TGFβR1介導TGF-β/Smad信號通路對綿羊顆粒細胞功能的影響
2023-08-25 09:03:10李悅欣劉愛菊馬曉菲胡伯欣智云霞田樹軍
畜牧獸醫學報 2023年8期
李悅欣,劉愛菊,馬曉菲,鄭 忠,胡伯欣,智云霞,田樹軍*
(1.河北農業大學動物科技學院,保定 071000;2.滄州職業技術學院農牧工程系,滄州 061000)
綿羊的排卵數/胎產羔數與卵泡的生長發育和閉鎖密切相關[1],顆粒細胞作為卵泡內數量最多、比例最大以及功能最全的細胞群,其增殖、凋亡、分化和激素生成等對卵泡生長發育及卵泡閉鎖至關重要[2-3]。本課題組最新研究發現,TGF-β信號通路在具有高繁殖力的綿羊卵泡內發揮正調控作用[4]。TGFβR1作為TGF-β信號通路的核心信號分子,是否通過介導TGF-β/Smad分支信號通路調控綿羊卵泡內顆粒細胞的功能,目前尚不清楚。
TGF-β/Smad是TGF-β信號通路的一個重要分支,其中磷酸化的TGFβ受體活性因子(TGFβR1)作為核心信號分子,會依次與SMAD2/3結合并將其活化,共同與SMAD4結合后調控下游基因的表達。在小鼠和豬等哺乳動物的卵巢顆粒細胞中,TGF-β信號通路的激活可促進顆粒細胞增殖[5-7],而失活則會導致顆粒細胞凋亡和卵泡閉鎖[8-10]。Zhang等[11]研究表明,TGFβR1和SMAD3的表達受到阻礙時,導致豬顆粒細胞的生長和分化減少,并致使細胞凋亡。現有研究發現,SMAD3基因敲除會導致大鼠雌激素產生減少和顆粒細胞凋亡增加[12]。Yu和Liu[13]的研究發現,SMAD2是miR-30d-5p的靶基因,SMAD2過表達增強了大鼠卵巢顆粒細胞增殖并抑制了細胞凋亡。SMAD4是細胞核中TGF-β信號通路的介體和效應器,調節眾多細胞過程,包括細胞增殖、分化、自身免疫、多能性和可塑性以及細胞生長、凋亡、自噬、侵襲和轉移[14-17]。SMAD4失調與胚胎、骨骼肌分化缺陷、干細胞丟失、神經系統發育障礙和延遲、女性不育和其他疾病有關[18-22]。
為此,本試驗擬構建pcDNA3.1(+)-TGFβR1和pcDNA3.1(+)-SMAD4過表達質粒載體及shRNA-TGFβR1和shRNA-SMAD4干擾表達質粒載體,對綿羊顆粒細胞中TGFβR1和SMAD4基因分別進行過表達和干擾,利用CCK-8細胞增殖檢測、流式細胞術、Annexin-V FITC/PI雙染技術檢測綿羊顆粒細胞增殖、周期及凋亡功能,并結合Real-Time PCR及Western blot檢測TGFβR1、SMAD4的mRNA水平和TGF-β/Smad信號通路中TGFβR1、SMAD2/3等的磷酸化水平,探究TGFβR1、SMAD4對顆粒細胞增殖、凋亡、周期及TGF-β信號通路的影響,進而揭示TGFβR1介導TGF-β/Smad信號通路調控綿羊卵泡內顆粒細胞功能的機制。
1 材料與方法
1.1 試驗材料
1.1.1 綿羊卵巢 從河北省保定市唐縣屠宰場采集綿羊卵巢,將其放入加有1%雙抗的37 ℃生理鹽水中,于3 h內運回實驗室。
