鐵皮石斛SPL膜結(jié)合(STM)轉(zhuǎn)錄因子的全基因組鑒定及表達(dá)分析
2023-07-17 06:24:36楊樂聶聰龍小琴何基澤顏超越朱乾坤王萬軍
廣西植物 2023年6期
關(guān)鍵詞:功能分析
楊樂 聶聰 龍小琴 何基澤 顏超越 朱乾坤 王萬軍
摘 要:? SPL轉(zhuǎn)錄因子廣泛參與植物生長發(fā)育、脅迫響應(yīng)等過程。目前,沒有關(guān)于鐵皮石斛SPL膜結(jié)合(SPL with transmembrane motif)轉(zhuǎn)錄因子即STM轉(zhuǎn)錄因子的研究。為了探究STM轉(zhuǎn)錄因子在鐵皮石斛生長發(fā)育及脅迫響應(yīng)等方面的作用,該文在鐵皮石斛全基因組水平鑒定出4個STM轉(zhuǎn)錄因子,并對DoSTM基因家族成員進行生物信息學(xué)分析,又利用逆轉(zhuǎn)錄PCR研究了DoSTM在不同組織部位及不同逆境處理下的表達(dá)情況。結(jié)果表明:(1)DoSTM1-4為親水蛋白,均具有SBP保守結(jié)構(gòu)域和一些激素響應(yīng)位點。(2)4個DoSTM在根莖葉中均有表達(dá),DoSTM2在葉中的相對表達(dá)量最低;DoSTM1/3/4的相對表達(dá)水平均無明顯差異。(3)DoSTM1-4在低溫、高溫、干旱脅迫下的相對表達(dá)水平都有顯著變化,DoSTM1/3/4的表達(dá)量降低最為明顯,故推測DoSTM與植物體內(nèi)激素響應(yīng)、溫度變化響應(yīng)及抗旱性有關(guān)。這些結(jié)論為后續(xù)進一步開展鐵皮石斛STM轉(zhuǎn)錄因子的研究提供了參考。
關(guān)鍵詞: 鐵皮石斛, STM轉(zhuǎn)錄因子, 基因鑒定, 功能分析, 基因表達(dá)
中圖分類號:? Q943 文獻標(biāo)識碼:? A 文章編號:? 1000-3142(2023)06-1041-10
Genome-wide identification and expression analysis of SPL with transmembrane motif (STM) transcription factor in Dendrobium officinale
YANG Le, NIE Cong, LONG Xiaoqin, HE Jize, YAN Chaoyue, ZHU Qiankun, WANG Wanjun*
( School of Life Science and Engineering, Southwest Jiaotong University, Chengdu 610031, China )
Abstract:? SPL transcription factors are widely involved in plant growth and development, stress response and so on. At present, there is no study on the? STM (SPL with transmembrane motif) transcription factor in Dendrobium officinale. In order to explore the role of STM in the growth, development and stress response of D. officinale, four STM transcription factors were identified at the whole genome of D. officinale, and bioinformatics analysis of DoSTM gene family members were carried out. The expression of DoSTM in different tissue parts and different stress treatments were studied by reverse transcription PCR. The results were as follows: (1)DoSTM1-4 were hydrophilic proteins with SBP conserved domains and some hormone response sites. (2)Four DoSTM were expressed in root, stem and leaf, and the relative expression of DoSTM2 was the lowest in leaf; there was no significant differences in the relative expression level of DoSTM1/3/4. (3)The relative expression level of DoSTM1-4 changed significantly under low temperature, high temperature and drought stress, and the expression of DoSTM1/3/4 decreased most significantly. Therefore, it is speculated that DoSTM is related to hormone response, temperature change response and drought resistance in plants. These conclusions provide the reference for the further research on STM transcription factor of D. officinale.
