鶴頂蘭種子的真菌共生萌發研究
2023-09-24 07:27:04高鑫禎邵士成王曉靜馬長樂
種子 2023年7期
汪 雨, 高鑫禎, 邵士成, 王曉靜, 羅 艷, 馬長樂
(1.西南林業大學園林園藝學院, 昆明 650224; 2.中國科學院西雙版納熱帶植物園, 云南 勐臘 666303;3.國家林業和草原局西南風景園林工程技術研究中心, 昆明 650224; 4.中國科學院大學, 北京 100049)
鶴頂蘭屬(Phaius)植物為地生蘭,屬于蘭科吻蘭族,全球約有40種,分布于非洲熱帶地區、亞洲熱帶和亞熱帶地區至大洋洲[1-2]。鶴頂蘭屬植物植株高大,花大且艷麗多姿,很多種類具有優良的園藝觀賞特征。該屬中應用最為廣泛的物種是鶴頂蘭[Phaiustancarvilleae(L'Hér.) Blume],花朵靚麗,優雅欣然,古人稱其為“鶴蘭”,具有極高的觀賞價值,已應用在鮮切花、園林植物配置、園藝美化及園藝品種育種等領域[3]。鶴頂蘭的假鱗莖粗壯,內用具有祛痰止咳、活血止血的作用,外用可治療跌打腫痛、外傷出血等[4]。由于生境破壞和人為過度采挖,加之其自然結實率低,自然更新能力較弱,致使其野生資源已瀕臨枯竭。探索鶴頂蘭保育的技術方法已成為加強鶴頂蘭資源保護和可持續利用中亟待解決的重要問題。
自然環境中,蘭科植物種子必須依賴共生真菌提供營養進行萌發[5-8]。近年來,多種蘭科物種都篩選到了可促進種子萌發的共生真菌,例如地生蘭類的龍頭蘭[Pecteilissusannae(L.) Raf.][9]、手參[Gymnadeniaconopsea(L.) R. Br.][10],附生蘭類的硬葉蘭(CymbidiummanniiRchb. f.)[11]、鐵皮石斛(DendrobiumofficinaleKimura &Migo)[12]等。徐錦堂等[13]從腐生蘭科植物天麻(GastrodiaelataBlume)的原球莖中分離得到一株促進天麻種子萌發的共生真菌,已廣泛應用于天麻的人工繁殖和生產栽培中。Shao等[14-15]建立了原地共生萌發的野外回歸技術,采用共生真菌與種子混合進行播種,成功實現了齒瓣石斛(DendrobiumdevonianumPaxton)的野外回歸。這些研究均表明,蘭科種子的真菌共生萌發是進行蘭科資源保護和可持續利用最為有效的方法。
目前,鶴頂蘭的栽培繁殖方式主要采用分株和無菌播種。分株繁殖通常在休眠期結合換盆進行,分株后的新株需帶有兩個以上的假鱗莖和芽,這種分株方式當年即可開花[16]。但分株繁殖的繁殖系數低,不適用于大規模生產。現階段在生產實踐中已實現了基于種子無菌萌發的組織培養技術,可以進行鶴頂蘭種苗的規模化生產。如蘭芹英等[17]對鶴頂蘭快繁技術的研究中篩選得到萌發率可達100%的培養基。姚紹嫦等[18]主要對組培過程中原球莖的增殖進行了研究,得到一種培養基配方,在培養30 d時增殖倍數可達6.67倍,培養60 d后超過30倍。但組織培養方式耗費巨大的人力物力,不能完全解決生產和保護實踐的應用,同時容易過度縱容幼苗獲得營養和理想的生長條件[19],使幼苗在進行馴化時,可能面臨氣候變化和病蟲害等在實驗室培養中不存在的問題。
在中國科學院西雙版納熱帶植物園(以下簡稱版納園)保育的鶴頂蘭的栽培養護過程中發現,其種子可以在栽培基質中自然萌發,深圳仙湖植物園的研究人員也開發了利用腐熟處理的栽培基質促進鶴頂蘭種子萌發的技術[20],這些發現表明,促進鶴頂蘭種子萌發的真菌可能廣泛存在,但目前仍不清楚是何種真菌可促進鶴頂蘭的種子萌發。本研究采用蘭科種子原地共生萌發的方法,從鶴頂蘭栽培基質中采集自然萌發的原球莖,從原球莖中分離菌株,篩選促進鶴頂蘭種子萌發和原球莖發育的真菌,以期為鶴頂蘭的生物技術育種、種質資源保存、快速繁殖提供理論依據和技術支持。
1 材料與方法
1.1 研究地點和研究材料
