牦牛Fas相關(guān)因子1多克隆抗體制備及初步應(yīng)用
2023-08-25 08:08:48王靜瑜潘陽陽徐庚全張文蘭王筱珊烏仁套迪照日格圖余四九
畜牧獸醫(yī)學報 2023年8期
王靜瑜,潘陽陽,徐庚全,張 瑞,張文蘭,王筱珊,烏仁套迪,照日格圖,崔 燕,余四九*
(1.甘肅農(nóng)業(yè)大學動物醫(yī)學院/甘肅省牛羊胚胎工程技術(shù)研究中心,蘭州 730070;2.阿拉善盟畜牧研究所,阿拉善 750300)
FAF1是1995年由Chu等[1]以酵母雙雜交方法篩選得到,FAF1蛋白是死亡誘導(dǎo)信號復(fù)合物(death inducing signaling complex,DISC)的組成部分。FAF1蛋白為胞內(nèi)蛋白[2],主要表達于不同細胞類型的細胞核及核周胞質(zhì),其在進化上較為保守,由650個氨基酸構(gòu)成,大小約為 74 ku,含有多個功能性結(jié)構(gòu)域,主要包括N末端泛素相關(guān)域(ubiquitin-associating,UBA)、C端泛素調(diào)節(jié)X域(ubiquitin regulatory X,UBX)、死亡效應(yīng)結(jié)構(gòu)域(death effector domain interacting domain,DEDID)、Fas相互作用域(FAS-interacting domain,FID)[3]。FAF1 mRNA及蛋白質(zhì)在人和小鼠的睪丸及卵巢中均有表達[4]。但是FAF1在牦牛睪丸和卵巢中的生物學作用還尚不清楚。
牦牛(Bosgrunniens)為青藏高原一帶獨有的原始牛種之一,一般分布于海拔3 000~5 000米的高原上,它不但能夠適應(yīng)高原寒冷、空氣稀薄、強輻射的地理環(huán)境,也可用于農(nóng)牧業(yè)和交通運輸。除此之外,還能為當?shù)啬撩裉峁┤狻⒛獭⒚⑵じ铩⑷剂系壬a(chǎn)和生活資料,被譽為藏區(qū)“牧民之寶”。素有“高原之舟”和“冰河之車”的美稱[5]。FAF1廣泛存在于多種組織和細胞中,含有多種功能結(jié)構(gòu)域,參與不同信號通路的調(diào)節(jié),具有多種重要的生殖生理功能。由于敲除FAF1基因的純合子會在胚胎早期發(fā)育死亡[6],限制了通過基因敲除技術(shù)探索FAF1基因的生物學功能,因此通過制備牦牛FAF1多克隆抗體來探索FAF1基因生物學功能具有重要的意義。高品質(zhì)的精液是保證受精和早期胚胎發(fā)育的重要條件,較好的卵巢機能對于受精具有重要意義[7]。細胞凋亡是生殖調(diào)節(jié)的機制之一,生殖細胞(精子和卵母細胞)的周期性變化與其凋亡過程密切相關(guān)[8],Fas-FasL系統(tǒng)被認為是生殖細胞(精子和卵子)凋亡的關(guān)鍵生理調(diào)節(jié)因子[9]。FAF1是Fas-FasL凋亡通路重要調(diào)控元件[10],FAF1通過多種方式作用于細胞凋亡[10],并且其在生殖系統(tǒng)中表達量較高。本研究以牦牛睪丸組織cDNA為模板,擴增FAF1基因全序列,構(gòu)建全長 pET-28a-FAF1原核表達載體,優(yōu)化誘導(dǎo)表達條件,經(jīng)鎳柱純化并鑒定后免疫試驗動物,獲得兔源FAF1多克隆抗體,以牦牛不同年齡睪丸組織及處于不同繁殖周期的卵巢為研究對象,采用實時熒光定量檢測不同年齡睪丸組織及處于不同繁殖周期的卵巢FAF1 mRNA相對表達水平,采用Western blot檢測不同年齡睪丸組織及處于不同繁殖周期的卵巢FAF1蛋白相對表達水平,采用免疫組織化學技術(shù)定位FAF1蛋白表達部位,以期為進一步研究FAF1在牦牛生殖方面的功能提供依據(jù)。
1 材料與方法
1.1 試驗材料