Key words: Dendrobium officinale, STM transcription factor, gene identification, functional analysis, gene expression
石斛主要分布在東南亞和大洋洲的熱帶及亞熱帶地區(qū),喜生長在溫暖潮濕的環(huán)境中,附著在樹干或巖石上 (羅凱等,2021)。鐵皮石斛(Dendrobium officinale)為蘭科石斛屬多年生草本植物,具有極高藥用價值,是一種藥食兩用中藥材,其成分主要包括多糖類、生物堿類、黃酮類以及酚酸類等,在抗腫瘤和增強免疫力等方面具有功效 (唐文文等,2021)。近年來由于過度挖采導(dǎo)致野生資源大量減少,故從分子水平研究鐵皮石斛的生長發(fā)育對其保育有重要意義 (曾丹琦等,2021)。
SPL(squamousa promoter binding protein like)基因家族作為植物特有的一類轉(zhuǎn)錄因子,主要通過結(jié)合下游基因啟動子區(qū)的順式作用元件來調(diào)控下游基因表達(dá)。SPL轉(zhuǎn)錄因子在植物的生長發(fā)育、信號傳導(dǎo)、脅迫響應(yīng)等方面有著重要的作用 (吳艷等,2019)。這類轉(zhuǎn)錄因子最早在金魚草中被發(fā)現(xiàn)(Huijser et al., 1992)。之后有更多研究發(fā)現(xiàn)SPL轉(zhuǎn)錄因子在植物生長發(fā)育以及在脅迫響應(yīng)中起著重要作用 (Yu et al., 2015; Xu et al., 2016)。在甘藍(lán)型油菜的愈傷組織、根、莖、葉、芽、花和角果中有大量BoSPL表達(dá),并且這些基因可能在甘藍(lán)耐寒性中起重要作用 (Shan et al., 2021)。據(jù)最近報道,擬南芥SPLs可能參與其從幼年到成年的營養(yǎng)轉(zhuǎn)化、生殖期的形態(tài)變化、花青素生物合成和防御脅迫 (Jiang et al., 2021)。其中,SPL3在花序和花器官的發(fā)育中發(fā)揮重要作用 (Gandikota et al., 2007),在各種低磷脅迫反應(yīng)以及低磷條件下磷饑餓誘導(dǎo)基因表達(dá)中都有SPL1的參與介導(dǎo) (雷凱健等,2016)。
轉(zhuǎn)錄因子的研究是植物分子生物學(xué)的重要內(nèi)容,在植物抗逆等方面起著重要作用。轉(zhuǎn)錄因子和基因順式作用元件的相互作用激活了相關(guān)抗逆基因的表達(dá),提高了植物的抗逆性。有一類特殊的轉(zhuǎn)錄因子存在,因其含有一段跨膜區(qū)被稱為膜結(jié)合轉(zhuǎn)錄因子,可直接整合在細(xì)胞內(nèi)的膜結(jié)構(gòu)上 (如細(xì)胞質(zhì)膜、內(nèi)質(zhì)網(wǎng)膜、核膜等),一般處于休眠狀態(tài),當(dāng)受到外界環(huán)境變化刺激后,膜結(jié)合轉(zhuǎn)錄因子便從膜上釋放,轉(zhuǎn)變?yōu)榧せ顮顟B(tài),并轉(zhuǎn)運到細(xì)胞核內(nèi)行使功能 (王楠等,2016)。SPL轉(zhuǎn)錄因子中也有這類膜結(jié)合轉(zhuǎn)錄因子,且目前并沒有專門研究SPL膜結(jié)合轉(zhuǎn)錄因子的報道,因此,本研究通過BLASTP在鐵皮石斛全基因組中鑒定出4個SPL膜結(jié)合轉(zhuǎn)錄因子,命名為DoSTM (SPL with transmembrane motif)轉(zhuǎn)錄因子,并進行后續(xù)生物信息學(xué)及表達(dá)分析。
1 材料與方法
1.1 材料
將鐵皮石斛幼苗分別置于1/2MS培養(yǎng)基、含100 mmol·L-1 NaCl的1/2MS液體培養(yǎng)基、含5 μmol·L-1脫落酸的1/2MS液體培養(yǎng)基、質(zhì)量分?jǐn)?shù)為10% PEG的1/2MS液體培養(yǎng)基、40 ℃ 1/2MS液體培養(yǎng)基、4 ℃ 1/2MS液體培養(yǎng)基中處理6 h,吸干水分后將幼苗按200~300 mg裝入1.5 mL離心管中,并做好標(biāo)記,迅速置于液氮中1 min,取出放入-80 ℃冰箱保存待用。以盆栽鐵皮石斛幼嫩的根莖葉組織作為原材料,進行根莖葉差異表達(dá)分析。