鶴頂蘭(圖1A)為地生蘭,廣泛分布于我國臺灣、福建、香港、海南、廣西、云南和西藏東南部,生長于亞熱帶和熱帶常綠闊葉林的林下潮濕處。鶴頂蘭在版納園已成功實現遷地保護。本研究的研究地點位于版納園的保育苗圃(21°54′N,101°46′E)。2021年4月,對栽培于保育苗圃的鶴頂蘭進行人工異交授粉,于2021年9月采集果莢變軟但尚未完全開裂的成熟蒴果(圖1B)。

注:A為花序;B為蒴果;C為TTC染色的種子;D為原地萌發誘導形成的原球莖。圖1 鶴頂蘭Fig.1 Phaius tancarvilleae
1.2 種子收集保存及原地共生萌發
用脫脂棉沾取75%的乙醇清洗果莢表面,再用無菌水沖洗3次。用無菌刀將果實剖開,輕拍果皮,將種子抖落。短期保存:用無菌的濾紙包好種子,在22 ℃條件下,用無水氯化鈣干燥3 d后裝入密閉離心管中,在4 ℃的條件下儲存。長期保存:將種子干燥3 d后存于滅菌的密閉離心管中,置于20 ℃條件下儲存。
采用TTC(氯化三苯基四氮唑)法進行鶴頂蘭種子活力的檢測,使用磷酸緩沖液作為溶劑配制濃度為1%的TTC作為染液。將種子置入1 mL離心管中,加入純水浸泡24 h后再加入預先配置的TCC染液,放置于37 ℃恒溫箱在黑暗條件下染色24 h后,在體式顯微鏡(Nikon SMZ800N)下隨機觀察種子的染色情況,統計視野中的種子總數和著色種胚的種子數。
2021年10月將成熟的鶴頂蘭種子直接撒播在成年鶴頂蘭植株(共計5盆)附近的土壤基質中。每隔一個月對土樣進行監測觀察。
1.3 原球莖內共生真菌的分離
將馬鈴薯葡萄糖培養基(PDA,馬鈴薯200 g/L+葡萄糖20 g/L+瓊脂15 g/L)配制好后在121 ℃高壓滅菌20 min,分裝于無菌培養皿中。將在苗圃中收獲的原地萌發的鶴頂蘭原球莖在自來水下清洗,用軟毛筆去除原球莖表面的土壤及雜質。在超凈工作臺上,將洗凈的原球莖浸入0.1%次氯酸鈉溶液中消毒10 min,用無菌水沖洗3~4次,用無菌刀將原球莖切成薄片,切口面貼于PDA培養基上,用封口膜將培養皿封好后放入霉菌培養箱內,(25±2)℃避光培養。每隔3 d觀察一次。
待原球莖切口附近長出菌絲時,在超凈工作臺用無菌接種針在長出真菌菌落的原球莖切片邊緣處挑取菌絲,接種到新的PDA培養基上進行純化培養,純化后的菌株以PDA試管斜面4 ℃保存備用。
1.4 菌株的分子鑒定及系統位置分析
將保存在PDA培養基中的真菌菌株,在無菌條件下接入盛有液體PDA培養基的錐形瓶中,置于搖床(SPH-310A,上海百典儀器設備有限責任公司,中國)(25±2)℃下震蕩培養。培養時間視真菌的生長速度而定,一般為3~6 d。抽濾約100 mg的菌絲用于真菌基因組DNA的提取,提取方法按照DNeasy plant mini kit(Qiagen69104)說明書上的方法步驟進行。參照常用的真菌分子鑒定方法,使用正向引物ITS1(5′TCCGTAGGTGAACCTGCGG 3′)和反向引物ITS4(5′TCCTCCGCTTATTGATATGC 3′)對核糖體DNA上大約650 bp的ITS區域進行PCR擴增反應[21]。擴增產物送至生工生物工程(上海)股份有限公司進行純化和雙向測序。擴增所得rDNA-ITS序列拼接后使用Geneious prime2022.0.2軟件進行手工校對。測序序列使用GenBank(https://www.ncbi.nlm.nih.gov/genbank/)上的BLAST工具進行序列比對,當rDNA-ITS序列相似性超過95%時,屬于同一個屬,當相似性超過97%時,屬于同一種[22]。
為了確定從鶴頂蘭原球莖中分離得到的真菌的系統發育位置,本研究將分離得到的真菌的ITS序列與從NCBI中獲得的近緣種序列構建系統發育樹。以Armillaria sinapina(FJ495039)為外類群,通過PhyloSuite v1.2.2平臺查找最適模型后使用IQ-TREE構建Maximum Likelihood(ML)系統發育樹[23],通過FigTree v1.4.0軟件進行系統發育樹的編輯。