本研究所使用的試劑主要有:TransZol(北京,全式金);GoScript Reverse Transcription System (美國,Promega);Goat anti-rabbit IgG-HRP(北京,博奧森);DNA純化回收試劑盒、質(zhì)粒小提試劑盒、DH5a 感受態(tài)細胞、BL21(DE3)/Rosetta感受態(tài)細胞(北京,天根);IPTG、卡那(北京,索萊寶);PVDF膜(美國,Millipore);酵母、氯化粉鈉、瓊脂、胰蛋白胨(英國,Oxoid);pET-28a為實驗室保存;限制性核酸內(nèi)切酶XhoⅠ、限制性核酸內(nèi)切酶EcoR I、T4連接酶(大連,TaKaRa);Ni-NTA-Sefinose Column(上海,生工);預(yù)混型 qPCR 試劑盒(湖南,艾科瑞生物);FAF1兔抗牦牛多克隆抗體為甘肅省牛羊胚胎工程中心制備保存。
本研究所使用的儀器主要有:PCR儀(美國,Bio-Rad);SDS-PAGE電泳槽(美國,Rio-Rad公司);超聲波破碎儀(寧波,新芝);恒溫培養(yǎng)箱(日本,松下);恒溫振蕩搖床(上海,恒字);超靈敏多功能成像儀(美國,GE Healthcare)。
1.2 樣品的采集
試驗樣品于2021年10月采集于青海省西寧市屠宰場。挑選不同年齡的健康雄性牦牛各3頭,挑選年齡相仿且處于不同繁殖周期(卵泡期卵巢上具有成熟卵泡、黃體期卵巢有明顯黃體和妊娠期卵巢在子宮內(nèi)有發(fā)育的胎兒的卵巢)的健康雌性牦牛各3頭,經(jīng)頸動脈放血致死后,迅速采集睪丸組織和卵巢組織,用0.9%的生理鹽水沖洗3次,將組織修剪為約1 cm3的組織塊并浸泡于4%的中性多聚甲醛溶液中固定,以便于開展后期的免疫組織化學試驗。剩余組織用錫箔紙包裹后放入凍存袋內(nèi),然后放入液氮罐中,過程中盡可能的快速,以防止組織降解變質(zhì),運回實驗室后轉(zhuǎn)存于-80 ℃超低溫冰箱中,以便于開展后期的分子試驗。
1.3 目的基因的獲取
利用GoScriptTMReverse Transcription System試劑盒將總RNA反轉(zhuǎn)錄為cDNA。以β-actin(肌動蛋白)為內(nèi)參基因,β-actin及FAF1基因引物序列如表1。Taq PCR Master Mix 10 μL,ddH2O 8 μL,模板cDNA 1 μL,β-actin-F、β-actin-R各0.5 μL,共20 μL體系置于PCR儀中,95 ℃預(yù)變性5 min,95 ℃變性30 s,60 ℃退火30 s,72 ℃延伸8 s,72 ℃條件下10 min,循環(huán)40次。反應(yīng)結(jié)束后瓊脂糖凝膠電泳進行驗證。模板cDNA驗證成功后,擴增FAF1基因,體系共20 μL:Taq PCR Master Mix 10 μL,ddH2O 8 μL,模板cDNA 1 μL,FAF1-F、FAF1-R各 0.5 μL,退火時間變?yōu)? min,其它條件不變。瓊脂糖凝膠電泳后,利用DNA純化回收試劑盒純化目的基因。
1.4 原核載體pET-28a-FAF1的構(gòu)建
純化的DNA及空載體pET-28a使用限制性內(nèi)切酶XhoⅠ和EcoR Ⅰ進行雙酶切。將酶切好的目的基因與酶切好的載體使用T4 DNA連接酶16 ℃條件過夜連接。將重組質(zhì)粒轉(zhuǎn)化至DH5a 感受態(tài)細胞中,涂布至含卡那的LB培養(yǎng)基上,37 ℃過夜培養(yǎng)。挑取陽性克隆菌,提取質(zhì)粒,分別進行PCR及雙酶切驗證,驗證成功后送至上海生工測序。將連接成功的重組質(zhì)粒轉(zhuǎn)化至BL21(DE3)/Rosetta感受態(tài)細胞。
1.5 重組蛋白優(yōu)化誘導(dǎo)
