三華李ANS 基因克隆及其在果實采后轉色過程中的表達分析
2023-08-11 10:17:40許薇陳振林劉英健勞琪珍宋慕波
山東農業科學 2023年7期
許薇,陳振林,劉英健,勞琪珍,宋慕波
(1.賀州學院食品科學與工程技術研究院,廣西 賀州 542899;2.大連工業大學食品學院,遼寧 大連 116034)
三華李(Prunus salicinaLindl.cv.Sanhua)屬于薔薇科李亞科,是華南地區著名特色水果,原產自廣東翁源縣,后逐漸擴展至南方各省份[1,2]。三華李果實為圓形或近圓形,果肉紫紅色,肉質爽脆,酸甜可口,營養價值高[3]。 三華李成熟果實中富含花青素,這是其重要特點[4];有研究表明三華李中總花青素含量僅次于黑布李,花青素主要成分為矢車菊素[5]。 目前針對三華李果實花青素合成的相關研究較少。
花青素是植物中廣泛存在的一種水溶性天然色素,屬于類黃酮化合物,在新鮮的蔬菜和水果中大量存在,使其呈現出明亮的顏色,從而吸引消費者;同時,花青素類物質也具有抗氧化、清除自由基等多種生理活性功能[6-9]。 花青素的合成主要分3 個階段:第一階段是苯丙氨酸經過多步反應生成4-香豆酰CoA;第二階段是4-香豆酰CoA在系列酶的催化作用下生成黃酮類物質(二氫槲皮素、二氫楊梅素)和黃酮醇;第三階段是二氫槲皮素、二氫楊梅素和黃酮醇這三類物質在二氫黃酮醇-4-還原酶(dihydroflavonol-4-reductase,DFR)的作用下生成無色的花青素(無色矢車菊素、無色天竺葵素和無色翠雀素),然后花青素合成酶(ANS)將無色的花青素轉化為相應的花青素類物質(矢車菊素、天竺葵素和翠雀素),再經過糖基化、酰基化、甲基化等過程的修飾作用,最終形成穩定可見的花青素[10]。 可見,ANS 是花青素生物合成途徑后期的關鍵酶,能催化無色花青素脫水氧化形成有色花青素。 編碼ANS 的基因首先在紫蘇中被克隆[11],在多數植物中,ANS由一個小基因家族所編碼。 已有研究發現,在參薯[12]、紫色不結球白菜[13]、蘋果梨[14]和鳳丹牡丹[15]中ANS基因的表達與組織中花青素含量呈正相關。 然而,草莓果實從轉紅到完全成熟的過程中,雖然花青素含量呈上升趨勢,但ANS基因表達量卻有所下降[16]。 葡萄果實經乙醇處理后花青素含量增加,但基因表達水平沒有變化[17]。 由此可見,花青素的生物合成存在復雜的調控機制。
三華李屬呼吸躍變型果實,后熟過程中花青素快速積累,但目前尚未有針對三華李花青素合成基因的系統研究。 雖然三月李的基因組測序工作已由國內課題組完成,為李屬果樹的分子生物學研究奠定了基礎[18],但仍未有針對三華李ANS基因的相關研究。 本研究從三華李果實中克隆獲得ANS基因,對其序列進行生物信息學分析,并研究其在三華李不同組織和果實后熟過程中的表達模式,為進一步探究三華李花青素合成途徑及其調控機理奠定基礎。
1 材料與方法
1.1 試驗材料
供試三華李植株種植于廣西賀州市賀街鎮三華李驛站果園。 2021 年6 月1 日(花后約130天)采集七八成熟的果實及莖、葉。 果實采后立即運回實驗室,挑選大小均一、無病害及機械損傷的果實,分為兩組,分別作為對照組和乙烯處理組,分別用自來水和5 mL/L 乙烯利溶液浸泡1 min,撈出晾干后自封袋密封24 h,之后打開自封口通風透氣,于25℃下貯藏;每兩天取樣一次,放入-80℃超低溫冰箱保存。 莖、葉及果皮和果肉各組織用液氮速凍,-80℃保存待用。
1.2 試驗方法
1.2.1 RNA 的提取與cDNA 合成 利用華越洋植物RNA 提取試劑盒提取三華李不同組織和不同后熟階段的總RNA,具體操作方法依照說明書進行。 得到總RNA 后通過超微量分光光度計測定其濃度。 以提取的總RNA 為模板,利用反轉錄試劑盒進行反轉錄合成cDNA,合成方法參照試劑盒說明書,保存于-20℃冰箱中,用于后續實驗。
1.2.2PsANS基因的克隆 從三華李轉錄組數據庫中獲得ANS 序列信息,利用Primer Premier 5.0軟件設計特異性引物,引物序列見表1。 以三華李不同后熟階段的cDNA 為模板,克隆PsANS基因的CDS 序列,進行PCR 擴增。 PCR 反應程序為:94℃預變性3 min;95℃變性15 s,55℃退火30 s,72℃延伸2 min,35 個循環;72℃延伸10 min,于4℃保存。 PCR 產物經過回收、連接并轉化至DH5α 感受態細胞,鑒定陽性克隆后測序。 引物合成和基因測序全部交由上海生物科技有限公司完成。