1.1.2 主要試劑 胎牛血清、胰酶、雙抗、0.25% Tripsin-EDTA均購自Gibco公司;DME/F-12、PBS購自Hyclone公司;TRIzol?Plus RNA Purification Kit購自Invitrogen公司;RNase-Free DNase Set購自Qiagen公司;SuperScriptTMIII First-Strand Synthesis SuperMix for qRT-PCR、Power SYBR?Green PCR Master Mix購自Roche公司;30%丙烯酰胺溶液購自Bio-Rad公司;PVDF轉印膜購自Millipore公司;ECL DualVue WB Marker購自GE公司;SuperSignal West Dura Extended Duration Substrate購自Thermo Pierce公司;X-ray film購自華東醫藥;細胞周期檢測試劑盒購自碧云天生物技術公司;Annexin V-FITC/PI細胞凋亡檢測試劑盒購自江蘇凱基生物。
1.1.3 主要儀器設備 二氧化碳培養箱購自Thermo Fisher公司;Quantstudio多重實時熒光定量PCR儀購自美國life technologies公司;Mini-PROTEAN電泳系統、Mini Trans-Blot轉印系統購自美國Bio-Rad公司;流式細胞儀購自美國 Becton-Dickinson FACS Calibur公司。
1.2 試驗方法
1.2.1 顆粒細胞采集及培養 剪下屠宰綿羊的卵巢,用75%酒精噴洗,放于37 ℃滅菌生理鹽水(含雙抗)的保溫瓶中,于4 h內回到實驗室,進行顆粒細胞采集。采用預熱37 ℃的生理鹽水沖洗卵巢,使用滅菌剪刀剪掉卵巢周圍系膜,生理鹽水反復沖洗至干凈,在DME/F-12培養基(含10%胎牛血清)中用無菌刀片劃破3~7 mm的卵泡,將卵泡液和培養基混合液轉移至離心管(10 mL)中,1 500 r·min-1離心8 min,棄上清,PBS清洗兩次,加入DME/F-12培養基(含10%胎牛血清)吹打混勻后接種于培養皿中,置于37 ℃,50 mL·L-1CO2培養箱中培養。參照韓紅葉[23]的方法對顆粒細胞進行免疫熒光鑒定。
1.2.2 pcDNA3.1(+)-TGFβR1和pcDNA3.1(+)-SMAD4真核表達載體的構建 使用NheI和Hind III兩個限制性內切酶對pcDNA3.1質粒載體進行雙酶切線性化處理,并通過瓊脂糖凝膠電泳檢測酶切產物,切膠回收線性化的pcDNA3.1載體。通過PCR擴增獲取TGFβR1和SMAD4基因的目的片段,引物由上海桑尼生物合成,TGFβR1與SMAD4引物序列見表1。PCR擴增程序:95 ℃ 3 min;95 ℃ 30 s,58 ℃ 30 s,72 ℃ 1 min,29個循環;72 ℃ 10 min。膠回收目的片段后進行重組反應,重組反應體系:目的片段1 μL,線性化載體1 μL,2×Super Fusion Mix 5 μL,H2O 3 μL,50 ℃水浴1 h完成重組反應。將重組產物轉化至感受態細胞DH5α中,將菌液涂板37 ℃恒溫箱培養12 h,挑取單克隆菌落37 ℃搖菌16 h,酶切驗證后將陽性菌液送公司測序鑒定。對符合條件的質粒命名為pcDNA3.1(+)-TGFβR1和pcDNA3.1(+)-SMAD4質粒載體(文中簡稱為pc3.1-TGFβR1和pc3.1-SMAD4),對照組為pcDNA3.1空白載體(文中簡稱為PC組)。

表1 PCR引物序列
