NR2F2基因調(diào)控豬PK15細(xì)胞增殖和凋亡的研究
2023-08-25 09:03:06張萬鋒趙天枝尤紫薇蔡春波高鵬飛曹果清郭曉紅李步高
畜牧獸醫(yī)學(xué)報(bào) 2023年8期
張萬鋒,趙天枝,李 嬌,尤紫薇,楊 陽,蔡春波,高鵬飛,曹果清,郭曉紅,李步高
(山西農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)學(xué)院,太谷 030801)
細(xì)胞增殖是細(xì)胞重要的生命活動(dòng),也是機(jī)體生長、發(fā)育、繁殖和遺傳的基礎(chǔ)[1-3]。細(xì)胞增殖是一個(gè)復(fù)雜而有序的過程,主要受細(xì)胞周期因子和周期蛋白依賴激酶調(diào)控,這些基因由很多信號(hào)通路所調(diào)控,如Wnt信號(hào)通路、Notch信號(hào)通路、TGF-β信號(hào)通路,而這些通路又受到許多轉(zhuǎn)錄因子的調(diào)控[4-11]。
雞卵清蛋白上游啟動(dòng)子轉(zhuǎn)錄因子2(COUP-TFII,也叫NR2F2)是核受體家族超家族成員,在1986年因直接結(jié)合于卵清蛋白的啟動(dòng)子區(qū)調(diào)節(jié)其轉(zhuǎn)錄被發(fā)現(xiàn)[12]。在脊椎動(dòng)物中,雞卵清蛋白上游啟動(dòng)子轉(zhuǎn)錄因子家族由3個(gè)成員組成,核受體亞家族2組F成員1(NR2F1)、NR2F2和核受體亞家族2組F成員6(NR2F6)。雖然它們在不同的染色體上,但氨基酸序列是高度同源的[13]。NR2F2基因在組織中廣泛表達(dá),在骨骼、肌肉、脂肪形成和代謝平衡中起重要調(diào)節(jié)作用[14-16]。NR2F2作為一種轉(zhuǎn)錄因子,通過多種機(jī)制調(diào)控下游靶基因的轉(zhuǎn)錄。NR2F2可以通過與DNA結(jié)合元件結(jié)合,直接或間接激活基因表達(dá)。NR2F2可以與Wnt家族成員10b(Wnt10b)啟動(dòng)子區(qū)域結(jié)合,抑制Wnt10b的表達(dá)[17]。NR2F2可以與CCAAT增強(qiáng)子結(jié)合蛋白(C/EBPα)啟動(dòng)子結(jié)合,抑制C/EBPα的表達(dá)[18]。NR2F2可以與維甲酸X受體(RXR)的DNA結(jié)合位點(diǎn)競爭性結(jié)合[19]。NR2F2還作為一種轉(zhuǎn)錄抑制劑,可與RXR和甲狀腺激素受體相互作用,抑制PPAR的表達(dá)[20]。此外,NR2F2還可以通過直接結(jié)合RXR的LBD結(jié)構(gòu)域來抑制核受體的轉(zhuǎn)錄[21]。
