and lower production cost.
Key words Seeding age;Inoculum size;Synergistic effect;Cellulase;Activity
我國作為農業大國,每年都會產生幾億噸的秸稈,如果不能良好地利用秸稈資源,則會造成環境污染,影響人類健康[1-2]。利用秸稈等纖維素原料生產燃料乙醇是目前研究的熱點[3],其中利用纖維素酶將纖維素轉化為小分子有機物,是目前利用纖維素類物質的有效措施[4-7]。燃料乙醇生產中存在的最大問題是如何降低生產成本,其中如何提高纖維素酶的產量是降低生產成本的一個重要因素。近年來,學者們從多方面對產纖維素酶的微生物進行了研究,包括產纖維素酶菌種的篩選[8-11]、微生物菌種的誘變,如常溫常壓等離子誘變[12]、紫外線誘變[13]、亞硝基胍誘變[14]等,以及利用分子手段提高微生物產纖維素酶[15-16]等,并且取得了較好的結果。
發酵工藝的發展最終都是為了提高發酵過程產物的產量。種子發酵階段的優化被認為是最后發酵階段優化提高培養物產量的補充。有大量的文獻已經報道了發酵最后階段優化提高最終發酵產物的例子[17-19]。筆者針對液體種子種齡和接種量對纖維素酶(CMC)活性的影響進行了研究,旨在為此方面的研究和生產奠定理論基礎。
1 材料與方法
1.1 試驗材料
1.1.1 菌種。
利用富集培養的方法從大慶野外分離得到的2株產纖維素酶鐮刀菌屬DQ1和DQ2。
1.1.2 柳枝草。
采自東北林業大學大慶生物技術研究院的柳枝草種植基地。
1.1.3 藥品。
購自天津市大茂化學試劑廠的分析純試劑。
1.1.4 纖維素分解菌初篩分離平板培養基。
KH2PO4 2.0 g,(NH4)2SO4 1.4 g,MgSO4·7H2O 0.3 g,CaCl2 0.3 g,FeSO4·7H2O 0.05 g,MnSO4 1.6 mg,ZnCl2 1.7 mg,CoCl2 0.2 mg,蒸餾水1 000 mL,pH為5.5~6.0,以濾紙為唯一碳源。
1.1.5 羧甲基纖維素鈉復篩選培養基。
CMC-Na 20 g,NaHPO4 2.5 g,KH2PO4 1.5 g,蛋白胨2.5 g,酵母膏0.5 g,蒸餾水1 000 mL,瓊脂20 g,pH為7.0~7.2。
1.1.6 牛肉膏蛋白胨分離培養基。
牛肉膏3 g,蛋白胨10 g,NaCl 5 g,瓊脂15 g,蒸餾水1 000 mL,pH為7.0~7.2。
1.1.7 柳枝草液體培養基。
KH2PO4 2.0 g,(NH4)2SO4 1.4 g,MgSO4·7H2O 0.3 g,CaCl2 0.3 g,FeSO4·7H2O 0.05 g,MnSO4 1.6 mg,ZnCl2 1.7 mg,CoCl2 0.2 mg,蛋白胨 0.5 g,尿素0.3 g,堿處理后的柳枝草粉10 g,蒸餾水1 000 mL,pH為5.5~6.0。
柳枝草粉的制備:將柳枝草剪成2~3 cm的小段,用2%的NaOH溶液浸泡24 h;用流水沖洗后,將pH調至中性,烘干;將烘干的小段粉碎至60目備用。
1.1.8 種子液體培養基。
同柳枝草液體培養基,將柳枝草粉改為羧甲基纖維素鈉。
1.2 試驗方法
1.2.1 培養方法。