1.2.3 shRNA-TGFβR1和shRNA-SMAD4質粒的構建 使用AgeI和EcoR I兩個限制性內切酶對pLKO.1質粒進行雙酶切線性化處理,并通過瓊脂糖凝膠電泳檢測酶切產物,切膠回收線性化的pLKO.1載體。TGFβR1和SMAD4的shRNA oligo序列信息見表2。shRNA oligo退火后得到雙鏈shRNA,使用T4連接酶將雙鏈shRNA和線性化的pLKO.1載體進行連接反應,連接反應體系:退火得到的雙鏈DNA片段2 μL,線性化載體1 μL,T4連接酶1 μL,T4 buffer 2 μL,H2O 14 μL,16 ℃反應過夜。將重組產物轉化至感受態細胞DH5α中,將菌液涂板37 ℃恒溫箱培養12 h,挑取單克隆菌落37 ℃搖菌16 h,酶切驗證后將陽性菌液送公司測序鑒定。對符合條件的質粒命名為shRNA-TGFβR1和shRNA-SMAD4質粒載體,對照組為shRNA-NC空白載體。

表2 shRNA oligo序列信息
1.2.4 過表達與干擾質粒載體轉染 顆粒細胞正常培養達到70%~80%融合度后,胰蛋白酶消化、PBS清洗,完全培養基重懸,以5×105個細胞鋪于6孔板中,每個組別3個重復。利用Lipofectamine 3000轉染試劑分別將pcDNA3.1(+)-TGFβR1、pcDNA3.1(+)-SMAD4、shRNA-TGFβR1和shRNA-SMAD4質粒轉染至綿羊顆粒細胞,參照lipo3000轉染說明書進行轉染,取兩個1.5 mL離心管,分別加入125 μL opti-MEM、5 μL lipo3000試劑和125 μL opti-MEM、2.5 μL質粒、5 μL lipo3000試劑,輕輕混勻,按1∶1輕輕混合兩管溶液于室溫下孵育10~15 min,將轉染溶液添加至6孔板中,在37 ℃培養48 h后收取細胞進行后續試驗。
1.2.5 細胞增殖、凋亡及周期的檢測 采用CCK-8細胞增殖檢測技術對顆粒細胞進行測定,將細胞接種于96孔板中培養24 h,在每孔中加入10 μL CCK-8溶液,置于培養箱中孵育2 h,用酶標儀測定450 nm處的OD值。采用細胞周期檢測試劑盒檢測顆粒細胞的細胞周期情況,收集處理好的細胞,加入500 μL PI/RNase A染色工作液,室溫混勻避光30 min,使用流式細胞儀檢測周期情況。采用Annexin-V FITC/PI試劑盒檢測顆粒細胞的凋亡情況,在懸浮細胞中加入10 μL Annexin-V FITC和5 μL PI溶液,混勻避光20 min,用流式細胞儀檢測細胞凋亡情況。
1.2.6TGFβR1和SMAD4基因表達水平的檢測 采用TRIzol法提取顆粒細胞的總RNA,并用cDNA鏈合成試劑盒得到cDNA,通過Real-Time PCR檢測轉染后顆粒細胞中TGFβR1和SMAD4的mRNA表達水平,采用Primer Premier 6.0 進行引物設計(表3)。配制20 μL反應體系:Master Mix 10.0 μL,上、下游引物各0.5 μL,cDNA 1.0 μL,ddH2O 8.0 μL。反應條件為:95 ℃ 1 min;95 ℃ 15 s,63 ℃ 25 s,40個循環。每個樣品重復3次,各個基因的相對表達水平用2-ΔΔct表示。

表3 Real-Time PCR的引物序列