挑取含有于pET-28a重組子的BL21(DE3)/Rosetta菌落接種于5 mL含卡那的LB培養(yǎng)基中,37 ℃振蕩培養(yǎng)過夜。取150 μL培養(yǎng)液于5 mL含卡那的LB培養(yǎng)基中,37 ℃條件下200 r·min-1振蕩培養(yǎng)3 h。取1 mL作為IPTG誘導(dǎo)前的樣品,12 000 r·min-1離心1 min,棄上清液,菌體沉淀用40 μL PBS懸浮,-20 ℃保存。其余樣品加入IPTG,使IPTG終濃度為0.5 mmol·L-1,分別在20 ℃、25 ℃、28 ℃、32 ℃、37 ℃條件下180 r·min-1振蕩培養(yǎng)。分別在各溫度誘導(dǎo)后2、3、4、5 h取1 mL樣品的菌液,12 000 r·min-1離心1 min,收集菌體,用30 μL PBS懸浮,-20 ℃保存。在誘導(dǎo)前以及誘導(dǎo)后各時間的樣品中各加入10 μL 4×SDS上樣緩沖液,渦旋振蕩1 min,使菌體完全溶菌,沸水浴10 min。在4 ℃條件下12 000 r·min-1離心1 min后取上清到新的離心管中。分別取20 μL誘導(dǎo)前以及10 μL誘導(dǎo)后的樣品進行SDS-PAGE分析。為提高FAF1重組蛋白的表達量,對誘導(dǎo)劑IPTG濃度(0.1、0.3、0.5、0.7、1.0 mmol·L-1)進行優(yōu)化,同時以未誘導(dǎo)樣品作為對照,取不同濃度的樣品經(jīng)SDS-PAGE分離并用考馬斯亮藍染色后進行分析。
1.6 重組蛋白大量誘導(dǎo)表達及純化
根據(jù)最佳誘導(dǎo)條件進行陽性克隆菌的大量培養(yǎng),收集菌體,加入PBS緩沖液重懸,冰浴條件下,超聲波破碎,功率40 W,工作3 s,間隔3 s,總時間約30 min,以菌體由渾濁變透明且不黏稠為準。將破碎的菌液于4 ℃條件下5 000 r·min-1離心10 min,收集上清和沉淀,將所得上清和沉淀變性,SDS-PAGE電泳考馬斯亮藍染色。按照Ni-NTA-Sefinose Column說明書對重組蛋白純化。
1.7 試驗動物免疫
選取2只健康雄性新西蘭大白兔,飼養(yǎng)1周觀察是否有應(yīng)激等不良反應(yīng),若無不良反應(yīng),則分別取耳緣靜脈血1 mL,留作陰性對照。以1.2 mg純化的FAF1重組蛋白與弗氏完全佐劑等量混勻并充分乳化,分多點背部皮下注射免疫2只新西蘭大白兔;第一次免疫兩周后進行第二次免疫,以1.2 mg純化的FAF1重組蛋白與弗氏不完全佐劑等量混勻并充分乳化,分多點肩胛部及腘淋巴結(jié)注射免疫2只新西蘭大白兔;第二次免疫3周后進行第三次免疫,以0.4 mg純化的FAF1重組蛋白與弗氏不完全佐劑等量混勻并充分乳化,分多點肩胛部及腘淋巴結(jié)注射免疫2只新西蘭大白兔;第三次免疫兩周后進行耳緣采血檢測效價,效價良好則可進行心臟采血收集血液,37 ℃恒溫箱靜置1 h,再于4 ℃冰箱靜置過夜,分離血清,至于-80 ℃超低溫冰箱保存?zhèn)溆谩?/p>
1.8 牦牛FAF1多克隆抗體效價檢測
將FAF1重組蛋白用包被液稀釋至1 μg·mL-1,96孔酶標板每孔加入100 μL,設(shè)置陰性對照和空白對照,前者加入含重組蛋白的包被液,后者只加入包被液,4 ℃過夜;使用1×PBST洗滌3次,拍干;加入5%脫脂奶粉封閉液37 ℃,3 h進行封閉;使用1×PBST洗滌3次,拍干;加入等比稀釋的兔多抗血清100 μL,陰性對照孔加入陰性血清,37 ℃孵育2 h;洗滌3次,拍干;每孔加入100 μL 以1∶10 000稀釋的HRP標記山羊抗兔二抗,37 ℃孵育2 h;洗滌3次,拍干;每孔加入100 μL TMB,37 ℃顯色15 min;每孔加入50 μL濃度為 2 mol·L-1的H2SO4終止顯色反應(yīng),放入酶標儀檢測450 nm處OD值。
1.9 多克隆抗體特異性檢測