1.2 試劑
RNAprep Pure多糖多酚植物總RNA提取試劑盒(北京擎科)、瓊脂糖(北京擎科)、2×TSINGKE Master Mix(北京擎科)、RNA無酶水(北京擎科)、PrimeScriptTM II 1st Strand cDNASynthesis Kit試劑盒(寶生物)、50×TAE(上海生工)。
1.3 方法
1.3.1 鐵皮石斛STM蛋白的理化性質(zhì)及亞細(xì)胞定位分析 從NCBI數(shù)據(jù)庫中下載擬南芥STM蛋白序列,將物種限定在鐵皮石斛(Dendrobium officinale)并進行BLAST搜索,下載搜索到的鐵皮石斛的STM蛋白序列,去除冗余序列,利用TMHMM(http://www.cbs.dtu.dk/services/TMHMM-2.0/)預(yù)測跨膜結(jié)構(gòu)域,最終選取XP_020685848、XP_020681923、XP_020672795、XP_020688542這4個蛋白進行研究,并分別命名為DoSTM1、DoSTM2、DoSTM3、DoSTM4。利用ProtParam(https://web.expasy.org/protparam/)分析鐵皮石斛STM蛋白家族的理化性質(zhì)。用Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant/) (Chou & Shen, 2007)對DoSTM蛋白家族進行亞細(xì)胞定位分析。
1.3.2 鐵皮石斛STM家族的系統(tǒng)進化樹分析 從NCBI數(shù)據(jù)庫中下載野蕉 (Musa balbisiana)、鐵皮石斛 (Dendrobium officinale)、擬南芥 (Arabidopsis thaliana)、蝴蝶蘭 (Phalaenopsis equestris)、毛白楊 (Populus tomentosa)、扇形文心蘭 (Erycina pusilla)、玉米 (Zea mays)、水稻 (Oryza sativa)、生姜 (Zingiber officinale)的STM蛋白序列Fasta格式。用MEGA-X鄰近法構(gòu)建進化樹,序列比對采用ClustalW,選擇Bootstrap method,Bootstrap replications選擇500,Model選擇p-distance,Gaps/Missing Data Treatment選擇Partial deletion,Site Coverage Cutoff設(shè)定為50,進行建樹。建樹結(jié)果在ITOL(https://itol.embl.de/)中進行美化。
1.3.3 鐵皮石斛STM蛋白質(zhì)二級結(jié)構(gòu)預(yù)測 利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)在線網(wǎng)站對鐵皮石斛STM蛋白質(zhì)的二級結(jié)構(gòu)進行分析,參數(shù)為默認(rèn)值。
1.3.4 鐵皮石斛STM蛋白質(zhì)序列比對及保守基序分析 將4個DoSTM蛋白導(dǎo)入MEME (Bailey & Elkan et al., 1994)在線網(wǎng)站(https://meme-suite.org/meme/tools/meme)進行分析,將motifs的數(shù)量設(shè)定在10,其他參數(shù)默認(rèn),進行分析。將鐵皮石斛STM蛋白序列Fasta格式導(dǎo)入MEGA-X中進行構(gòu)樹,序列比對采用ClustalW,選擇Bootstrap method,Bootstrap replications選擇500,Model選擇p-distance,Gaps/Missing Data Treatment選擇Partial deletion,Site Coverage Cutoff設(shè)定為50,進行構(gòu)樹。從NCBI數(shù)據(jù)庫中下載鐵皮石斛STM蛋白的CDD,利用TBtools (Chen et al., 2020)對DoSTM保守基序、保守結(jié)構(gòu)域、進化樹進行繪圖。將鐵皮石斛STM蛋白的ClustalW比對結(jié)果導(dǎo)入ESPript 3.0在線網(wǎng)站(https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi)進行分析。