1.2.7 蛋白水平的檢測 提取顆粒細胞的總蛋白,通過Western blot方法檢測TGFβR1、BAX、BCL2、Caspase3、SMAD4、p-TGFβR1、p-TGFβR2、p-SMAD2/3的蛋白水平。做SDS-PAGE電泳分析,電泳結束后進行蛋白質轉膜和轉印膜封閉后進行一抗雜交,4 ℃孵育過夜,T-TBS漂洗4次,每次5 min;二抗室溫孵育1 h,T-TBS漂洗5次,每次5 min;ECL工作液處理后放于成像儀中顯色留圖,采用Image J軟件對蛋白條帶進行分析。
1.2.8 統計分析 本試驗均重復3次,使用SPSS 26.0軟件進行統計學分析,結果均以“平均值±標準差”表示,采用單因素方差分析檢驗數據的差異顯著性,P<0.05表示具有顯著性差異,P<0.01表示具有極顯著性差異,采用Origin 2021軟件進行繪圖。
2 結 果
2.1 綿羊卵巢顆粒細胞的鑒定
綿羊卵巢顆粒細胞可特異性表達FSHR蛋白,檢測FSHR的存在狀況可鑒定顆粒細胞,如圖1所示,FSHR在細胞質中表達,經FSHR染色后,細胞的胞漿部分呈綠色熒光,而DAPI染色后細胞核清晰呈現藍色熒光,通過重疊可直觀表現出細胞的細胞核及細胞質的分布區域,顯微鏡下可見的FSHR表達率>90%,表明分離培養的綿羊顆粒細胞純度較高,符合后續試驗對顆粒細胞的條件要求。

圖1 綿羊顆粒細胞FSHR免疫熒光染色情況
2.2 TGFβR1過表達和干擾的轉染效率
通過Real-Time PCR及Western blot對轉染過表達和干擾TGFβR1載體的顆粒細胞中TGFβR1的表達量進行檢測,結果顯示(圖2A),pc3.1-TGFβR1過表達載體轉染顆粒細胞后,TGFβR1的表達量較對照組(PC組)極顯著升高(P<0.01),而shRNA-TGFβR1組轉染顆粒細胞后,TGFβR1的表達量較shRNA-NC組極顯著降低(P<0.01)。通過Western blot方法對轉染過表達或干擾TGFβR1載體的顆粒細胞中TGFβR1蛋白表達變化進行檢測,結果表明(圖2B),過表達TGFβR1會極顯著增加其蛋白的表達量(P<0.01),干擾TGFβR1會極顯著降低蛋白的表達量(P<0.01)。以上結果表明,TGFβR1過表達載體和干擾載體能夠顯著改變TGFβR1的表達水平,可用于后續試驗。

A.TGFβR1 mRNA相對表達量;B.TGFβR1蛋白相對表達量。*.P<0.05;**.P<0.01,下同。過表達組基因相對表達量均按PC=1的標準進行均一化處理;干擾組基因相對表達量均按shRNA-NC=1的標準進行均一化處理。下同
2.3 TGFβR1過表達和干擾對顆粒細胞增殖/周期/凋亡的影響
用CCK-8方法檢測轉染過表達和干擾TGFβR1載體的顆粒細胞增殖的情況,結果表明(圖3A),TGFβR1過表達促進顆粒細胞增殖,TGFβR1干擾較shRNA-NC對照組會抑制顆粒細胞的增殖。細胞周期檢測結果顯示(圖3B),TGFβR1過表達,G0/G1期細胞數量極顯著降低,對應S期細胞數量極顯著增加(P<0.01),G2/M期細胞數量無顯著變化(P>0.05);TGFβR1干擾可極顯著降低G0/G1和S期的細胞數量,而G2/M期細胞數量則極顯著增加(P<0.01)。通過Annexin-V FITC/PI檢測細胞凋亡結果顯示(圖3C),與PC組對比,TGFβR1過表達可以顯著降低顆粒細胞的凋亡數量(P<0.05);與shRNA-NC對照組相比,TGFβR1干擾則顯著增加顆粒細胞凋亡數量(P<0.05)。TGFβR1過表達和干擾會影響凋亡蛋白的表達(圖3D),TGFβR1過表達會極顯著抑制BAX和Caspase3蛋白的表達水平,TGFβR1干擾會極顯著抑制BCL2蛋白的表達水平(P<0.01)。