為了確定NR2F2基因?qū)?xì)胞增殖的作用機(jī)制,Ma等[22]研究發(fā)現(xiàn),在WJ-MSCs細(xì)胞中敲除NR2F2基因后導(dǎo)致CyclinD1和CDK4表達(dá)降低,細(xì)胞增殖減慢。Qin等[23]對敲除NR2F2基因的PC3細(xì)胞與正常細(xì)胞進(jìn)行了轉(zhuǎn)錄組比較,通過基因富集發(fā)現(xiàn),在NR2F2基因表達(dá)低時(shí),TGF-β信號(hào)通路下游調(diào)控的基因全部富集,說明NR2F2能調(diào)控TGF-β信號(hào)通路。敲除NR2F2基因后,顯著改變了TGF-β信號(hào)通路下游基因p21、p15和CyclinD1的表達(dá)。Wang等[24]研究發(fā)現(xiàn),NR2F2基因在直腸癌細(xì)胞中與PTEN、Smad4基因的表達(dá)相關(guān),而Smad4基因是TGF-β信號(hào)通路的關(guān)鍵基因。Smad4基因主要功能是參與TGF-β信號(hào)通路的信號(hào)傳導(dǎo)。進(jìn)一步研究發(fā)現(xiàn),NR2F2蛋白能夠與Smad4蛋白結(jié)合,與突變型的Smad4不能結(jié)合,直接證明了NR2F2與Smad4之間的結(jié)合作用。表明在癌細(xì)胞中,NR2F2基因可以通過與Smad4基因相互作用,進(jìn)而影響TGF-β信號(hào)通路傳導(dǎo)。
目前,圍繞NR2F2基因的研究主要集中在其對人類癌癥等疾病、小鼠肌肉與脂肪等組織的作用機(jī)理,針對NR2F2基因調(diào)控豬源細(xì)胞增殖的機(jī)制還未見報(bào)道。本試驗(yàn)旨在探究NR2F2基因?qū)?xì)胞增殖、凋亡的影響,為進(jìn)一步研究豬生長發(fā)育的分子機(jī)制提供理論基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 細(xì)胞系和質(zhì)粒 本研究中使用的PK15細(xì)胞和293 T細(xì)胞來自山西農(nóng)業(yè)大學(xué)動(dòng)物遺傳育種與繁殖博士點(diǎn)實(shí)驗(yàn)室。
pLenti-CMV-GFP-SV-puro、pLenti-CMV-NR2F2-GFP-SV-puro、pCMV-dR8.91、pCMV-VSV-G、pCDNA3.1-Flag、pCDNA3.1-Flag-NR2F2、pHS-CR054、pHS-CR054-sg1、pHS-CR054-sg2質(zhì)粒均為山西農(nóng)業(yè)大學(xué)動(dòng)物遺傳育種實(shí)驗(yàn)室保存。
1.1.2 主要試劑 PrimeScriptTMRT Master Mix、RNAiso Plus regent和SYBR Premix Ex Taq II購于TaKaRa公司;PBS粉末、青鏈霉素混合液、胰蛋白酶購于中國索萊寶公司;FBS和DMEM購自Gibco公司;兔二抗、鼠二抗購于LI-COR公司;Lipofectamine 3000購自Invitrogen公司;CCK-8試劑盒購自Dojindo公司;EdU試劑盒、細(xì)胞周期試劑盒、細(xì)胞凋亡試劑盒購自凱基生物公司;RIPA裂解液(強(qiáng))、蛋白酶抑制劑、蛋白凝膠試劑盒、NC膜、脫脂奶粉購自博士德公司;β-actin抗體購自博奧森公司;NR2F2抗體購自Abcam公司;PCNA抗體購自CST公司;siRNA購自吉瑪公司,siRNA序列見表1。

表1 siRNA序列
1.2 試驗(yàn)方法