1.3.5 鐵皮石斛STM基因結(jié)構(gòu)及順式作用元件分析 從NCBI數(shù)據(jù)庫中下載鐵皮石斛的基因組信息,用TBtools進行基因結(jié)構(gòu)繪制,同時獲得啟動子上游2 000 bp的序列信息,獲得的序列信息用PlantCARE (Lescot et al., 2002)網(wǎng)站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)進行順式作用元件預(yù)測,并用TBtools進行繪圖。
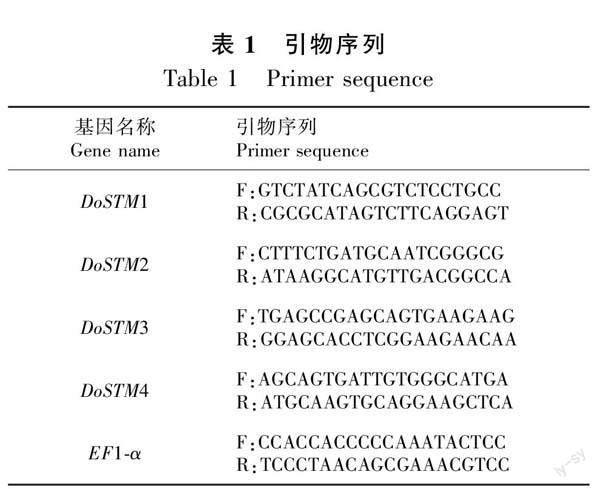
1.3.6 引物設(shè)計 利用4個DoSTM的cDNA序列,將所得的cDNA序列用Primer-BLAST進行引物設(shè)計,引物大小設(shè)定為18~22 bp,PCR產(chǎn)物大小設(shè)定為400~800 bp,其他參數(shù)均為默認(rèn)參數(shù),在輸出的引物中選擇靠近3′端的作為試驗所用引物,選用EF1-α做內(nèi)參,相應(yīng)的引物序列如表1所示。
1.3.7 鐵皮石斛STM基因表達(dá)分析 將鐵皮石斛幼苗分別置于1/2MS液體培養(yǎng)基、含100 mmol·L-1 NaCl的1/2MS液體培養(yǎng)基、含5 μmol·L-1 脫落酸的1/2MS液體培養(yǎng)基、含10%PEG的1/2MS液體培養(yǎng)基、40 ℃ 1/2MS液體培養(yǎng)基、4 ℃ 1/2MS液體培養(yǎng)基中處理6 h,迅速置于液氮中1 min,取出放入-80 ℃冰箱保存待用。首先,用RNAprep Pure多糖多酚植物總RNA提取試劑盒提取6種處理的鐵皮石斛幼苗的RNA和鐵皮石斛幼苗根莖葉的RNA,并用PrimeScriptTM Ⅱ 1st Strand cDNA Synthesis Kit進行cDNA第一鏈的合成。然后,以cDNA第一鏈為模板,2×TSINGKE Master Mix及引物進行PCR擴增。PCR反應(yīng)程序:98 ℃預(yù)變性2 min;98 ℃變性15 s,55 ℃退火15s,72 ℃延伸1 min,變性-退火-延伸30個循環(huán);72 ℃繼續(xù)延伸10 min。最后,進行1%凝膠電泳,拍照記錄電泳條帶。
1.3.8 數(shù)據(jù)處理及分析 用ImageJ測定電泳條帶的灰度值,Excel對所得灰度值進行處理,將處理后的數(shù)據(jù)導(dǎo)入GraphPad Prism 5中進行分析及作圖,分析方法選用Students t-test,顯著水平均設(shè)為0.05。
2 結(jié)果與分析
2.1 鐵皮石斛STM蛋白家族理化性質(zhì)及亞細(xì)胞定位分析
DoSTM蛋白家族的理化性質(zhì)分析結(jié)果表明,DoSTM1-4的氨基酸數(shù)目分別為1 025、977、1 104、830,相對分子質(zhì)量分別為113.53、109.09、121.67、93.177;等電點pI在6.07~8.06之間;不穩(wěn)定系數(shù)在44.37~58.41之間,較為不穩(wěn)定;脂肪系數(shù)在78.15~83.04之間;總平均親水系數(shù)均為負(fù)數(shù),在-0.405~-0.335之間,表明DoSTM蛋白均為親水蛋白質(zhì)(表2)。亞細(xì)胞定位預(yù)測結(jié)果顯示DoSTM1-3定位在細(xì)胞核,DoSTM4定位在葉綠體。將篩選到的鐵皮石斛STM蛋白DoSTM1-4進行跨膜結(jié)構(gòu)域預(yù)測 (圖1),結(jié)果顯示跨膜結(jié)構(gòu)域位于C末端22個氨基酸殘基處。
2.2 鐵皮石斛STM基因家族的系統(tǒng)進化分析
系統(tǒng)進化分析顯示9個物種的34個STM蛋白聚為4組 (A-D)(圖2),A組的STM蛋白共有3個,B組的STM蛋白共有6個,C組的STM蛋白共有9個,D組的STM蛋白共有16個。其中,DoSTM1-2蛋白共同聚類在B組,有可能是較近發(fā)生的基因復(fù)制事件所造成;DoSTM4聚類在C組;DoSTM3蛋白聚類在D組。