將純化后的重組蛋白上樣進行SDS-PAGE電泳,65 V條件35 min,調(diào)至110 V 1 h。110V條件下轉(zhuǎn)膜70 min,5%脫脂奶粉封閉過夜,加制備的兔源多克隆抗體室溫孵育3 h,PBST洗滌5次,每次10 min,加二抗(Goat anti-rabbit IgG-HRP)室溫孵育40 min,PBST洗滌6次,每次10 min。在PVDF膜上滴加ELC超敏發(fā)光液,置于超靈敏多功能成像儀曝光成像。
1.10 實時熒光定量PCR
制備不同組織cDNA工作液,使終濃度為100 ng·μL-1。加入2X SYBR?Green Pro Taq HS Premix 10 μL,不同組織cDNA工作液1 μL,FAF1-F1、FAF1-R1各0.4 μL,RNase free water 8.2 μL,反應(yīng)體系共20 μL。反應(yīng)條件:95 ℃預(yù)變性300 s,95 ℃變性10 s,58 ℃退火30 s,72 ℃延伸16 s,共進行40個循環(huán)。每個模板,設(shè)4個重復(fù)組,使用 2-ΔΔCt法計算不同年齡睪丸組織及不同繁殖周期卵巢組織FAF1 mRNA的相對表達量。
1.11 Western blot
根據(jù)SDS-PAGE 分離膠和濃縮膠配制體系分別配制濃度為8%的分離膠和5%的濃縮膠。將變性好的蛋白樣品進行上樣電泳,待電泳結(jié)束后,使用濕轉(zhuǎn)的方式將蛋白轉(zhuǎn)印至0.45 μm的PVDF膜上,轉(zhuǎn)膜完成后用PBST 洗去膜上的殘留轉(zhuǎn)膜液,使用5%脫脂奶粉搖床上孵育過夜,PBST洗滌2次,每次30 min,一抗(兔抗牦牛FAF1多克隆抗體)按1∶1 000稀釋,室溫孵育2.5 h,PBST 洗滌 5 min,共洗滌6次,二抗(Goat Anti-Rabbit IgG-HRP)1∶8 000稀釋搖床上孵育 40 min,PBST 洗滌3次,每次20 min。將超敏化學發(fā)光 A 液與超敏化學發(fā)光B液按1∶1混勻配置成工作液,將工作液滴加至PVDF膜上并避光作用5 s,使用Amersham Imager 600 System進行化學發(fā)光檢測。根據(jù)成像結(jié)果,利用目的灰度值/內(nèi)參灰度值分析FAF1蛋白相對表達量。
1.12 免疫組織化學
將固定于4%中性多聚甲醛的組織在自來水中沖洗24 h,再經(jīng)梯度酒精脫水后使用酒苯透明,然后浸蠟,最后進行石蠟包埋。利用切片機將組織切為4 μm的連續(xù)性切片。在烘片機上60 ℃烘烤6 h,下行脫蠟水化,采用檸檬酸鹽進行抗原修復(fù),將載玻片放入0.01 mol·L-1的檸檬酸鹽中,高火加熱直至煮沸,再使用中火維持10 min,待冷卻至室溫后,PBS洗滌3 次,每次3 min;使用3% H2O2溶液于37 ℃條件下孵育10 min進行阻斷,PBS洗滌3次,每次3 min;山羊血清工作液室溫封閉(SPA 試劑盒A液)15 min,試驗組一抗(兔抗牦牛FAF1多克隆抗體)按1∶400稀釋后置于濕盒4 ℃孵育過夜,陰性對照組使用PBS作為一抗孵育過夜,PBS洗滌3 次,每次3 min;生物素標記山羊抗兔IgG(SPA試劑盒B液)37 ℃條件下孵育15 min,PBS洗滌3 次,每次3 min;辣根酶標記鏈霉卵白素工作液(SPA試劑盒C液)室溫孵育15 min,PBS洗滌3次,每次3 min;使用DAB顯色液進行顯色,經(jīng)蘇木精復(fù)染后鹽酸酒精分化,在自來水中返藍10 min。脫水透明后封片。待晾干后置于顯微鏡下拍照。
1.13 數(shù)據(jù)分析
使用 SPSS 25.0 對 FAF1 mRNA相對表達量和蛋白相對表達量進行單因素方差分析,每組3次重復(fù),計算結(jié)果均以“平均值±標準誤”來表示,P<0.05表示二者差異顯著,使用Graphpad prism 8 進行繪圖。
2 結(jié) 果
2.1 目的基因的獲取