1.2.1 慢病毒包裝及轉(zhuǎn)染細(xì)胞 轉(zhuǎn)染前1 d,胰酶消化293 T細(xì)胞,接種細(xì)胞于10 cm細(xì)胞培養(yǎng)皿,培養(yǎng)1 d,當(dāng)細(xì)胞濃度達(dá)到60%~80%時(shí)進(jìn)行轉(zhuǎn)染。用Lipofectamine 3000將10 μg pLenti-CMV-GFP-SV-puro或pLenti-CMV-NR2F2-GFP-SV-puro質(zhì)粒與7.5 μg pCMV-dR8.91、5 μg pCMV-VSV-G包裝質(zhì)粒共轉(zhuǎn)染293 T細(xì)胞中,轉(zhuǎn)染6 h后,更換為完全培養(yǎng)基,繼續(xù)培養(yǎng)48、72 h后,收集293 T細(xì)胞培養(yǎng)液,4 ℃、1 000 r·min-1離心5 min,去除細(xì)胞碎片,0.45 μm針式濾器過濾,收集病毒液置于-80 ℃冰箱,用于感染PK15細(xì)胞。
將PK15細(xì)胞接種于6孔板中,待細(xì)胞匯合度達(dá)到50%~60%,每孔加入1 mL病毒上清液,感染6 h后,更換為完全培養(yǎng)基培養(yǎng),感染72 h后,熒光顯微后熒光顯微鏡檢測EGFP的表達(dá),形成穩(wěn)定轉(zhuǎn)染NR2F2基因的細(xì)胞。
1.2.2 質(zhì)粒和siRNA轉(zhuǎn)染 PK15細(xì)胞接種至10 cm培養(yǎng)皿中,使用含10%FBS和1%雙抗的低糖DMEM培養(yǎng)基在37 ℃,5% CO2條件下進(jìn)行培養(yǎng)。將細(xì)胞進(jìn)行消化計(jì)數(shù),接種至6孔板中,細(xì)胞生長匯合至40%~50%即可進(jìn)行轉(zhuǎn)染。利用Lipofectamine 3000脂質(zhì)體分別將pHS-CR054、pHS-CR054-sg1、pHS-CR054-sg2質(zhì)粒、siRNA-NC及siRNA-Smad4轉(zhuǎn)染至PK15細(xì)胞中,每組3個(gè)重復(fù)。按轉(zhuǎn)染試劑操作說明,取兩個(gè)1.5 mL離心管,分別加入125 μL DMEM培養(yǎng)液,然后于其中一管加入100 pmol siRNA或4 μg質(zhì)粒吹打混勻;另一管加入10 μL轉(zhuǎn)染試劑吹打混勻,室溫靜置5 min后,將含有siRNA或質(zhì)粒的培養(yǎng)液加入含轉(zhuǎn)染試劑的培養(yǎng)液中吹打混勻,室溫靜置20 min。將制備好的轉(zhuǎn)染溶液加至6孔板,并用DMEM加至每孔2 mL,于37 ℃,5% CO2中培養(yǎng),6 h后更換為含10%胎牛血清的培養(yǎng)液。轉(zhuǎn)染后觀察結(jié)果并收取細(xì)胞進(jìn)行后續(xù)試驗(yàn)。
1.2.3 RNA提取及實(shí)時(shí)熒光定量PCR 使用RNAiso Plus提取總RNA,提取方法參照試劑盒說明書。將提取的總RNA用核酸蛋白測定儀測定其純度及濃度,OD260nm/OD280 nm為1.8~2.0的總RNA用于后續(xù)研究,采用PrimeScriptTMRT Master Mix試劑盒將RNA反轉(zhuǎn)錄成cDNA。