A.TGFβR1過表達/干擾后顆粒細胞CCK-8檢測結果;B.TGFβR1過表達/干擾處理后顆粒細胞周期流式檢測結果;C.TGFβR1過表達/干擾處理后顆粒細胞凋亡檢測結果(Q1.壞死;Q2.凋亡晚期;Q3.凋亡早期;Q4.正常);D.TGFβR1過表達/干擾處理凋亡蛋白BAX、BCL2、Caspase3表達水平檢測結果
2.4 SMAD4過表達和干擾的轉染效率
通過Real-Time PCR及Western blot對過表達和干擾SMAD4載體的顆粒細胞中SMAD4的表達量進行檢測,結果顯示(圖4A),pc3.1-SMAD4載體轉染顆粒細胞后,SMAD4的表達量較對照組(PC組)極顯著升高(P<0.01),而shRNA-SMAD4組轉染顆粒細胞后,SMAD4的表達量較shRNA-NC組極顯著降低(P<0.01)。通過Western blot方法對轉染過表達和干擾SMAD4載體的顆粒細胞中SMAD4的蛋白表達變化進行檢測,結果表明(圖4B),SMAD4過表達會極顯著增加其蛋白的表達量(P<0.01),干擾SMAD4極顯著降低其蛋白的表達量(P<0.01)。以上結果表明,SMAD4過表達載體和干擾載體能夠顯著改變SMAD4的表達水平,均可用于后續試驗。

A.SMAD4 mRNA相對表達量;B.SMAD4蛋白相對表達量
2.5 SMAD4過表達和干擾對顆粒細胞增殖/周期/凋亡的影響
通過CCK-8方法檢測轉染過表達和干擾SMAD4載體的顆粒細胞增殖的情況(圖5A),SMAD4過表達可極顯著促進顆粒細胞的增殖(P<0.01),干擾SMAD4可極顯著抑制顆粒細胞的增殖(P<0.01)。細胞周期檢測結果顯示(圖5B),SMAD4過表達,G0/G1期的細胞數量極顯著下降(P<0.01),G2/M期細胞數量顯著降低(P<0.05),而S期細胞數量則極顯著增加(P<0.01);干擾SMAD4后的顆粒細胞,G0/G1期的細胞數量極顯著下降,G2/M期和S期細胞數量極顯著增多(P<0.01)。細胞凋亡檢測結果顯示(圖5C),pc3.1-SMAD4載體轉染顆粒細胞后,降低了凋亡細胞比例,對細胞凋亡的抑制具有積極作用;shRNA-SMAD4組轉染顆粒細胞后與對照組(shRNA-NC組)相比,細胞凋亡數量有所增多。進一步探究SMAD4對凋亡蛋白表達水平的影響發現(圖5D),SMAD4過表達會極顯著抑制BAX和Caspase3蛋白的表達,干擾SMAD4會極顯著抑制BCL2蛋白的表達(P<0.01)。

A.SMAD4過表達/干擾后顆粒細胞CCK-8檢測結果;B.SMAD4過表達/干擾后顆粒細胞周期流式檢測結果;C.SMAD4過表達/干擾處理后顆粒細胞凋亡檢測結果(Q1.壞死;Q2.凋亡晚期;Q3.凋亡早期;Q4.正常);D.SMAD4過表達/干擾后凋亡蛋白BAX、BCL2、Caspase3的蛋白表達水平檢測結果
2.6 TGFβR1過表達和干擾對SMAD4蛋白表達的影響
TGFβR1過表達與干擾載體轉染顆粒細胞后發現(圖6),TGFβR1過表達會極顯著促進SMAD4蛋白的表達(P<0.01),干擾TGFβR1后極顯著降低SMAD4蛋白的表達(P<0.01),表明TGFβR1對SMAD4蛋白的表達具有促進作用。

圖6 TGFβR1過表達和干擾對SMAD4蛋白表達的影響