以牦牛cDNA為模板,以FAF1-F、FAF1-R進行PCR擴增,其結(jié)果如圖1所示,在1 953 bp處有清晰單一的條帶,與預(yù)期大小一致。

M.2000bp ladder; 1.牦牛睪丸組織cDNA為模板
2.2 重組質(zhì)粒pET-28a-FAF1的鑒定
以重組質(zhì)粒pET-28a-FAF1為模板,FAF1-F、FAF1-R為引物進行PCR,結(jié)果如圖2A所示,在1 953 bp處有清晰明亮的條帶,產(chǎn)物大小與預(yù)期相符。將重組質(zhì)粒pET-28a-FAF1雙酶切后,結(jié)果如圖2B所示,在5 335bp和1 953bp處有較清晰的條帶,且與預(yù)期大小一致。重組質(zhì)粒pET-28a-FAF1經(jīng)上海生工測序確認pET-28a-FAF1原核表達載體構(gòu)建成功。
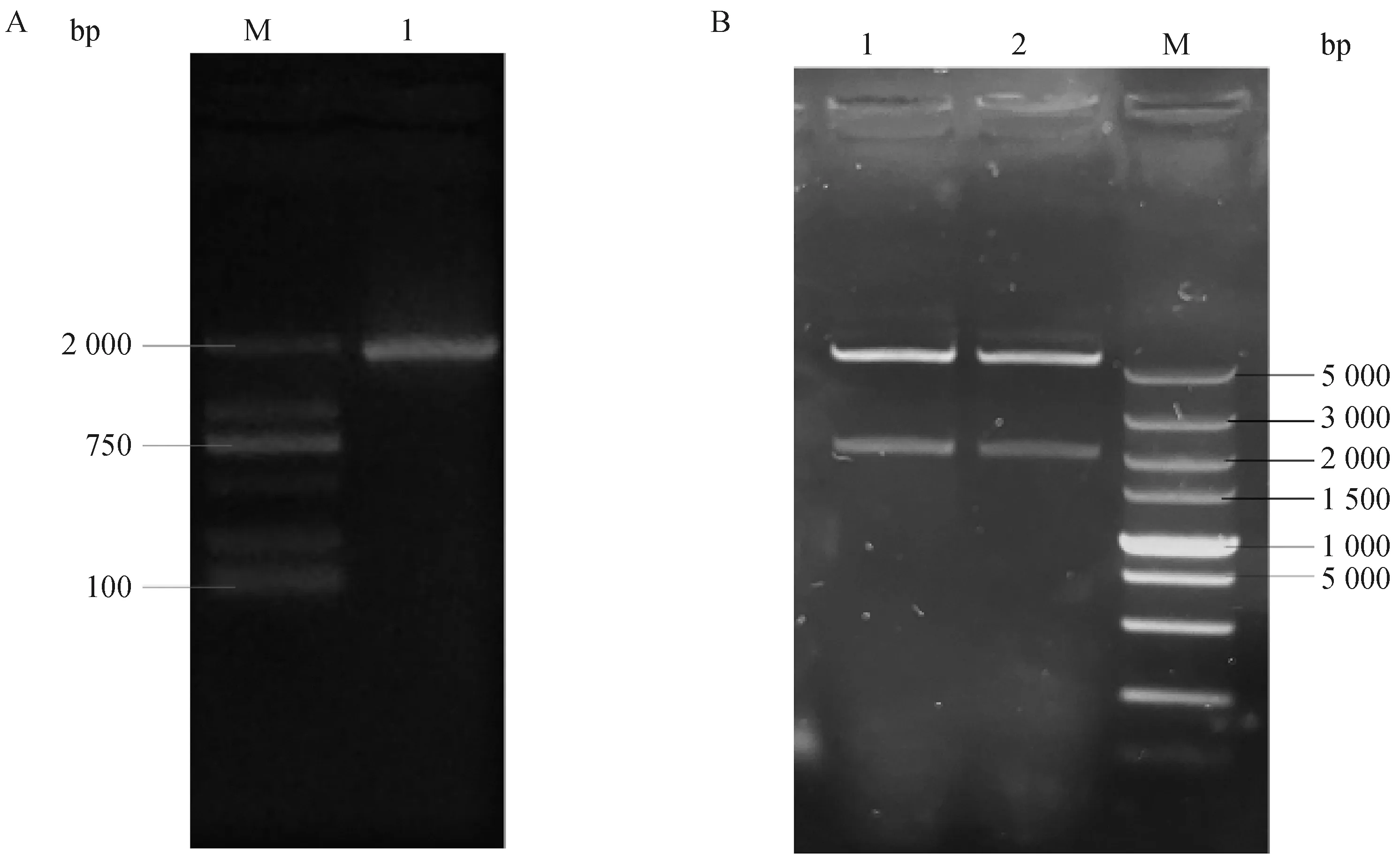
A.重組質(zhì)粒pET-28a-FAF1 PCR:M.2000 bp ladder;1.構(gòu)建重組質(zhì)粒。B.重組質(zhì)粒pET-28a-FAF1雙酶切:M.5000 bp ladder;1~2.不同菌落重組質(zhì)粒
2.3 重組蛋白的優(yōu)化誘導(dǎo)
重組蛋白pET-28a-FAF1在20 ℃(圖3A)、25 ℃(圖3B)、28 ℃(圖3C)、32 ℃(圖3D)條件下,隨誘導(dǎo)時間增加,表達量呈現(xiàn)增加的趨勢,37 ℃(圖3E)條件下溫度過高,導(dǎo)致該重組蛋白表達量降低。該重組蛋白大小與預(yù)期一致,為74 ku,其在25 ℃表達量最大,該溫度為該重組蛋白的最優(yōu)誘導(dǎo)溫度;25 ℃下誘導(dǎo)5 h表達量最高,認為5 h為最佳誘導(dǎo)時間。不同濃度IPTG對FAF1重組蛋白誘導(dǎo)表達(圖4)并無太大影響,因此選擇0.5 mmol·L-1作為IPTG最佳誘導(dǎo)濃度。

A.20 ℃不同誘導(dǎo)時間;B.25 ℃不同誘導(dǎo)時間;C.28 ℃不同誘導(dǎo)時間;D.32 ℃不同誘導(dǎo)時間;E.37 ℃不同誘導(dǎo)時間。M.蛋白預(yù)染Marker;1.未誘導(dǎo);2.誘導(dǎo)2 h;3.誘導(dǎo)3 h;4.誘導(dǎo)4 h;5.誘導(dǎo)5 h

M.蛋白預(yù)染Marker;1~6.IPTG濃度分別為0.1、0.3、0.5、0.7、1.0和0 mmol·L-1
2.4 重組蛋白的表達和純化
將破碎后的上清和沉淀變性,經(jīng)SDS-PAGE電泳,考馬斯亮藍染色后,如圖5A所示,經(jīng)破碎后重組蛋白pET-28a-FAF1基本全部存在于沉淀中,該重組蛋白為不可溶性蛋白。重組蛋白pET-28a-FAF1經(jīng)鎳柱純化后結(jié)果如圖5B所示,其大小為74 ku,與預(yù)期一致,成功獲得較高純度的重組蛋白pET-28a-FAF1。
2.5 牦牛FAF1多克隆抗體效價檢測
ELISA法檢測多抗血清抗體效價結(jié)果(圖6)分析顯示,稀釋度為1∶1 024 000時的陽性血清OD450 nm>2.1倍陰性血清OD450 nm陰性血清,因此,兔源FAF1多克隆抗體效價為1∶1 024 000。
2.6 牦牛FAF1多克隆抗特異性檢驗
以重組FAF1蛋白為抗原,1∶2 000稀釋兔抗血清作為一抗,Goat anti-rabbit IgG-HRP 以1∶8 000稀釋,進行Western blot檢測,結(jié)果(圖7)顯示在74 ku處出現(xiàn)特異性條帶。結(jié)果表明,本研究制備的兔抗FAF1多克隆抗體可與牦牛重組FAF1蛋白能夠發(fā)生特異性反應(yīng)。
2.7 FAF1 mRNA在不同年齡睪丸組織相對表達量