向裝有150 mL種子液體培養基的2個三角瓶中一個接種菌株DQ1兩環,另一個接種菌株DQ1與DQ2各1環,在150 r/min、28 ℃條件下振蕩培養。分別于接種后的第1天、第2天、第3天、第4天、第5天取種子液按照體積比1%和2%的接種量分別接種到裝有200 mL柳枝草液體培養基中同條件下振蕩培養,同時測定種子液菌種濃度。加入菌種后第2天測定CMC活力,連續測定10 d,每個樣品重復3次。
1.2.2 酶活力測定方法。
取一定的培養液于4 ℃、4 000 r/min離心15 min,取上清液作為粗酶液測定酶活力。
CMC活力測定[20]:取經適當稀釋的酶液1 mL,加入1 mL含1% CMC的pH 4.8的醋酸-醋酸鈉緩沖液,混勻,50 ℃ 恒溫水浴30 min,加入2.5 mL DNS試劑,沸水中煮沸5 min,后用流水冷卻至室溫,定容至25 mL,在520 nm下比色,測出OD值,查閱標準曲線后,求出溶液中的葡萄糖含量。以國際單位為依據,定義每分鐘催化纖維素水解生成1 μmol 葡萄糖的酶量為一個酶活力單位IU。
DNS試劑配制:稱取200 g酒石酸鉀鈉,溶于一定量水中,加熱溶解,添加10.0 g 3,5-硝基水楊酸、10.0 g氫氧化鈉,溶解后加入2.0 g苯酚、0.5 g無水亞硫酸鈉,全部加熱溶解后,冷卻至室溫,定容至1 000 mL。用前7 d配制。
1.3 數據統計分析
利用SPSS 17.0軟件對試驗數據進行統計分析。
2 結果與分析
2.1 種子濃度的變化
從各種齡種子濃度(圖1)可以看出,無論是單菌種(DQ1)還是混合菌種(DQ1+DQ2)在培養過程中菌種濃度都是先升高后降低,在種齡3時達到最高,單菌種的濃度達到2.1×105 CFU/mL,混合菌種的濃度達到了4.9×106 CFU/mL。
2.2 CMC活力的變化趨勢
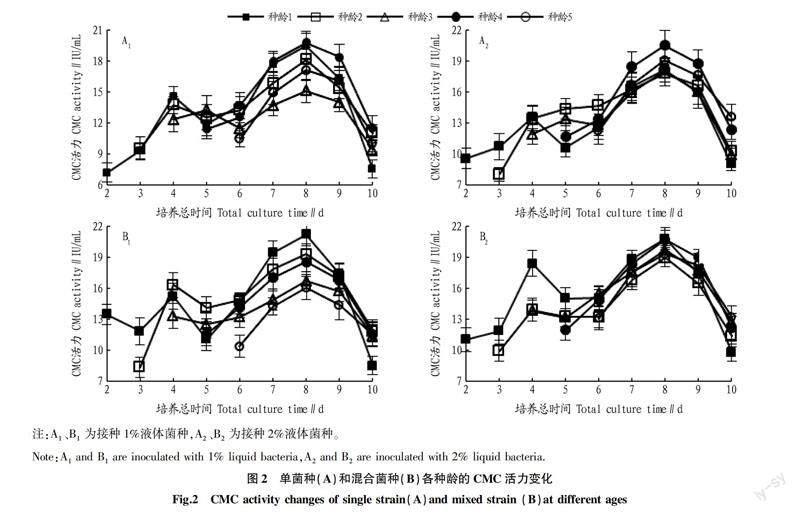
從不同菌種不同接種量的各種齡CMC活力變化趨勢(圖2)可以看出,在整個的試驗過程中,種齡1和種齡2在第4天時的CMC活力有一個小幅升高,而后降低再升高,在第8天時達到最高,然后再降低;種齡4和種齡5的CMC活力的變化趨勢都是先升高,在第8天時達到最高,而后再降低。接種單菌種種齡3的CMC活力變化趨勢與種齡1和種齡2相同,但是小幅增高的時間向后推遲到了第5天;接種混合菌種種齡3的CMC活力與種齡2的CMC活力在同一時間的變化趨勢相同,即逐漸降低,再逐漸升高,在第8天時達到最高,而后逐漸降低。
韓美玲等[21]研究發現糙皮側耳在不同碳源條件下都表現出羧甲基纖維素酶活性升高—降低—升高的過程,這與該研究中種齡1和種齡2的CMC活力變化趨勢相同。CMC活力出現小高峰可能是菌體數量較少,并且適應性弱,在接種后種子液中易利用的營養成分促進了菌種的快速生長,從而使CMC活力升高;當這部分碳源消耗盡后,隨著CMC對培養液中纖維素的作用,CMC活力降低。但隨著微生物數量的增加,CMC活力又開始升高。
2.3 不同種齡及接種量對CMC活力的影響
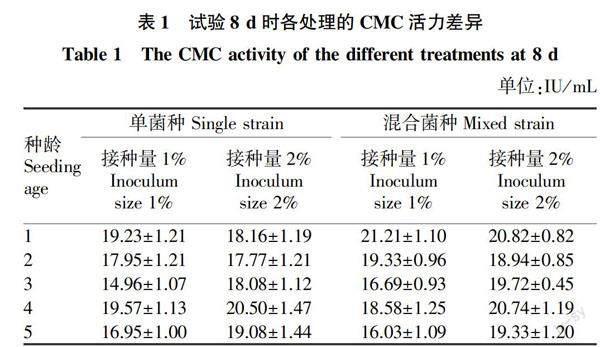