表1 三華李ANS 基因克隆和表達所用引物
1.2.3PsANS基因生物信息學分析 利用NCBI中的BLASTN、BLASTP 進行基因和蛋白序列比對;通過Expasy-ProtParam Tool 在線軟件對蛋白的基本理化性質進行預測,初步分析目的基因的功能;親疏水性利用ProtScale 以Hphob./Kyte &Doolittle 算法進行預測;采用TMHMM 2.0 進行跨膜結構預測;采用SignalP-5.0 預測蛋白質有無信號肽;采用ProtComp Version 9.0 進行蛋白亞細胞定位預測;采用NCBI 的Conserved Domains 分析功能結構域;使用SMART 在線數據庫分析蛋白結構域;蛋白質的二級結構采用ExPaSy-SOPMA在線軟件進行分析;蛋白質三級結構采用SWISSMODEL 進行在線預測。 利用在線軟件STRING預測ANS 與其他蛋白之間的互作情況(以擬南芥蛋白數據庫為參考);蛋白序列和同源比對分別使用DNAMAN 和ClustaX 軟件。 采用MEGA 5 軟件的Neighbor-Joining 法構建系統進化樹。
1.2.4PsANS表達分析 根據獲得的PsANS基因cDNA 序列設計熒光定量PCR 特異引物(表1)。 以三華李莖、葉、果皮、果肉不同組織以及三華李后熟過程中的樣品cDNA 為模板,參考TaKa-Ra 公司的SYBR Green qPCR Master Mix 說明書進行熒光定量PCR 擴增。 試驗設置3 個生物學重復,用2-ΔΔCt方法計算PsANS的相對表達量,采用SPSS 26 軟件進行LSD 方差分析。
2 結果與分析
2.1 三華李ANS 基因的克隆
三華李果實二代有參轉錄組參考物種為三月李(Prunus salicina),參考基因組版本為GCA_014863905.1_SCAU_Psal_1.0。 在三華李果實二代有參轉錄組數據庫中篩選初步注釋為ANS基因的轉錄本,其在轉錄組中的編號為evm.TU.UTG5995.3;在NCBI 中的比對結果表明,該片段與其他物種ANS基因的蛋白序列同源性在80%以上,其中與日本裸櫻(Prunus yedoensisvar.nudiflora, PQQ12998) 和 歐 洲 李(Prunus domestica,AHZ30597)ANS的蛋白序列同源性最高。 參考轉錄組中該基因序列設計引物,以三華李的cDNA為模板進行特異性PCR 擴增,獲得ANS基因的CDS 序列(圖1);將獲得的目的片段與pMD18-T載體連接并轉化DH5α 大腸桿菌,挑選陽性克隆測序。 經測序后發現該片段長度為1 074 bp,可編碼357 個氨基酸,將其命名為PsANS,GenBank登錄號為OP131916。 將該cDNA 片段與三月李基因組序列進行比對,發現該轉錄本對應的DNA序列在510~726 bp 間有1 個長度為217 bp 的內含子,其基因結構如圖2 所示。