qRT-PCR 結(jié)果統(tǒng)計分析(圖8)表明FAF1 mRNA的表達普遍存在于不同年齡睪丸組織中。其中3、4、7周歲雄性牦牛睪丸組織中FAF1mRNA相對表達量顯著高于其他年齡段。FAF1 mRNA的相對表達水平在牦牛不同年齡睪丸組織中存在差異性表達。

柱上所標字母相異表示差異顯著(P<0.05),所標字母相同表示差異不顯著(P>0.05)。下同
2.8 FAF1 蛋白在不同年齡睪丸組織相對表達量
Western blot結(jié)果(圖9)顯示,FAF1蛋白普遍存在于雄性牦牛不同年齡睪丸組織。統(tǒng)計分析后結(jié)果顯示(圖 10),11周歲時FAF1相對蛋白表達量顯著高于其他年齡,6、7周歲次之,3、4周歲FAF1相對蛋白表達量最低,FAF1 蛋白在牦牛不同年齡的相對表達量存在差異性。
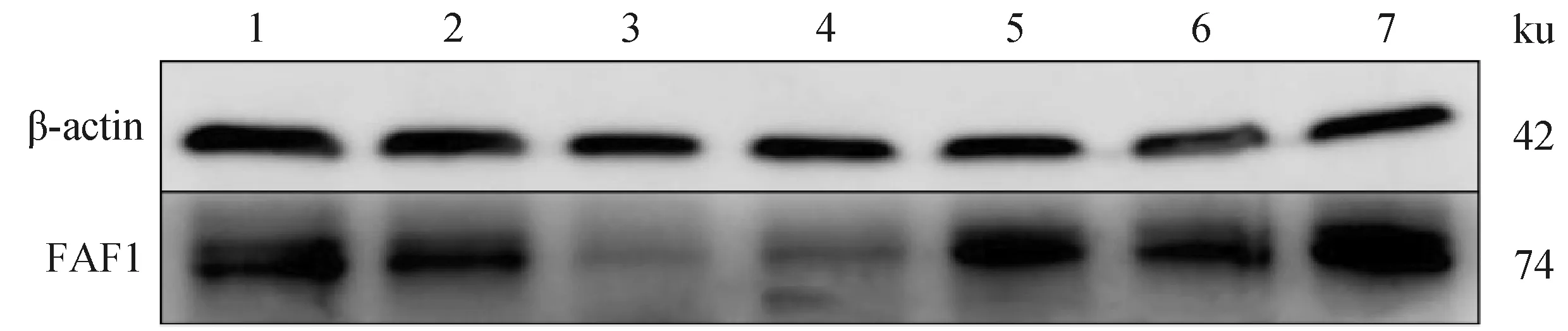
1~7.1、2、3、4、6、7、11周歲牦牛睪丸樣品
2.9 FAF1 蛋白在不同年齡睪丸組織的表達定位
免疫組織化學染色結(jié)果(圖11)顯示,FAF1蛋白在不同年齡段牦牛睪丸中均有棕褐色陽性表達,不同年齡睪丸FAF1蛋白分布表達部位基本一致,主要表達于精子、精子細胞、精原細胞、初級精母細胞、支持細胞、睪丸間質(zhì)細胞和管周肌樣細胞。
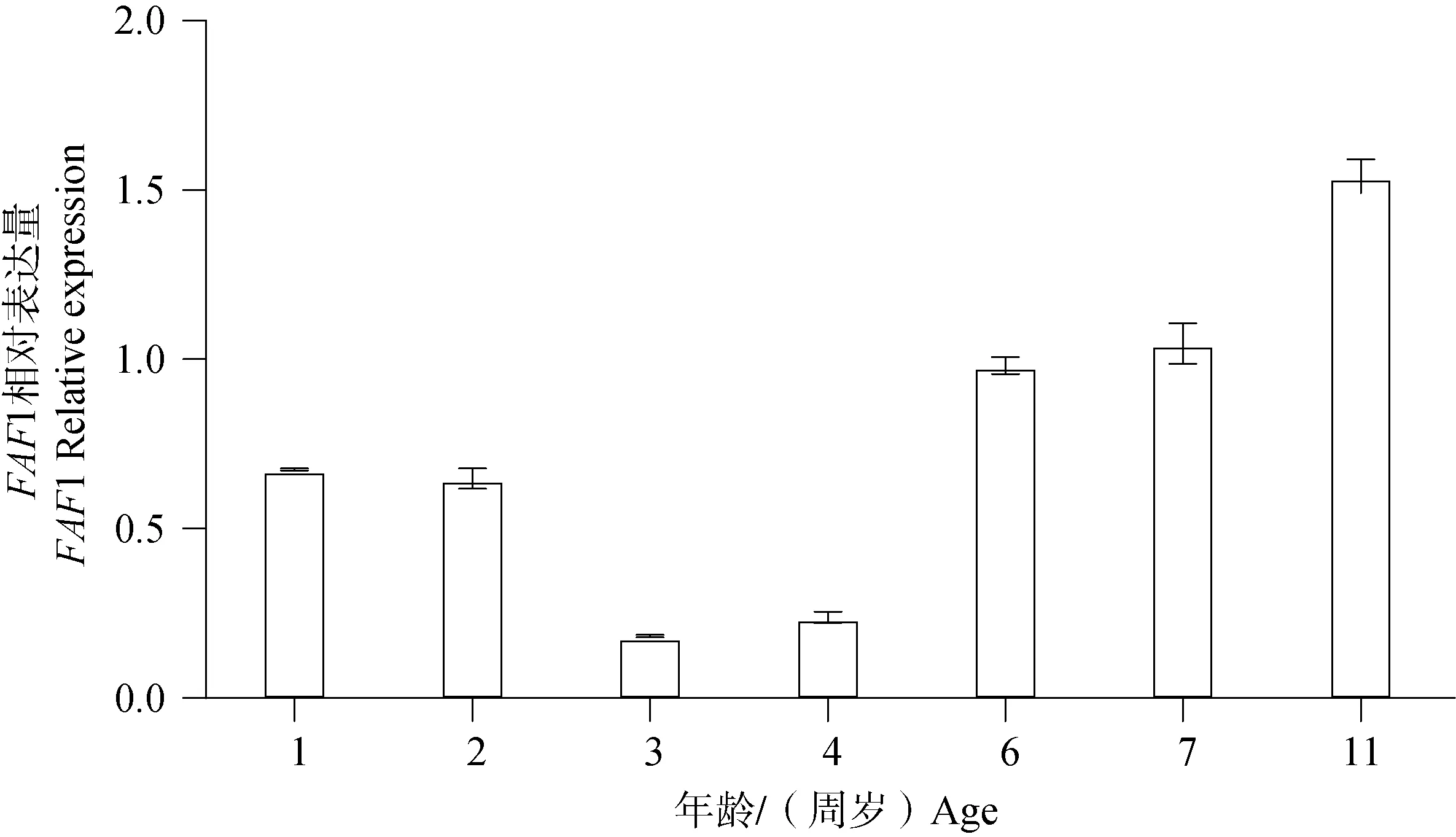
圖10 FAF1蛋白在不同年齡雄性牦牛睪丸組織中的相對表達水平
2.10 FAF1 基因在不同繁殖周期卵巢的相對表達水平
qRT-PCR結(jié)果(圖12)統(tǒng)計分析表明FAF1 mRNA普遍表達存于不同時期的卵巢。黃體期卵巢FAF1 mRNA相對表達量顯著高于卵泡期和妊娠期,妊娠期卵巢次之,卵泡期卵巢FAF1 mRNA相對表達量最低。FAF1 mRNA的相對表達量在牦牛不同繁殖周期卵巢組織存在差異化表達。
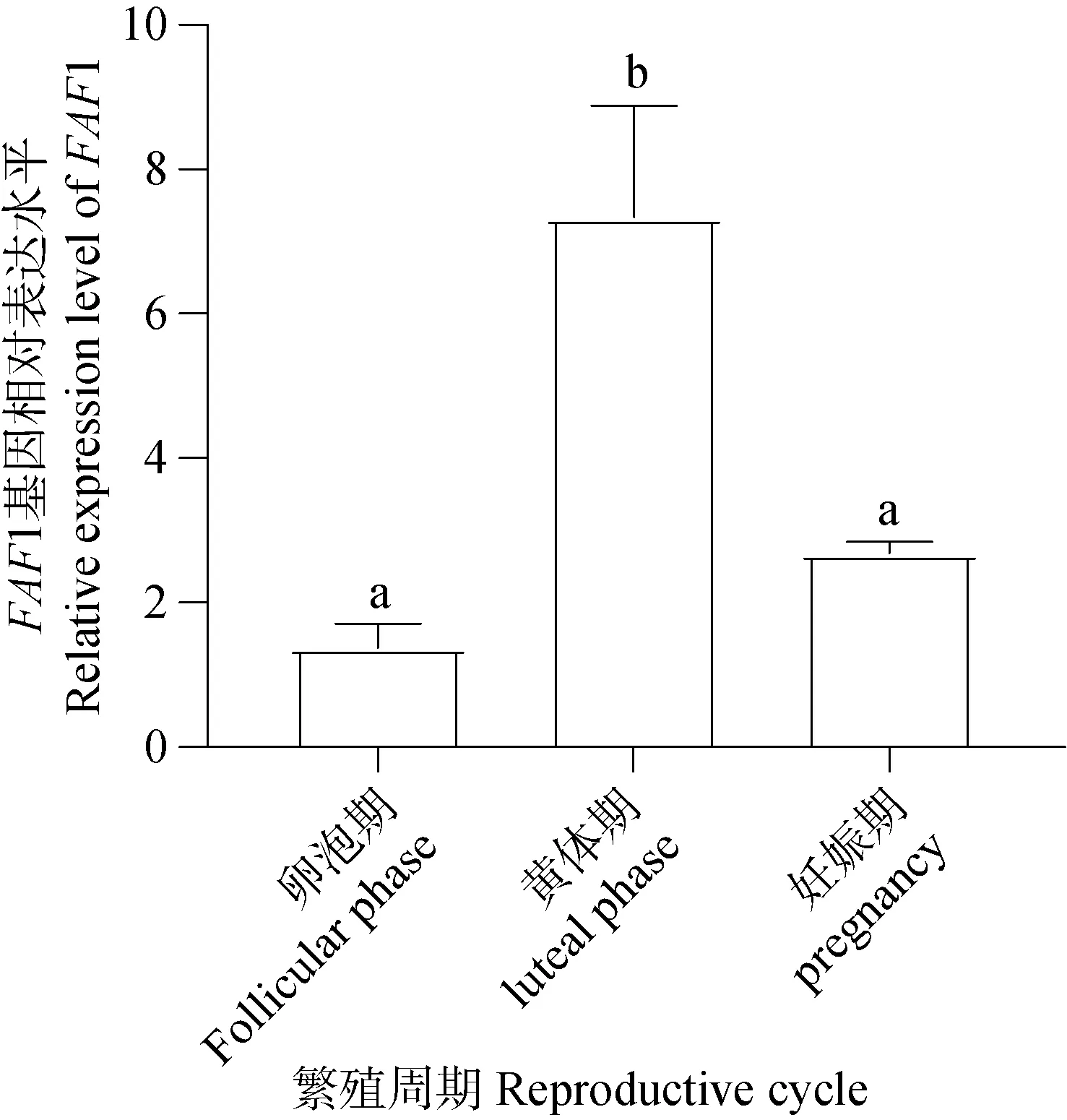
圖12 FAF1 mRNA在不同繁殖周期卵巢的表達定位
2.11 FAF1 蛋白在不同繁殖周期卵巢的相對表達水平
Western blot檢測結(jié)果(圖13)顯示,FAF1蛋白普遍存在于牦牛不同繁殖周期的卵巢組織中。經(jīng)統(tǒng)計分析結(jié)果(圖14)顯示,卵泡期和妊娠期卵巢FAF1蛋白相對表達量顯著高于黃體期,卵泡期卵巢表達量最高,妊娠期卵巢次之,黃體期卵巢中的FAF1相對表達量最低,FAF1蛋白在牦牛不同繁殖周期卵巢組織中相對表達存在差異性。

1.卵泡期卵巢;2.黃體期卵巢;3.妊娠期卵巢
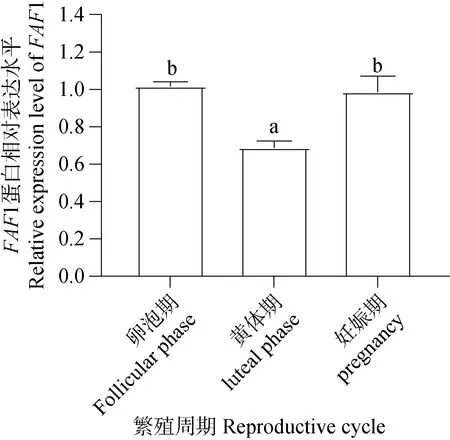
圖14 FAF1蛋白在不同繁殖周期卵巢相對表達水平
2.12 FAF1 蛋白在不同繁殖周期卵巢的表達定位
免疫組織化學染色結(jié)果(圖15)顯示,FAF1蛋白在不同繁殖周期卵巢均存在棕褐色陽性表達。在不同繁殖周期卵巢中表達部位并無明顯的差異,主要表達于卵巢生殖上皮細胞、顆粒細胞、卵丘細胞(圖15A)卵泡膜細胞和黃體細胞(圖15B、15C)。
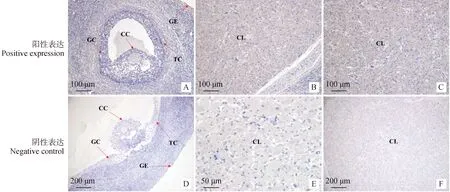
A、D為卵泡期卵巢;B、E為黃體期卵巢;C、F為妊娠期卵巢;GC.顆粒細胞;GE.生殖上皮細胞;TC.卵泡膜細胞;CL.黃體細胞;CC.卵丘細胞
3 討 論
FAF1含有多個功能結(jié)構(gòu)域,主要包括N末端UBA、C端UBX、DEDID、FID[11]。UBA能夠募集對FAF1介導(dǎo)的細胞凋亡和應(yīng)激反應(yīng)至關(guān)重要的多泛素化蛋白[12],從而介導(dǎo)細胞凋亡和應(yīng)激反應(yīng)[13-14]。UBX與泛素-蛋白酶體系統(tǒng)中的分子伴侶含valosin的蛋白(valosin-containing protein,VCP)相結(jié)合[15],VCP是酵母中多功能Cdc48p和爪蟾中多功能p97的哺乳動物同源物[16],它是與多種細胞活動相關(guān)的ATP酶家族的成員,在細胞周期調(diào)節(jié)、凋亡、轉(zhuǎn)錄激活、細胞器生物發(fā)生、囊泡轉(zhuǎn)運、內(nèi)質(zhì)網(wǎng)降解等多種生理過程中起分子伴侶的作用[17]。