2.7 SMAD4過表達和干擾對TGFβR1蛋白表達的影響
SMAD4過表達和干擾載體轉染顆粒細胞后,檢測TGFβR1蛋白的表達情況發現(圖7),SMAD4過表達會極顯著增加TGFβR1蛋白的表達(P<0.01),干擾SMAD4極顯著降低TGFβR1蛋白的表達(P<0.01)。
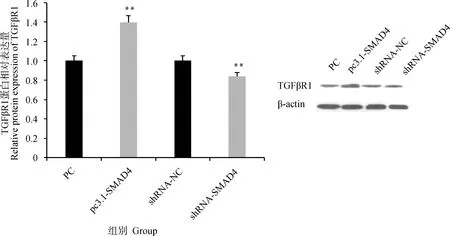
圖7 SMAD4過表達和干擾對TGFβR1蛋白表達的影響
2.8 TGFβR1過表達和干擾對TGF-β信號通路蛋白表達的影響
TGFβR1過表達和干擾載體轉染顆粒細胞后,檢測TGF-β信號通路蛋白(p-TGFβR1、p-TGFβR2、p-SMAD2/3)的表達情況發現(圖8),TGFβR1過表達極顯著促進了TGFβR1、TGFβR2及SMAD2/3蛋白的磷酸化水平(P<0.01),與此同時,TGFβR1干擾會極顯著抑制p-TGFβR1與p-TGFβR2的表達(P<0.01),會降低SMAD2/3的磷酸化水平。

A.TGFβR1過表達后p-TGFβR1、p-TGFβR2、p-SMAD2/3蛋白的相對表達量;B.TGFβR1干擾后p-TGFβR1、p-TGFβR2、p-SMAD2/3蛋白的相對表達量
3 討 論
綿羊胎產羔數的提升是近年來國內外研究的熱點之一。綿羊卵泡發育影響繁殖力的高低,許多研究表明,顆粒細胞的發育狀態與卵泡的命運有關。顆粒細胞增殖促進卵泡成熟和排卵,而凋亡導致卵泡閉鎖和退化[24],抑制卵巢顆粒細胞凋亡,可促進卵泡發育,提高排卵率和胎產羔數。TGF-β/Smad信號通路在細胞增殖分化、個體發育、軟骨發生以及生殖發育、組織創傷修復等過程中都發揮著極其重要的作用[25-26]。本試驗探討了TGFβR1對綿羊顆粒細胞增殖凋亡的作用,并進一步探究TGFβR1如何通過TGF-β/Smad信號通路參與顆粒細胞功能的調控。結果發現,過表達TGFβR1可加快細胞進程進入S期,促進綿羊顆粒細胞增殖,抑制顆粒細胞凋亡,干擾TGFβR1會發生G2/M期阻滯,抑制綿羊顆粒細胞的增殖,促進其凋亡。這些發現與前人的研究結果類似,TGFβR1是TGFβ配體的1型受體[27-28],參與下丘腦-垂體-卵巢軸的調控,促進卵子的發育成熟及黃體形成,參與滋養細胞的增殖、分化[29];TGFβR1還可以介導生長分化因子9(GDF9)的信號,促進顆粒細胞的增殖和人腔前卵泡的生長[30];敲除TGFβR1基因的小鼠能排卵和產生卵母細胞,但會發生胚胎死亡[31],同時還伴隨著嚴重的輸卵管畸形[28,32],這些結果顯示,TGFβR1會對顆粒細胞的增殖產生促進作用。進一步研究發現,TGFβR1會抑制促凋亡基因BAX和Caspase3的表達,促進抗凋亡基因BCL2的表達。有研究表明,抗凋亡蛋白BCL2和促凋亡蛋白BAX是細胞凋亡的關鍵調節因子,其主要作用部位是線粒體膜:前者抑制細胞色素c(Cytc)的釋放,后者啟動Cytc的釋放。BCL2蛋白通過調節生殖細胞和體細胞的凋亡,在卵泡生長發育和卵泡閉鎖中發揮重要作用。BCL2基因缺陷的小鼠表現出卵母細胞和原始卵泡數量的減少[33],而BCL2的過表達減少了大的有腔卵泡的顆粒細胞凋亡,從而增加卵泡發生,提高排卵率。而BAX在卵母細胞和顆粒細胞中都有表達,缺失BAX基因的小鼠表現出過多的異常卵泡[34]。與正常卵泡相比,閉鎖卵泡顆粒細胞中BAX的表達較強[35]。