圖1 三華李ANS 基因cDNA 片段瓊脂糖凝膠電泳圖譜
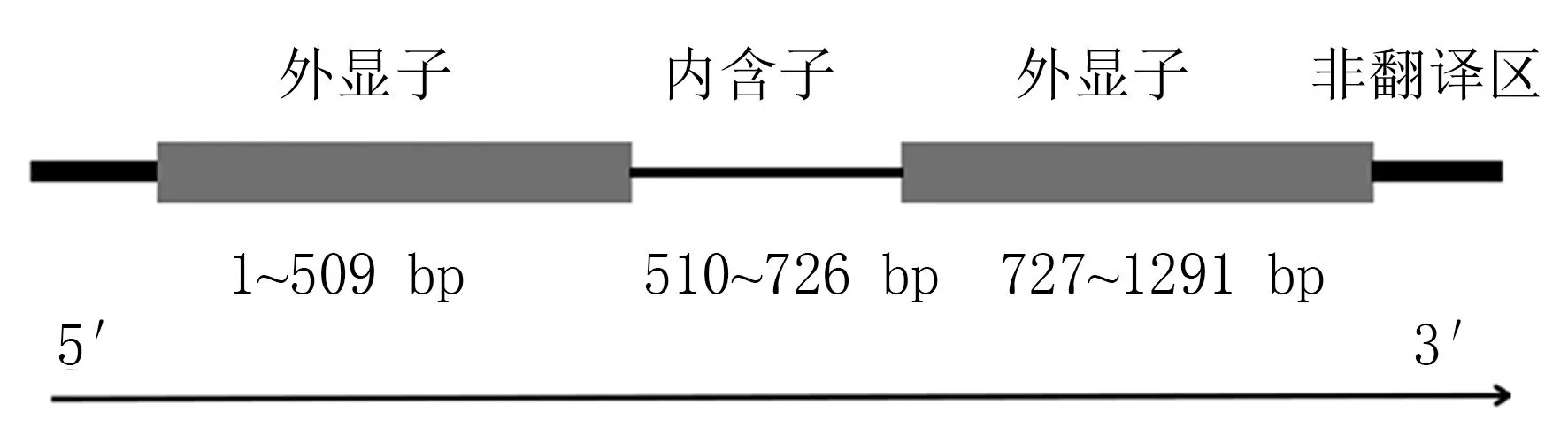
圖2 三華李ANS 基因DNA 結構圖
2.2 PsANS 蛋白理化性質分析
通過在線軟件Expasy-ProtParam Tool 分析,該蛋白質的分子式為C1901H3048N508O566S19,分子量為40 401.37,總原子數為5 717。 組成PsANS 蛋白的氨基酸中占比最多的是谷氨酸(Glu),達10.9%,其次為亮氨酸(Leu),占比10.1%。 PsANS蛋白的理論等電點為5.46,不穩定系數為47.96,屬于不穩定蛋白。
2.3 PsANS 蛋白親疏水性分析
預測發現(圖3),PsANS 的氨基端和羧基端表現出疏水性,親水性大部分集中在中心區域;疏水出現在第191 位的氨基酸殘基,親水出現在第65 位氨基酸殘基。 PsANS 蛋白的親水性總平均值(GRAVY)為-0.371,預測該蛋白屬于親水性蛋白。 PsANS 蛋白不存在跨膜區,無信號肽,為非分泌蛋白。 通過ProtComp Version 9.0 在線軟件預測PsANS 蛋白在植物細胞中可能定位在細胞質。

圖3 PsANS 蛋白疏水/親水性預測
2.4 PsANS 蛋白二級和三級結構預測
在線軟件ExPaSy-SOPMA 預測PsANS 蛋白的二級結構,結果(圖4)表明該蛋白二級結構中α-螺旋(h)占35.29%,β-折疊(t)占5.32%,無規則卷曲(c)占41.18%,延伸鏈(e)占18.21%。 利用SWISS-MODEL 以擬南芥ANS 蛋白為模板構建PsANS 蛋白的三維結構模型(圖5)。 以擬南芥蛋白數據庫為參考,通過STRING 預測PsANS 潛在的互作關系,結果(圖6)表明,ANS 蛋白主要與TT8(LDOX,無色花青素雙加氧酶)、TT4(CHS,查爾酮合成酶)、DFR(二氫黃酮醇-4-還原酶)、UF3GT(類黃酮3-O-葡萄糖基轉移酶)等存在互作關系。

圖4 PsANS 蛋白二級結構預測
圖5 PsANS 編碼蛋白三級結構模型

圖6 ANS 蛋白互作網絡模型預測
2.5 蛋白序列保守結構域、同源比對和系統進化分析
利用NCBI-CDD 對PsANS 蛋白的結構域進行分析,顯示PsANS 為PLN03178(花青素合成酶)多域蛋白,有1 個保守結構域,屬于2-酮戊二酸-Fe2+-雙加氧酶家族[2OG-Fe(Ⅱ)-Oxy,第216 ~311位];該保守結構域含有與2-酮戊二酸特異性結合的精氨酸Arg(R293,302)和絲氨酸Ser(S240,270,283,289,304)位點7 個,與Fe2+結合具有雙加氧功能的組氨酸His(H236,238,247,274,292)和天冬氨酸Asp(D277)位點6個,這些位點在不同物種的ANS 序列中高度保守。通過與其他12 種植物的ANS 蛋白進行同源性多重比對發現,ANS 氨基酸序列較為保守(圖7),PsANS 與同為薔薇科的歐洲李更近源(圖8)