2.3 鐵皮石斛STM蛋白的二級結(jié)構(gòu)
對DoSTM蛋白質(zhì)的二級結(jié)構(gòu)進行分析,二級結(jié)構(gòu)占比如表3所示,二級結(jié)構(gòu)圖如圖3所示。結(jié)果表明,鐵皮石斛STM蛋白的二級結(jié)構(gòu)包括α螺旋、伸展鏈、β轉(zhuǎn)角和無規(guī)卷曲,其中,鐵皮石斛STM蛋白的α螺旋占比為28.62%~33.76%,伸展鏈占比為11.68%~14.70%,β轉(zhuǎn)角占比為4.09%~4.78%,無規(guī)卷曲占比為49.56%~55.34%。
2.4 鐵皮石斛STM家族蛋白結(jié)構(gòu)分析
對4個DoSTM蛋白進行多序列比對 (圖4),結(jié)果顯示DoSTM1-2蛋白序列同源性較高。DoSTM蛋白的保守基序分析結(jié)果表明4個DoSTM蛋白均有SBP保守結(jié)構(gòu)域,而DoSTM1/3還同時包含Ank_2 superfamily,DoSTM2則還包含ANKYR (圖5)。同源的DoSTM1與DoSTM2含有motif 1-10,而DoSTM3與其相比缺少motif 7/10,DoSTM4不僅缺少motif 7/10,還缺少motif 3/4 (圖5)。表明4個DoSTM的蛋白結(jié)構(gòu)并不高度保守。
2.5 鐵皮石斛STM基因結(jié)構(gòu)及順式作用元件分析
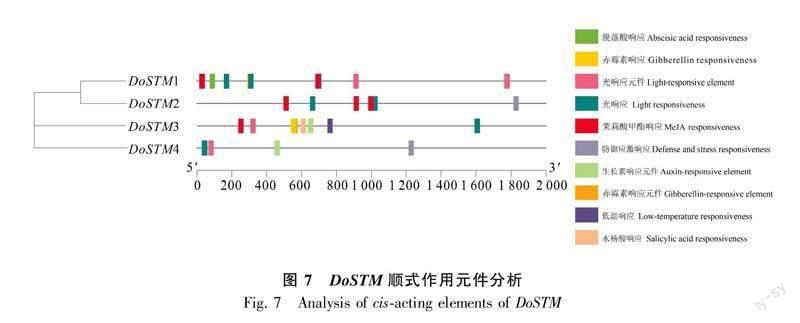
DoSTM基因結(jié)構(gòu)分析如圖6所示,發(fā)現(xiàn)4個DoSTM均有10個編碼區(qū)(由于gff3未注釋UTR,故未標(biāo)識非編碼區(qū))。順式作用元件分析發(fā)現(xiàn)4個DoSTM中均有光響應(yīng)區(qū)和光響應(yīng)元件,其中DoSTM1-3共同包含茉莉酸甲酯響應(yīng)區(qū),而防御應(yīng)急響應(yīng)只在DoSTM2/4中具備,除此之外DoSTM4還含有生長素響應(yīng)元件(圖7)。而DoSTM1則可以對脫落酸作出響應(yīng),DoSTM3作為包含順式作用元件最多的STM蛋白,它還可以對赤霉素、水楊酸、生長素及低溫作出響應(yīng)。
2.6 鐵皮石斛STM的表達(dá)分析
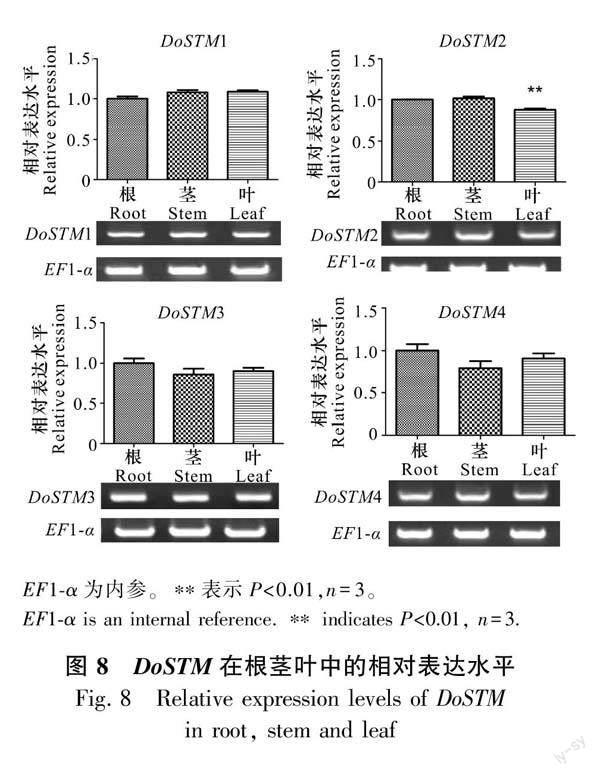
2.6.1 鐵皮石斛STM在根莖葉中的表達(dá)情況
我們對DoSTM在根莖葉3個不同器官的相對表達(dá)水平進行了分析 (圖8)。本研究發(fā)現(xiàn),DoSTM在根莖葉中均有表達(dá),DoSTM2在葉中的相對表達(dá)量顯著降低,而在莖中的表達(dá)水平則無明顯差異,DoSTM1/3/4的相對表達(dá)水平均無明顯差異,DoSTM1在莖和葉中的表達(dá)量相差無幾,但略高于根,而DoSTM3-4則在葉中的相對表達(dá)水平高于莖,且都低于根。
2.6.2 鐵皮石斛STM在脅迫處理下的表達(dá)情況 鐵皮石斛多生長在懸崖峭壁,會受到多種非生物脅迫,STM轉(zhuǎn)錄因子也有被報道參與多種抗逆反應(yīng) (王楠等,2016),故本研究進一步分析了DoSTM在不同脅迫條件下的表達(dá)情況 (圖9)。結(jié)果表明,DoSTM1在4 ℃低溫、40 ℃高溫和10% PEG脅迫處理下的相對表達(dá)水平低于對照組,而在ABA脅迫處理下的相對表達(dá)水平無顯著差異,但NaCl脅迫處理下,表達(dá)量略有降低;DoSTM2在4 ℃低溫和40 ℃高溫脅迫處理下的相對表達(dá)水平與對照組相比顯著降低,在脫落酸、NaCl、10%PEG脅迫處理下無明顯差異;DoSTM3在NaCl、4 ℃低溫和40 ℃高溫脅迫處理下的相對表達(dá)水平與對照組相比顯著降低,在脫落酸和10%PEG脅迫處理下與對照組相比稍有差異;與DoSTM3相似的是,DoSTM4在4 ℃低溫和40 ℃高溫脅迫處理下有顯著變化,在脫落酸脅迫處理下也發(fā)生顯著變化,但在NaCl脅迫處理下并無明顯差異。
3 討論與結(jié)論