以18 S rRNA為內(nèi)參基因,采用qRT-PCR對基因的表達(dá)水平進(jìn)行檢測。qRT-PCR反應(yīng)體系:cDNA 2 μL,2×SYBR Premix Ex TaqⅡ10 μL,上、下游引物(表2)各0.5 μL,RNAase Free ddH2O補(bǔ)至20 μL。反應(yīng)程序:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,35個(gè)循環(huán);熔解曲線程序?yàn)?5 ℃ 15 s,60 ℃ 35 s,95 ℃。結(jié)果采用2-ΔΔCt法進(jìn)行分析,每個(gè)樣本做3次重復(fù)。

表2 qRT-PCR引物序列
1.2.4 總蛋白提取和Western blot檢測 提取細(xì)胞總蛋白,利用核酸蛋白濃度測定儀測定蛋白濃度。取200 ng蛋白上樣,進(jìn)行聚丙烯酰胺凝膠電泳。電泳結(jié)束后,采用濕轉(zhuǎn)法進(jìn)行轉(zhuǎn)膜,5%封閉蛋白粉室溫封閉1 h,一抗采用1∶1 000稀釋,4 ℃孵育過夜。隔天回收一抗,PBST洗膜3次,加入熒光二抗,避光室溫孵育1 h,洗膜后使用LICOR儀器曝光,使用儀器自帶軟件Image Studio計(jì)算分析條帶光密度值。
1.2.5 CCK-8、EdU試驗(yàn) 將細(xì)胞接種至96孔細(xì)胞板中,每孔細(xì)胞約1 000個(gè),在鋪板后0、1、2、3、4、5 d分別進(jìn)行檢測,檢測前每孔加入10 μL的CCK-8試劑,在細(xì)胞培養(yǎng)箱孵育3 h后,使用酶標(biāo)儀檢測450 nm吸光度。
取處于對數(shù)生長期的細(xì)胞,以每孔4 000~10 000個(gè)細(xì)胞接種于96孔板中,每孔加入100 μL EdU染液和100 μL Hoechst33342溶液,染色完畢后使用熒光顯微鏡成像。
1.2.6 流式細(xì)胞術(shù)檢測 將細(xì)胞接種至6 cm細(xì)胞培養(yǎng)皿,培養(yǎng)至細(xì)胞濃度80%~90%時(shí),收集的細(xì)胞在70%冷乙醇中重懸,4 ℃固定過夜,復(fù)溫后加入400 μL PI染色,4 ℃避光孵育30 min,流式細(xì)胞儀檢測。
將細(xì)胞接種至6 cm細(xì)胞培養(yǎng)皿,培養(yǎng)至細(xì)胞濃度80%~90%時(shí),PBS洗2次,收集細(xì)胞;加入500 μL的Binding Buffer懸浮細(xì)胞,加入5 μL Annexin V-APC混勻后,加入5 μL 7-AAD染液,混勻,室溫、避光、孵育15 min;流式細(xì)胞儀檢測。
1.2.7 傷口愈合試驗(yàn) 將細(xì)胞接種至6孔板中,待其長滿后,用1 mL藍(lán)槍頭在六孔板內(nèi)劃線,PBS清洗2次,加培養(yǎng)基繼續(xù)培養(yǎng),觀察0、12、24、36、48 h時(shí)劃痕處細(xì)胞的生長狀態(tài)。