圖7 PsANS 與其他已知ANS 蛋白的同源性比較

圖8 三華李與其他植物ANS 蛋白系統進化樹
2.6 PsANS 在三華李不同組織中的表達分析
植物次生代謝合成相關基因往往具有特異性的時空表達模式,即在代謝旺盛的組織中,相關基因的表達水平較高[10]。 由圖9 可見,PsANS在三華李的4 個不同組織中均有表達,其表達量在三華李果肉中最高,其余依次是果皮、葉、莖,各組織間存在顯著的表達差異。

圖9 PsANS 在三華李不同組織中的表達
2.7 PsANS 在三華李果實后熟過程中的表達分析
利用熒光定量PCR 技術分析PsANS在三華李后熟過程中的表達變化,結果(圖10)顯示,對照組和乙烯處理組PsANS表達量在后熟過程中都增加,貯藏6 d 的表達量分別是0 d 的3.6 倍和6.2 倍。 與對照組相比,乙烯處理組PsANS表達顯著上調(P<0.05)。 貯藏4 d 時,乙烯處理組的PsANS達到表達高峰,是0 d 的8.9 倍。
3 討論與結論
三華李因富含花青素等抗氧化活性物質深受廣大消費者喜愛和科研工作者關注[7]。 研究三華李果實花青素合成關鍵基因可為培育具有優良外觀品質的李子品種提供理論依據。 花青素合成酶(ANS)是花青素合成通路末端的酶,參與花青素的合成和累積,在果實著色及花色形成中具有重要作用[12]。
本研究首次從三華李果實中克隆獲得ANS基因,并將其命名為PsANS,該序列的編碼區長度為1 074 bp,可編碼一個由357 個氨基酸組成的蛋白質;與同屬薔薇科的蘋果梨[14]ANS基因長度一致。 本研究發現ANS 的氨基酸序列在不同植物中具有較高的保守性,都具有典型的2OG-Fe(Ⅱ)-Oxy 家族保守結構域和花青素合成酶催化位點Arg、Ser、His、Asp。 PsANS 與其他植物ANS蛋白性質相似,為無跨膜區、無信號肽的不穩定親水性蛋白,是一種非分泌性蛋白[19]。 花青素主要貯藏于植物細胞的液泡中,但其合成主要在細胞質的內質網表面進行,需經各種修飾后才被運至液泡等部位儲存[20],因此,包括ANS 在內的一系列酶集中形成多酶復合體在細胞質中催化花青素的合成[21]。 本研究預測發現PsANS 也定位于細胞質中。
以往研究發現ANS基因表達具有明顯的組織特異性,在植物體的各個組織中均有表達,但表達量存在差異[12,22,23]。 本研究發現,PsANS基因在三華李不同組織中均有表達,果肉中的表達量最高,莖中最低,暗示PsANS在三華李果肉花青素合成過程中扮演重要角色。 花青素含量是判斷三華李果實成熟度的顯著標志,在成熟過程中快速上升。 三華李是呼吸躍變型果實,后熟過程中果肉的花青素快速合成,而外源乙烯處理能促進果實快速軟化和轉紅。 以往研究表明外源乙烯對果實花青素合成有顯著影響,例如乙烯處理可上調桑椹ANS的表達并促進果實花青素合成[24];但紅梨和桃果實的花青素合成受乙烯處理顯著抑制[25,26]。 可見,乙烯對不同植物花青素合成的影響較為復雜。 本研究發現,三華李果實后熟過程中PsANS表達呈上升趨勢,外源乙烯處理進一步促進其表達,表明PsANS響應乙烯信號,在李果實后熟轉色過程中起重要作用。 但乙烯信號途徑和花青素合成途徑中轉錄因子間的作用機制還需進一步研究。
本研究首次克隆得到三華李果實花青素合成酶基因PsANS全長cDNA,該序列長度為1 074 bp,編碼357 個氨基酸。 PsANS 蛋白具有花青素合成酶的典型結構特征,氨基酸序列與日本裸櫻和歐洲李的ANS 氨基酸序列相似度高;三華李花青素合成酶基因的表達存在組織器官差異性,果肉中表達量最高,莖中最低;三華李后熟轉色過程中PsANS表達呈上升趨勢,乙烯處理進一步上調了PsANS的表達,表明其對三華李果實發育和后熟過程中花青素的合成起著重要作用。