本研究從鐵皮石斛基因組上鑒定出4個STM蛋白序列,對4個STM編碼的蛋白進行生物信息學(xué)分析。結(jié)果表明,4個DoSTM蛋白均為親水蛋白質(zhì);DoSTM1/3/4的等電點均小于7,DoSTM2則大于7,這一點與大部分STM蛋白家族成員的理論等電點大于7有出入 (張曉紅等,2016;劉闖等,2017;祁香寧等,2018)。SPL基因家族蛋白含有1個高度保守的SBP-BOX結(jié)構(gòu)域,它是SPL蛋白和DNA分子特異性結(jié)合所必需的。這與我們在保守結(jié)構(gòu)域預(yù)測時的發(fā)現(xiàn)一致。
李豆等 (2021)將白樺BpSPL6基因啟動子驅(qū)動GUS基因在轉(zhuǎn)基因擬南芥營養(yǎng)生長期的根尖及根的其他部位表達(dá),發(fā)現(xiàn)在營養(yǎng)生長時期其在根部的表達(dá)也隨植物的生長逐漸增加,與前人研究一致,故他推測BpSPL6基因可能在植物的根發(fā)育過程中起作用。這與我們測得DoSTM2-4在根中的相對表達(dá)水平略高一致,推測DoSTM可能參與根的發(fā)育。
激素參與了植物生長發(fā)育的各個方面,已有多項研究表明SPL基因參與植物的激素響應(yīng)過程。對白樺BpSPL6基因啟動子的順式作用元件分析表明 (李豆等,2021),其啟動子區(qū)含有10種激素響應(yīng)元件 (生長素、赤霉素、水楊酸、脫落酸等)。這與我們得到的DoSTM順式作用元件分析結(jié)果類似,并且與在脫落酸脅迫條件下DoSTM3-4的相對表達(dá)水平發(fā)生顯著變化一致。利用RNA-seq研究發(fā)現(xiàn)擬南芥SPL10對茉莉酸、水楊酸和生長素等激素響應(yīng)過程有廣泛影響。AtSPL10通過調(diào)節(jié)生長素的生物合成抑制根的再生 (Ye et al., 2020)。赤霉素通過調(diào)節(jié)莖分生組織中的AtSPL3-5基因促進開花 (Galvo et al., 2012)。在DoSTM啟動子上也發(fā)現(xiàn)了多種激素響應(yīng)元件,故推測DoSTM可能參與多種激素響應(yīng)。
大量研究表明SPL基因能夠響應(yīng)低溫、高溫等非生物脅迫的信號。如Stief等 (2014)發(fā)現(xiàn)在高溫條件下,SPL2/9/11基因被miR156-f和miR156-h調(diào)控使其表達(dá)降低,進而延長植物對高溫脅迫的耐受性。除高溫條件外,葡萄對低溫條件也較敏感,在低溫條件下 (5 ℃),體內(nèi)的VvSBP3和VvSBP5表達(dá)均被上調(diào),而VvSBP4和VvSBP7表達(dá)明顯被下調(diào),表明VvSBP3和VvSBP5參與葡萄的低溫脅迫反應(yīng) (吳艷等,2019)。這與DoSTM1-4在低溫、高溫脅迫條件下的相對表達(dá)量發(fā)生顯著變化一致。
對白樺BpSPL6轉(zhuǎn)基因擬南芥進行了NaCl和PEG脅迫實驗,發(fā)現(xiàn)受到脅迫后,其表達(dá)量有下降(李豆等,2021),與我們觀測到的在PEG脅迫條件下,DoSTM的相對表達(dá)水平顯著下調(diào)一致,推測DoSTM很可能參與植物的干旱脅迫響應(yīng)。崔揚等 (2019)發(fā)現(xiàn)玉米遭受干旱脅迫時,ZmSPL16在根中的表達(dá)量顯著上調(diào),這與DoSTM4結(jié)果一致,但與DoSTM1/3不一致,具體原因有待進一步研究。
本研究在鐵皮石斛全基因組水平鑒定出4個STM,生物信息學(xué)分析顯示他們均為親水蛋白,大部分定位在細(xì)胞核,都具有高度保守的SBP結(jié)構(gòu)域以及與激素響應(yīng)有關(guān)的順式作用元件。表達(dá)分析顯示他們在根中的表達(dá)量略高于莖和葉,脅迫分析顯示其在低溫、高溫及干旱脅迫下相對表達(dá)水平顯著變化,且部分DoSTM可以對脫落酸作出響應(yīng)。以上研究結(jié)果為進一步研究DoSTM轉(zhuǎn)錄因子的生物學(xué)功能奠定了基礎(chǔ)。
參考文獻:
BAILEY TL, ELKAN C, 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers [J]. Proc Int Conf Intell Syst Mol Biol: 28-36.