1.2.8 CO-IP試驗(yàn) 收集轉(zhuǎn)染了過表達(dá)NR2F2且攜帶Flag標(biāo)簽蛋白載體的PK15細(xì)胞,加入40 μL Flag樹脂的裂解液在4 ℃下輕輕搖晃孵育過夜,然后用500 μL細(xì)胞裂解緩沖液漂洗樹脂,混勻儀上室溫漂洗5 min,4 ℃ 500 r·min-1離心5 min,重復(fù)3次,加入50 μL洗脫緩沖液,漩渦震蕩20 s,放搖勻儀上,室溫洗脫10~15 min,漩渦震蕩20 s,4 ℃ 12 000 r·min-1離心5 min,取上清到新的離心管中,用于Western blot。
1.2.9 數(shù)據(jù)分析 應(yīng)用SPSS 22.0進(jìn)行統(tǒng)計(jì)分析。數(shù)據(jù)采用單因素方差分析和獨(dú)立樣本t檢驗(yàn)進(jìn)行比較,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結(jié) 果
2.1 慢病毒包裝及NR2F2基因過表達(dá)效率
病毒液感染PK15細(xì)胞48 h后,熒光顯微鏡觀察發(fā)現(xiàn)OE-NC組和OE組都表達(dá)了綠色熒光蛋白(圖1A)。qRT-PCR和WB檢測過表達(dá)NR2F2基因的效率,結(jié)果發(fā)現(xiàn),慢病毒液感染PK15細(xì)胞后,NR2F2基因mRNA水平和蛋白水平極顯著增加(P<0.01,圖1B~D)。

A.綠色熒光蛋白EGFP表達(dá)(標(biāo)尺為100 μm);B~D.過表達(dá)NR2F2基因mRNA和蛋白水平表達(dá)。“**”表示差異極顯著(P<0.01),“*”表示差異顯著(P<0.05)。下同
2.2 過表達(dá)NR2F2基因?qū)?xì)胞增殖、凋亡的影響
慢病毒感染PK15細(xì)胞后,結(jié)果顯示Ki67基因mRNA表達(dá)極顯著增加(P<0.01,圖2A);qRT-PCR、WB檢測結(jié)果顯示,PCNA表達(dá)量顯著增加(P<0.05,圖2A、2B、2C)。CCK-8結(jié)果顯示,過表達(dá)NR2F2基因后細(xì)胞的吸光度值隨著時(shí)間的增加極顯著上升(P<0.01),差異具有統(tǒng)計(jì)學(xué)意義,表明促進(jìn)了細(xì)胞的增殖(圖2D)。EdU結(jié)果顯示,過表達(dá)NR2F2基因后S期細(xì)胞數(shù)量顯著增多(P<0.01),表明促進(jìn)了細(xì)胞的增殖(圖2E、F)。流式細(xì)胞儀檢測過表達(dá)NR2F2基因后細(xì)胞周期變化,發(fā)現(xiàn)過表達(dá)NR2F2基因后,處于G1期細(xì)胞的數(shù)量顯著低于對照組(P<0.01),S期細(xì)胞的數(shù)量顯著高于對照組(P<0.01),表明NR2F2基因可以改變細(xì)胞周期,促進(jìn)G1期向S期轉(zhuǎn)變,從而促進(jìn)細(xì)胞增殖(圖2G、H)。通過流式細(xì)胞儀檢測過表達(dá)NR2F2基因?qū)?xì)胞凋亡的影響,結(jié)果發(fā)現(xiàn),過表達(dá)NR2F2基因后,早凋細(xì)胞占比增多,晚凋細(xì)胞占比減少,表明NR2F2基因可以抑制細(xì)胞的凋亡(圖2I)。通過劃痕試驗(yàn)檢測NR2F2基因?qū)?xì)胞遷移的影響,結(jié)果表明,過表達(dá)NR2F2基因后,細(xì)胞遷移速度加快,在48 h后基本匯合,而對照組細(xì)胞還未完全融合(圖2J)。