DEDID可與Fas相關(guān)死亡域(Fas-associated protein with death domain,FADD)蛋白及caspase-8相互作用增強或者啟動Fas介導(dǎo)的凋亡[18])。FID 與IKKβ直接相互作用來阻止IKKα、IKKβ和IKKγ/NEMO復(fù)合物形成,從而抑制NF-κB信號傳導(dǎo),FAF1也被稱為IκB激酶(IκB kinase,IKK)激活的抑制劑[14]。除此之外FAF1還可以通過干擾NF-κB的RelA亞基(p65)的核易位而抑制NF-κB的活性[19]。近年來,相關(guān)的研究報道還表明FAF1是一種抑癌基因,在多種腫瘤組織和細胞中的表達都是降低的,例如胃癌[20]、肺癌[21]、乳腺癌[22]等,FAF1表達下調(diào)可能導(dǎo)致腫瘤的發(fā)生。FAF1還通過抑制MAVS來調(diào)節(jié)抗病毒免疫過程[21]。
睪丸是雄性動物的生殖腺,是精子形成和睪酮分泌的主要器官[23]。哺乳動物睪丸內(nèi)精子發(fā)生是一個復(fù)雜而又十分精細的生理過程,其中包括凋亡的調(diào)節(jié)機制,正常的精子在發(fā)生的過程中就存在自發(fā)性的生殖細胞凋亡現(xiàn)象[24-25],睪丸生殖細胞凋亡這一過程排除了受到損傷或具有畸形染色體的生殖細胞,既保證了遺傳信息傳遞的準確性,又使得生殖細胞從支持細胞獲得充足的營養(yǎng)和支持[26],而過度的凋亡又會使得生殖細胞數(shù)量過少,形成的精子數(shù)量減少,從而導(dǎo)致雄性個體生育能力下降。Fas信號通路與生精細胞的敏感性密切相關(guān),可能主導(dǎo)生精細胞凋亡[27],FAF1為Fas通路的重要調(diào)控元件,對于Fas通路活化具有重要的調(diào)控作用[28]。本研究表明隨著牦牛年齡的增長,從幼年到青年,再至老年,FAF1 mRNA及FAF1蛋白普遍表達于各個年齡階段,但在不同年齡又有存在差異性,這與Adham等[6]在小鼠睪丸上的研究結(jié)果一致。FAF1 mRNA相對表達量在3、4周歲時升高,6周歲下降,7周歲再次升高,后又下降,差異化表達表明FAF1在睪丸發(fā)育及精子發(fā)生等生理過程發(fā)揮重要生物學作用。FAF1 mRNA相對表達量隨著雄性牦牛達到性成熟而增加,可能是由于性成熟睪酮分泌量的改變,生殖細胞數(shù)量增加,造成睪丸生殖細胞凋亡增加[29],從而FAF1 mRNA相對表達量增加,老年時表達量下降則可能是由于睪酮分泌量減少,性功能降低,精子發(fā)生減少,相應(yīng)凋亡也減少;FAF1蛋白相對表達量在青年初期(3、4周歲)時下降,后逐步升高,與FAF1 mRNA相對表達量不盡相同,可能因為 FAF1蛋白不能單獨啟動細胞凋亡程序,除了在人類BOSC23細胞中可直接啟動細胞凋亡[30],FAF1蛋白需與其他蛋白結(jié)合形成死亡效應(yīng)絲(death effector filament,DEF)或DISC發(fā)揮凋亡作用[31],因此,FAF1蛋白相對表達量降低可能是形成了蛋白復(fù)合體來發(fā)揮凋亡作用而導(dǎo)致。FAF1蛋白主要表達于不同年齡的精子、精子細胞、精原細胞、初級精母細胞、支持細胞、睪丸間質(zhì)細胞和管周肌樣細胞中。支持細胞是為生殖細胞增殖分化提供營養(yǎng)和支持;睪丸間質(zhì)細胞是分泌睪酮及影響支持細胞分化;管周肌樣細胞主要是協(xié)同支持細胞,為精原細胞提供基質(zhì),產(chǎn)生不同階段的生精細胞[32];提示FAF1可能參與睪丸發(fā)育、精子發(fā)生及睪酮分泌等重要生理過程。