CHOU KC, SHEN HB, 2007. Large-scale plant protein subcellular location prediction [J]. J Cell Biochem, 100(3): 665-78.
CHEN CG, CHEN H, ZHANG Y, et al., 2020. TBtools: An integrative toolkit developed for interactive analyses of big biological data [J]. Mol Plant, 13(8): 1194-1202.
CUI Y, FENG YH, CHEN ZF, et al., 2019. Cloning and functional identification of maize transcription factor ZmSPL16 [J]. Mol Plant Breed, 17(20): 6583-6589.? [崔揚, 馮彥輝, 陳眾峰, 等, 2019. 玉米轉(zhuǎn)錄因子ZmSTM16的克隆與功能鑒定 [J]. 分子植物育種, 17(20): 6583-6589.]
GALVO VC, HORRER D, KTTNER F, et al., 2012. Spatial control of flowering by DELLA proteins in Arabidopsis thaliana [J]. Development, 139(21): 4072-4082.
GANDIKOTA M, BIRKENBIHL RP, HOHMANN S, et al., 2007. The miRNA156/157 recognition element in the 3′UTR of the Arabidopsis SBP box gene SPL3 prevents early flowering by translational inhibition in seedlings [J]. Plant J, 49(4): 683-693.
HUIJSER P, KLEIN J, LNNIG WE, et al., 1992. Bracteomania, an inflorescence anomaly, is caused by the loss of function of the MADS-box gene squamosa in Antirrhinum majus \[J\]. EMBO J, 11(4): 1239-1249.
JIANG XW, CHEN P, ZHANG XW, et al., 2021. Comparative analysis of the SPL gene family in five Rosaceae species: Fragaria vesca, Malus domestica, Prunus persica, Rubusoccidentalis, and Pyrus pyrifolia [J]. Open Life Sci, 16(1): 160-171.
LUO K, LI ZS, BAI YB, et al., 2021. Current situation of diversity utilization and protection of Dendrobium [J]. Heilongjiang Agric Sci, (8): 85-89. [羅凱, 李澤生, 白燕冰, 等, 2021. 石斛蘭多樣性利用及保護現(xiàn)狀 [J]. 黑龍江農(nóng)業(yè)科學(xué), (8): 85-89.]
LEI KJ, REN J, ZHU YY, et al., 2016. Arabidopsis SPL1 gene is involved in regulating rhizosphere acidification under low phosphorus conditions [J]. Acta Bot Sin, 51(2): 184-193. [雷凱健, 任晶, 朱園園, 等, 2016. 擬南芥SPL1基因參與調(diào)節(jié)低磷條件下的根際酸化反應(yīng) [J]. 植物學(xué)報, 51(2): 184-193.]
LESCOT M, DEHAIS P, THIJS G, et al., 2002. Plant CARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences [J]. Nucl Acids Res, 30(1): 325-7.
LI D, SU GB, HU XQ, et al., 2021. Cloning and expression analysis of BpSPL6 gene promoter of Betula platyphylla [J]. J Beijing For Univ. https://kns.cnki.net/kcms/detail/ 11.1932.S.20210610. 0948.001.html. [李豆, 蘇功博, 胡曉晴, 等, 2021. 白樺BpSPL6基因啟動子的克隆及表達(dá)分析 [J]. 北京林業(yè)大學(xué)學(xué)報. https://kns.cnki.net/kcms/detail/ 11.1932.S.20210610.0948.001. html.]
LIU C, 2017. Identification of 18 Betula platyphylla STMs genes and functional analysis of BpSPL8 gene [D]. Harbin: Northeast Forestry University. [劉闖, 2017. 18個白樺STMs基因的鑒定及BpSPL8基因的功能分析 [D]. 哈爾濱: 東北林業(yè)大學(xué).]
QI XN, 2018. Identification, evolution and expression analysis of Actinidia sinensis SBP-box transcription factor gene [D]. Yangling: Northwest A & F University. [祁香寧, 2018. 獼猴桃SBP-box轉(zhuǎn)錄因子基因的鑒定、進化及其表達(dá)分析 [D]. 楊凌: 西北農(nóng)林科技大學(xué).]