從試驗中第8天各處理的CMC活力(表1)可以看出,以單菌種接種培養的CMC活力2種接種量(1%、2%)種齡4的CMC活力都最高,分別達19.57和20.50 IU/mL。以混合菌種接種培養的CMC活力,接種量為1%時種齡1的CMC活力最高,達21.21 IU/mL;接種量為2%時種齡1的CMC活力最高,達20.82 IU/mL。經方差分析和多重比較表明,試驗第8天時,接種相同菌種和接種量的不同種齡處理的CMC活力有一定差異。從圖1~2可以看出,種齡3的種子濃度最高,但在接種培養過程中的CMC活力普遍較低,這與孫寶婷等[22]的研究結果相反。這可能是種子培養階段與擴大培養階段的培養基碳源不同導致的。接種帶來的種子液中易利用的營養成分較少,接入新的培養液后,菌種生長緩慢,大量接種的微生物死亡或停滯[23],導致酶活力較低。師璐等[24]研究發現,羧甲基纖維素鈉是作為產纖維素酶最好的碳源。而種齡1和種齡2雖然菌種生長力較弱,但接種帶來的種子液中易利用的營養成分相對較多,有利于微生物快速生長繁殖;雖然種齡4的種子液中易利用的營養成分也很少,但種子生長力強,適應性強,能很快地在新的培養液中生長繁殖,因此培養液中的CMC活力都比種齡3的CMC活力高。因為種齡5的種子種齡較大,生長力較弱,加上易利用的營養成分少,所以CMC活力也較低。
從表1還可以看出,無論是單菌種還是混合菌種,種齡1和種齡2接種量2%的CMC活力均低于接種量1%的CMC活力;種齡3、種齡4和種齡5接種量2%的CMC活力均高于接種量1%的CMC活力。這說明接種量對CMC活力的影響還受到種齡的影響,在確定最佳接種條件時,只利用單因素試驗得到的結果是不準確的。產生這種結果的原因是當種齡小、接種量大時,初期菌種密度大,初級代謝產物多,菌體易衰老;而接種量較少則不利于菌體快速增殖和產酶[24]。接種量較小會延長菌種在發酵罐中生長速度,降低生產效率;而接種量較大會降低培養基中溶解氧的含量,影響好氧菌的生長和產物的合成[25]。進一步對相同菌種第8天的CMC活力方差分析和多重比較表明,在接種單菌種時,種齡3和種齡5不同接種量的CMC活力差異顯著,其他種齡的CMC活力差異不顯著;在接種混合菌種時,種齡3、種齡4和種齡5不同接種量的CMC活力差異顯著,其他2個種齡的CMC活力差異不顯著。余夢頔等[26]研究發現,接種量對納豆枯草桿菌產納豆激酶無顯著影響。因此,接種量與種齡共同對發酵產物的產量產生影響。
3 結論
在第8天時,相同菌種和接種量的不同種齡的纖維素酶活力有一定差異。種齡和接種量對微生物產纖維素酶具有協同作用;種齡不同,不同的接種量對纖維素酶活性影響不同。采用該研究的菌種制備纖維素酶時,無論單菌種還是混合菌種,最優接種條件都是種齡4、接種量2%,可以縮短發酵期的培養時間,達到節能降耗的效果。
參考文獻
[1] 帥正彬,陳璐,姚雅韋,等.秸稈原料綜合利用的研究進展[J].農業科技通訊,2015(12):186-192.
[2]? 王寶山,周景宇.對農作物秸稈綜合利用發展方向的探索[J].農業機械,2009(18):75-76.
[3] 佟毅.纖維素乙醇產業發展現狀及展望[J].中國糧食經濟,2019(12):47-50.
[4] 王文丹,唐新仁,吳天佑,等.纖維素酶在奶牛生產上的應用[J].畜牧與飼料科學,2017,38(12):34-37.