2.3 NR2F2與Smad4物理結(jié)合并調(diào)控其表達(dá)進(jìn)而影響下游基因的表達(dá)
為了確定NR2F2基因調(diào)控細(xì)胞增殖的具體機(jī)制,本研究通過蛋白相互作用軟件預(yù)測了一些與NR2F2相互作用的蛋白。發(fā)現(xiàn)能與NR2F2互作的蛋白有15個(gè),其中我們將關(guān)注點(diǎn)聚焦在Smad4蛋白上(圖3A)。進(jìn)行了Co-IP分析來檢測NR2F2和Smad4之間的相互作用。用Flag標(biāo)簽蛋白調(diào)取與它結(jié)合的復(fù)合物,能檢測到Smad4蛋白,說明NR2F2蛋白能與Smad4蛋白相互作用行使功能(圖3B)。
為了探究NR2F2基因是否調(diào)控Smad4基因的表達(dá),檢測了過表達(dá)和敲除NR2F2基因?qū)mad4基因表達(dá)的影響,發(fā)現(xiàn)過表達(dá)NR2F2基因后,Smad4基因表達(dá)量升高(P<0.01,圖4A),下游基因CyclinB、CDK1、CDK4、CyclinD1表達(dá)量均顯著上升(P<0.01,P<0.05,圖4C);而敲除NR2F2基因后,Smad4基因表達(dá)量降低(P<0.01,圖4B),下游基因CyclinB、CDK1、CDK4、CyclinD1表達(dá)量均顯著下降(P<0.01,圖4D)。

A.過表達(dá)NR2F2后Smad4的表達(dá)變化;B.敲除NR2F2后Smad4的表達(dá)變化;C.過表達(dá)NR2F2后Smad4調(diào)控的下游基因的表達(dá)變化;D.敲除NR2F2后Smad4調(diào)控的下游基因的表達(dá)變化。不同大寫字母表示差異極顯著(P<0.01);不同小寫字母表示差異顯著(P<0.05),下同
挽救試驗(yàn)中,在過表達(dá)NR2F2基因后,干擾Smad4基因表達(dá),Smad4表達(dá)量顯著降低(P<0.01,圖5A),且能夠顯著緩解過表達(dá)NR2F2基因引起的Smad4下游基因CyclinD1、CyclinB、CDK1的表達(dá)的升高,降低了它們的表達(dá)量(P<0.01,P<0.05,圖5B)。通過檢測增殖相關(guān)基因的表達(dá),結(jié)果發(fā)現(xiàn),干擾Smad4基因表達(dá),能緩解過表達(dá)NR2F2基因?qū)?xì)胞增殖的影響,PCNA和Ki67的表達(dá)量顯著降低(P<0.01,P<0.05,圖5C)。
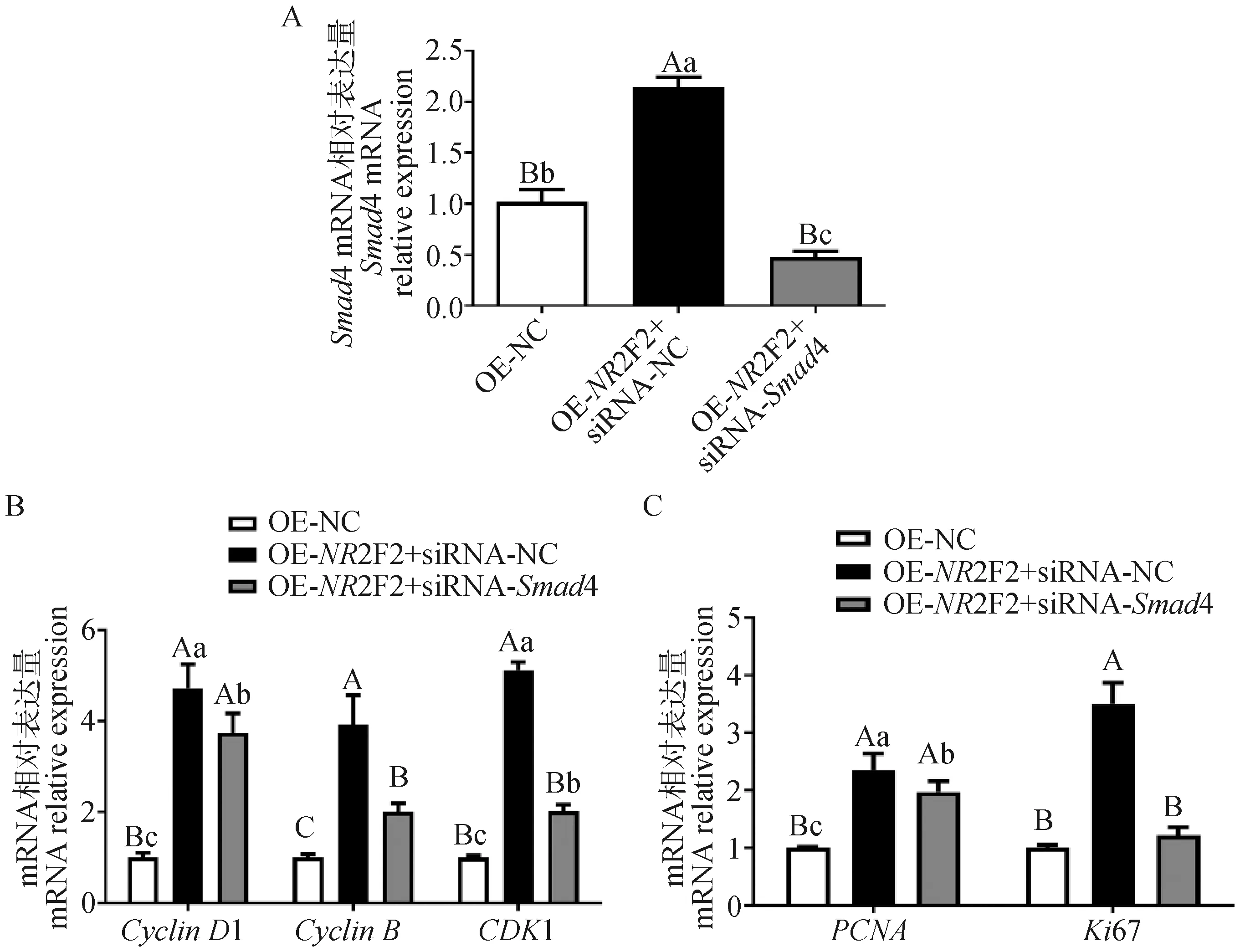
A.干擾Smad4的效率;B.過表達(dá)NR2F2且干擾Smad4后下游基因表達(dá)的變化;C.過表達(dá)NR2F2且干擾Smad4后增殖基因表達(dá)的變化
3 討 論