SHAN X, ZHANG W, HUANG JX, et al., 2021. Identification and characterization of SPL transcription factor family reveals organization and chilling-responsive patterns in cabbage (Brassica oleracea var. capitata L.) [J]. Agronomy, 11(7): 1445-1445.
STIEF A, ALTMANN S, HOFFMANN K, et al., 2014. Arabidopsis miR156 regulates tolerance to recurring environmental stress through SPL transcription factors [J]. Plant Cell, 26(4): 1792-1807.
TANG WW, XIA JL, CHEN Y, 2021. Functional components, antioxidant activity and correlation of stem, leaf and flower of Dendrobium officinale [J]. Food Mach, 37(7): 45-50. [唐文文, 夏俊麗, 陳垣, 2021. 鐵皮石斛莖、葉、花功能性成分、抗氧化活性及其相關(guān)性 [J]. 食品與機械, 37(7): 45-50.]
WANG N, XIANG FN, LI S, 2016. Advance in plant membrane-bound transcription factors and stress response [J]. Chin Bull Life Sci, 28(7): 799-806. [王楠, 向鳳寧, 李朔, 2016. 植物膜結(jié)合轉(zhuǎn)錄因子與脅迫響應(yīng) [J]. 生命科學(xué), 28(7): 799-806.]
WU Y, HOU ZH, CHENG Q, et al., 2019. Research progress of SPL transcription factors [J]. Soybean Sci, 38(2): 304-310. [吳艷, 侯智紅, 程群, 等, 2019. SPL轉(zhuǎn)錄因子的研究進展 [J]. 大豆科學(xué), 38(2): 304-310.]
XU ML, HU TQ, ZHAO JF, et al., 2016. Developmental functions of miR156-regulated SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) genes in Arabidopsis thaliana [J]. PLoS Genet, 12(8): e1006263.
YE BB, SHANG GD, PAN Y, et al., 2020. AP2/ERF transcription factors integrate age and wound signals for root regeneration [J]. Plant Cell, 32(1): 226-241.
YU ZX, WANG LJ, ZHAO B, et al., 2015. Progressive regulation of sesquiterpene biosynthesis in Arabidopsis and patchouli (Pogostemon cablin) by the miR156-targeted SPL transcription factors [J]. Mol Plant, 8(1): 98-110.
YANG ZT, XIONG ML, JIAN Y, et al., 2018. Research progress on the activation mechanism of membrane-bound transcription factors in endoplasmic reticulum stress [J]. Mol Plant Breed, 16(24): 8028-8033. [楊正婷, 熊孟連, 簡燕, 等, 2018. 內(nèi)質(zhì)網(wǎng)應(yīng)激中膜結(jié)合轉(zhuǎn)錄因子的活化機制研究進展 [J]. 分子植物育種, 16(24): 8028-8033.]
ZENG DQ, ZHANG MZ, HE CM, et al., 2021. Identification and analysis of WOX transcription factors in Dendrobium officinale [J]. J Trop Subtrop Plants, 29(3): 301-310. [曾丹琦, 張明澤, 何春梅, 等, 2021. 鐵皮石斛WOX轉(zhuǎn)錄因子的鑒定和分析 [J]. 熱帶亞熱帶植物學(xué)報, 29(3): 301-310.]
ZHANG XH, 2016. Functional study and regulatory analysis of flowering related genes in upland cotton [D]. Yangling: Northwest A & F University.? [張曉紅, 2016. 陸地棉開花相關(guān)基因的功能研究及調(diào)控分析 [D]. 楊凌: 西北農(nóng)林科技大學(xué).]
(責(zé)任編輯 周翠鳴)
收稿日期:? 2022-01-02
基金項目:? 國家自然科學(xué)基金(31900164); 中央高校基本科研業(yè)務(wù)費(2682021CX121); 西南交大個性化實驗項目(GX2021160027)。
第一作者: 楊樂(1997-),碩士,主要從事植物生長發(fā)育與次生代謝研究,(E-mail)2295402195@qq.com。
*通信作者:? 王萬軍,博士,教授,主要從事植物生長發(fā)育與次生代謝研究,(E-mail) wanjunwang@home.swjtu.edu.cn。
猜你喜歡
現(xiàn)代交際(2016年20期)2017-02-15 18:33:15
科技資訊(2016年19期)2016-11-15 10:39:12
電子技術(shù)與軟件工程(2016年18期)2016-11-14 00:49:04
戲劇之家(2016年20期)2016-11-09 23:08:07
老區(qū)建設(shè)(2016年4期)2016-09-28 05:09:17
電腦知識與技術(shù)(2016年16期)2016-07-22 19:10:51
科技視界(2016年13期)2016-06-13 21:00:36
戲劇之家(2016年6期)2016-04-16 11:52:45
科技視界(2016年4期)2016-02-22 20:53:20
科技資訊(2015年8期)2015-07-02 18:49:25