在豬顆粒細胞中,閉鎖卵泡BAX的表達高于健康卵泡[24,36]。Caspase家族在介導細胞凋亡的過程中發揮重要作用[37],Caspase3是參與多種凋亡信號轉導途徑的關鍵執行分子[38-39],活化的Caspase3會裂解相應的胞漿胞核底物,最終導致細胞凋亡[40]。這些結果表明,TGFβR1通過調控BCL2、BAX和Caspase3基因的表達,參與顆粒細胞的增殖凋亡進程。干擾TGFβR1會抑制TGFβR1、TGFβR2以及SMAD2/3的磷酸化,表明TGF-β信號通路被阻斷。SMAD2/3是轉化生長因子-β信號轉導的核心成分,是轉化生長因子-β的信號中間產物,激活的TGFβR1通過磷酸化SMAD2/3(R-Smads)蛋白激活Smad信號通路,磷酸化的R-Smads共同與SMAD4蛋白結合,將信號由胞漿轉移至細胞核內,調節靶基因的轉錄,誘導下游目的基因的表達[11,41]。
由于在TGF-β/Smad信號通路中,TGF-β二聚體結合TGFβR2,并導致TGFβR2自身磷酸化激活,同時將TGFβR1募集到高度保守的近膜區域,之后TGFβR2磷酸化激活TGFβR1并和其之間形成異四聚體復合物,磷酸化的TGFβR1將依次招募SMAD2和SMAD3與其結合將其磷酸化后轉為活化狀態,將信號傳遞到胞漿中,SMAD2/3與SMAD4形成異構體復合物,最終將信號由胞漿傳遞至細胞核中,在細胞核中調節下游基因的表達。SMAD4在該通路中發揮著關鍵作用,故進一步探究了TGF-β/Smad通路中TGFβR1下游的關鍵調節因子SMAD4與顆粒細胞功能的關系,研究發現,TGF-β/Smad通路的關鍵調節因子SMAD4與顆粒細胞的增殖凋亡有密切關系,過表達SMAD4會加快細胞進程進入至S期,促進顆粒細胞的增殖,抑制促凋亡蛋白BAX和Caspase3的表達,促進抗凋亡蛋白BCL2的表達;抑制SMAD4則發生G2/M期阻滯,抑制顆粒細胞增殖,抑制BCL2的表達,促進BAX和Caspase3的表達。SMAD4作為一種常見的TGF-β信號轉導介質,被認為在調節卵泡生長和雌性生殖方面發揮了重要作用[9]。敲除卵巢特異性基因SMAD4的小鼠出現生育能力下降,生殖力隨著時間的推移而下降,卵泡發生中存在多種缺陷,如嚴重的卵丘細胞缺陷和顆粒細胞過早黃體化[42]。已有研究發現,腔前顆粒細胞中SMAD4基因的缺失會導致后續卵泡發育和分化的嚴重缺陷[9]。以上發現證實,TGFβR1介導的TGF-β/Smad通路可以調控綿羊顆粒細胞功能,當SMAD4基因過表達即該信號通路信號傳遞增強時,可促進綿羊顆粒細胞的增殖,并抑制其凋亡。
4 結 論
TGFβR1促進綿羊顆粒細胞的增殖及周期進程,抑制顆粒細胞凋亡,通過介導TGF-β/Smad信號通路中SMAD2/3及SMAD4的表達正向調控綿羊顆粒細胞的增殖與周期,并負向調控其凋亡。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
鴨綠江(2021年35期)2021-04-19 12:24:18
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
海峽科技與產業(2016年3期)2016-05-17 04:32:12
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