NR2F2作為一種轉(zhuǎn)錄因子,在細(xì)胞的生命過程中起著重要的作用。嚙齒類動(dòng)物的遺傳模型揭示了NR2F2基因在脂肪形成、脂質(zhì)代謝、肝臟糖異生、胰島素分泌和血壓調(diào)節(jié)等生物過程中的重要作用[25-27]。本研究發(fā)現(xiàn),在PK15細(xì)胞中,NR2F2基因促進(jìn)細(xì)胞增殖。NR2F2基因?qū)K15細(xì)胞的生長促進(jìn)作用是通過加速細(xì)胞周期從G1期到S期的進(jìn)程來介導(dǎo)的。這些結(jié)果表明,NR2F2基因能促進(jìn)細(xì)胞增殖。在之前的研究中,NR2F2已被證明在細(xì)胞的增殖中發(fā)揮促進(jìn)作用。NR2F2在多種腫瘤細(xì)胞中高表達(dá),促進(jìn)細(xì)胞增殖,抑制細(xì)胞凋亡[28]。在腎細(xì)胞癌細(xì)胞中,下調(diào)NR2F2可通過上調(diào)BRCA1來抑制細(xì)胞增殖并誘導(dǎo)細(xì)胞凋亡[29]。NR2F2通過調(diào)節(jié)IGF-1表達(dá)來調(diào)節(jié)小鼠出生后小腦的生長和成熟[30]。過表達(dá)NR2F2基因可改變細(xì)胞粘附蛋白和細(xì)胞增殖的表達(dá)[31]。敲除NR2F2后,Ki67基因表達(dá)減少,細(xì)胞增殖被抑制,凋亡細(xì)胞增加[32]。NR2F2沉默后,細(xì)胞增殖和侵襲減少,細(xì)胞被阻斷在G1期[33]。可見,NR2F2基因在細(xì)胞增殖中起著重要作用,但其具體的作用機(jī)制還需進(jìn)一步研究。
NR2F2基因?qū)儆诤耸荏w轉(zhuǎn)錄因子家族。作為一種轉(zhuǎn)錄因子,NR2F2不直接調(diào)節(jié)激酶活性。NR2F2可能通過調(diào)節(jié)其他基因的表達(dá)來調(diào)節(jié)激酶活性。在不同的細(xì)胞模型中,ERα、CEBP/α、Wnt10b已被證明受NR2F2的調(diào)控。在本研究中,通過蛋白質(zhì)相互作用軟件預(yù)測了一些與NR2F2相互作用的蛋白質(zhì),發(fā)現(xiàn)有15個(gè)蛋白可以與NR2F2相互作用,其中,Smad4比較值得注意。本研究采用Co-IP直接驗(yàn)證了PK15細(xì)胞中NR2F2蛋白與Smad4蛋白的結(jié)合。而過表達(dá)NR2F2可提高Smad4的水平。NR2F2的沉默可降低Smad4的水平。這說明NR2F2可能改變了Smad4基因的表達(dá)。
Smad4為抑癌基因,屬于Smad家族。轉(zhuǎn)化生長因子-β(TGF-β)信號(hào)可以通過以下反應(yīng)激活:1)TGF-β與受體結(jié)合,然后誘導(dǎo)Smad2/3磷酸化;2)共同介質(zhì)Smad4與磷酸化的Smads結(jié)合,然后遷移到細(xì)胞核;3)Smad復(fù)合物與各種轉(zhuǎn)錄因子相互作用,然后調(diào)節(jié)細(xì)胞的增殖和遷移。先前的研究表明,Smad4的缺失會(huì)影響細(xì)胞的增殖和遷移。敲除Smad4的細(xì)胞增殖較慢,細(xì)胞遷移減少[34-35]。miR-144可以靶向Smad4基因,降低Smad4基因的表達(dá),從而抑制細(xì)胞增殖和遷移[36]。Smad4基因表達(dá)水平的降低增加了對DNA拓?fù)洚悩?gòu)酶抑制劑的敏感性,促進(jìn)細(xì)胞凋亡,從而抑制細(xì)胞增殖[37]。這些研究表明Smad4可以調(diào)節(jié)細(xì)胞周期。