[5]? 劉晶,李晨曦,劉洪斌.纖維素酶在造紙過程中的應用[J].中國造紙,2018,37(2):48-53.
[6]? 石麗華,曾方明,李勉.纖維素酶在生物質轉化中的應用研究進展[J].生物產業技術,2019(3):69-76.
[7] 楊騰騰,周宏,王霞,等.微生物降解纖維素的新機制[J].微生物學通報,2015,42(5):928-935.
[8] 趙萍,夏文旭,郭健,等.一株玉米秸稈纖維素分解菌株的分離鑒定及酶學性質[J].微生物學通報,2016,43(5):991-997.
[9] 黃春凱,左小明,王紅蕾,等.一株產纖維素酶菌株的分離、鑒定及產酶特性[J].微生物學通報,2015,42(4):646-653.
[10] 穆春雷,武曉森,李術娜,等.低溫產纖維素酶菌株的篩選、鑒定及纖維素酶學性質[J].微生物學通報,2013,40(7):1193-1201.
[11] 孟建宇,李蘅,樊兆陽,等.低溫纖維素降解菌的分離與鑒定[J].應用與環境生物學報,2014,20(1):152-156.
[12] 鄒榮松,李素艷,孟童瑤,等.一株分離自綠化廢棄物堆肥枯草芽孢桿菌的常壓室溫等離子誘變及其發酵條件[J].微生物學通報,2019,46(11):2919-2926.
[13] KRISHNA S,MASLOV S,SNEPPEN K.UV-induced mutagenesis in Escherichia coli SOS response:A quantitative model[J].PLoS computational biology,2007,3(3):1-12.
[14] YOON S H,LEE E G,DAS A,et al.Enhanced vanillin production from recombinant E.coli using NTG mutagenesis and adsorbent resin[J].Biotechnology progress,2007,23(5):1143-1148.
[15] 于欣水,曾鈺,李潔,等.MHF組蛋白折疊復合體組分編碼基因MHF1過表達對重組釀酒酵母纖維素酶生產的影響[J].微生物學通報,2019,46(1):75-83.
[16] 張珂珂,周嬌嬌,佘煒怡,等.敲除組蛋白去乙酰化酶基因hdac對里氏木霉纖維素酶表達的調控作用[J].微生物學通報,2020,47(1):88-96.
[17] 田文,金美玉,于垚,等.生物反應器中金線蓮叢生芽的增殖及酚和黃酮積累的研究[J].延邊大學農學學報,2018,40(2):8-13,23.
[18] CIMINI D,ROSA M D,SCHIRALDI C.Production of glucuronic acid-based polysaccharides by microbial fermentation for biomedical applications[J].Biotechnology journal,2012,7(2):237-250.
[19] HEIDARI M,KARIMINIA H R,SHAYEGAN J.Effect of culture age and initial inoculum size on lipid accumulation and productivity in a hybrid cultivation system of Chlorella vulgaris[J].Process safety & environmental protection,2016,104:111-122.
[20] 王曉芳,徐旭士,吳敏,等.一株纖維素分解菌的分離與篩選[J].生物技術,2001,11(2):27-30.
[21] 韓美玲,邊祿森,姜宏浩,等.不同碳氮源對糙皮側耳木質纖維素酶活性的影響[J].菌物學報,2020,39(8):1538-1550.
[22] 孫寶婷,祝文興,劉新利.產埃博霉素B纖維堆囊菌發酵條件的優化[J].齊魯工業大學學報(自然科學版),2017,31(2):35-40.
[23] 劉元望,李兆君,馮瑤,等.微生物降解抗生素的研究進展[J].農業環境科學學報,2016,35(2):212-224.
[24] 師璐,李陽,李國龍,等.高產纖維素酶枯草芽孢桿菌S-16的篩選及其發酵工藝優化[J].生物資源, 2017,39(2):118-124.
[25] 毛婷,魏亞琴,楊紅建,等.牦牛糞便中纖維素降解菌的篩選及產酶優化[J].中國農業大學學報,2019,24(11):106-116.
[26] 余夢頔,余瓊.納豆枯草桿菌產酶條件的優化[J].黑龍江科學,2019,10(12):38-39.
基金項目 中央高校基本科研業務費專項資金項目(2572014BA16)。
作者簡介 賈洪柏(1978—),男,遼寧遼陽人,實驗師,博士,從事環境微生物學和微生物遺傳學研究。
收稿日期 2022-05-03