卵巢在動物生殖中占有極其重要的地位,因為它不僅能產(chǎn)生卵細胞,還能分泌數(shù)種性激素。FAF1 mRNA相對表達量在黃體期卵巢顯著高于卵泡期和妊娠期,妊娠期卵巢次之,卵泡期卵巢表達量最低。在卵巢組織中,閉鎖卵泡、排卵時的卵巢上皮和黃體發(fā)生時都會發(fā)生細胞凋亡[33]。黃體期卵巢時由于形成黃體,孕激素分泌增加,FAF1 mRNA表達量顯著上升,Taniguchi等[34]發(fā)現(xiàn)牛的黃體期卵巢Fas mRNA表達水平顯著高于其他階段,FAF1是Fas家族成員,為Fas通路重要調(diào)控原件,本研究結(jié)果與Taniguchi等[34]研究結(jié)果相似,推測FAF1 mRNA表達量受孕激素調(diào)控,可能參與黃體溶解的生理過程。FAF1蛋白相對表達量在卵泡期卵巢顯著高于黃體期和妊娠期,妊娠期卵巢次之,黃體期相對表達量最低。FAF1蛋白表達情況與FAF1 mRNA表達情況相反,這可能是因為一般情況下FAF1蛋白需要與其他蛋白形成復(fù)合蛋白才能發(fā)揮其生物學作用,與上述FAF1蛋白在睪丸中表達情況相似,因此導(dǎo)致黃體期卵巢中FAF1蛋白表達量較低,用以形成復(fù)合物溶解黃體。FAF1表達于不同繁殖周期卵巢生殖上皮細胞、顆粒細胞、卵丘細胞和黃體細胞。生殖上皮在卵細胞形成中是不可或缺的因素[35];顆粒細胞的增殖分化與卵泡的發(fā)育密切相關(guān)[36],卵泡閉鎖其實質(zhì)就是顆粒細胞凋亡而產(chǎn)生的結(jié)果[37];卵丘細胞通過縫隙連接調(diào)節(jié)卵母細胞中激素蛋白質(zhì)和相關(guān)調(diào)節(jié)因子的表達,從而影響卵母細胞成熟[38];卵泡膜細胞可以和顆粒細胞,卵母細胞發(fā)生相互作用,在卵泡發(fā)育過程中提供結(jié)構(gòu)支持,還可以合成雄激素[39];黃體細胞在妊娠建立及黃體維持、發(fā)育和退化中具有重要作用[40]。而FAF1在雌性牦牛不同繁殖周期差異性表達,因此推測FAF1蛋白可能參與卵泡閉鎖、發(fā)育、成熟和黃體溶解等生理過程。
4 結(jié) 論
本研究成功構(gòu)建pET-28a-FAF1原核表達載體,并優(yōu)化其誘導(dǎo)溫度,誘導(dǎo)表達后的融合蛋白為不可溶性蛋白,經(jīng)鎳柱純化后,利用Western blot鑒定,成功獲得純度較高的FAF1重組蛋白,免疫試驗動物后獲得兔源FAF1多克隆抗體,效價為1∶1 024 000,具有良好特異性。FAF1在不同年齡睪丸及不同繁殖周期卵巢組織中均有表達,且不同年齡睪丸及不同繁殖周期卵巢表達水平存在顯著差異,揭示FAF1在牦牛不同階段中具有重要生物學作用,推測FAF1可能與雄性牦牛的睪丸發(fā)育、精子發(fā)生及睪酮分泌密切相關(guān),與雌性牦牛的卵泡閉鎖、發(fā)育、成熟和黃體溶解生理過程相關(guān),但相關(guān)機制有待進一步深入研究。

- 畜牧獸醫(yī)學報的其它文章
- 單寧酸對低劑量T-2毒素誘導(dǎo)小鼠結(jié)腸黏膜損傷與菌群失調(diào)的保護效應(yīng)
- 基于網(wǎng)絡(luò)藥理學和試驗驗證分析小檗堿治療雞沙門菌感染的作用機制
- 內(nèi)質(zhì)網(wǎng)應(yīng)激預(yù)適應(yīng)對LPS誘導(dǎo)的山羊子宮內(nèi)膜上皮細胞炎性反應(yīng)的保護作用
- 基于RNA-Seq技術(shù)研究枸杞多糖對環(huán)磷酰胺致雛雞免疫抑制的拮抗機制
- 阿司匹林丁香酚酯顆粒劑有關(guān)物質(zhì)檢測方法的建立與驗證
- 黑果枸杞花青素對低氧誘導(dǎo)的H9c2大鼠心肌細胞凋亡的影響