TGF-β信號(hào)通路在組織發(fā)育、穩(wěn)態(tài)和修復(fù)中起著重要作用,并調(diào)控細(xì)胞增殖、分化、凋亡和遷移等細(xì)胞過程[38-39]。Smad4作為唯一的通用受體和TGF-β信號(hào)通路的中介,在將受體信號(hào)轉(zhuǎn)導(dǎo)到核中的靶基因中起核心作用[40]。Smad4的異常表達(dá)可引起細(xì)胞中TGF-β超家族成員的異常變化,影響下游靶基因的表達(dá),并影響細(xì)胞的生長發(fā)育。細(xì)胞增殖周期相關(guān)因子是TGF-β信號(hào)通路的下游效應(yīng)因子[41]。研究發(fā)現(xiàn),敲除NR2F2可以顯著下調(diào)細(xì)胞周期相關(guān)基因的表達(dá);而過表達(dá)NR2F2可顯著上調(diào)細(xì)胞增殖周期相關(guān)基因的表達(dá)。挽救試驗(yàn)中過表達(dá)NR2F2和干擾Smad4后,可以有效緩解過表達(dá)NR2F2基因的影響。這說明NR2F2通過與Smad4相互作用,影響TGF-β信號(hào)通路,從而影響細(xì)胞增殖周期相關(guān)因子的表達(dá)。因此,發(fā)現(xiàn)NR2F2通過影響Smad4基因的表達(dá)來影響細(xì)胞增殖。
4 結(jié) 論
過表達(dá)NR2F2基因后促進(jìn)了細(xì)胞的增殖,抑制了凋亡,且NR2F2蛋白能與Smad4蛋白結(jié)合并影響Smad4基因的表達(dá),進(jìn)而影響下游基因的表達(dá)。
猜你喜歡
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
鴨綠江(2021年35期)2021-04-19 12:24:18
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25

- 畜牧獸醫(yī)學(xué)報(bào)的其它文章
- 單寧酸對低劑量T-2毒素誘導(dǎo)小鼠結(jié)腸黏膜損傷與菌群失調(diào)的保護(hù)效應(yīng)
- 基于網(wǎng)絡(luò)藥理學(xué)和試驗(yàn)驗(yàn)證分析小檗堿治療雞沙門菌感染的作用機(jī)制
- 內(nèi)質(zhì)網(wǎng)應(yīng)激預(yù)適應(yīng)對LPS誘導(dǎo)的山羊子宮內(nèi)膜上皮細(xì)胞炎性反應(yīng)的保護(hù)作用
- 基于RNA-Seq技術(shù)研究枸杞多糖對環(huán)磷酰胺致雛雞免疫抑制的拮抗機(jī)制
- 阿司匹林丁香酚酯顆粒劑有關(guān)物質(zhì)檢測方法的建立與驗(yàn)證
- 黑果枸杞花青素對低氧誘導(dǎo)的H9c2大鼠心肌細(xì)胞凋亡的影響