1.5 真菌對種子萌發的有效性檢測
將保存的供試菌株(編號為HDL-2)在PDA培養基上進行活化,在(25±2)℃黑暗條件下培養8~10 d,待菌絲長滿培養皿。采用燕麥瓊脂培養基(OMA,燕麥5 g/L+瓊脂10 g/L,調節pH值為5.6~5.8)作為種子和真菌共培養的培養基。將保存的鶴頂蘭種子經1% NaClO溶液消毒12 min,于無菌水中漂洗3~5次,然后浸于0.1%的瓊脂溶液中制成無菌種子懸浮液,采用移液槍吸取1 mL(約含580粒種子)均勻播種在培養基表面。用移液槍槍頭將在PDA培養基上的菌絲打孔制成菌餅,取3個菌餅成三點狀放置于距培養皿邊緣約1 cm處。另取相同大小的未接菌的空白PDA培養基打孔制成餅狀放置于OMA培養基中作為陰性對照。另外,將鶴頂蘭種子放置在改良的1/2MS培養基(1/2MS+10%椰汁+蔗糖30 g/L+瓊脂5.5 g/L+活性炭1.0 g/L,調節pH值為5.6~5.8)上進行播種作為無菌萌發的對照培養。播種方法與前述一致。每組播種實驗設置6個重復。將播種后的培養皿放置于組培間進行培養,培養條件:溫度為(25±2)℃光照強度為1 800~2 500 lx,光照周期為光照12 h/黑暗12 h。每周監測種子的萌發情況,記錄和觀測種子萌發及原球莖發育的情況。按照表1劃分的種子萌發階段的標準,在體視顯微鏡下統計處于各萌發階段的種子、原球莖、幼苗數量,計算種子萌發率(G)及各階段的種子、原球莖或幼苗的比率(C)。即G=g/t;C=c/t,其中,g為培養皿中已萌發的種子數,c為培養皿中各階段的種子、原球莖或幼苗的數,t為播種的種子總數。利用SPSS23.0軟件,對不同處理的種子萌發率及各階段的萌發種子、原球莖或幼苗比率進行顯著性差異分析。

表1 鶴頂蘭種子萌發的不同發育階段Table 1 Developmental stages of seed germination of Phaius tancarvilleae
1.6 原球莖被真菌侵染情況的觀察
為檢測鶴頂蘭種子與共培養的真菌的共生關系是否成功建立,培養至60 d時,用臺盼藍染色法觀察鶴頂蘭種子及原球莖被真菌侵染的情況。將不同發育階段的鶴頂蘭種子/原球莖用10% KOH在90 ℃下透明處理3 h,用0.3%過氧化氫洗滌3次,再用30%過氧化氫進行漂白處理40~60 min(視材料大小決定漂白時間),最后在0.05%臺盼藍醋酸甘油溶液中染色10 min,經醋酸甘油溶液脫色后在光學顯微鏡下觀察并拍照。
2 結果與分析
2.1 種子活力及種子原地共生萌發
鶴頂蘭種子經TTC染色后觀察發現,絕大部分種子種胚發育良好,絕大部分能被染為淺橙色,極個別種子未被染色,少數種子染色較深(圖1C)。以種子被染色計為具有活力。結果表明,本研究所用種子95.62%都具有活力。
將鶴頂蘭種子直接撒播在成年植株附近,至第6個月,土壤中開始有肉眼可見的原球莖產生,大小為(1~1.5)mm×(2~4)mm,黃白色,不規則的長圓錐形(圖1D)。
2.2 原球莖真菌的分離與鑒定
采用組織塊分離法從鶴頂蘭原球莖中共計分得5株真菌菌株。形態學和ITS分子鑒定的結果(表2)顯示,5株真菌菌株分別為鐮刀菌屬(Fusarium)2種Fusariumsolani(HDL-1,圖2A)和Fusariumoxysporum(HDL-3,圖2B)、球殼菌屬1種(Plectosphaerellacucumerina,HDL-4,圖2C)、裸囊菌屬1種(Gymnoascusdankaliensis,HDL-5,圖2D)和膠膜菌屬1種[24](Tulasnellasp.,HDL-2,圖2E)。將分離獲得的真菌與鶴頂蘭種子進行共培養,僅HDL-2可以促進鶴頂蘭種子萌發,而其他真菌均未能促進種子萌發。菌株HDL-2在PDA培養基上的菌落呈淡黃色至透明狀(圖2E),菌絲呈圓柱狀,具隔膜和分枝,有厚垣孢子產生(圖2F),生長速度約為2 mm/d。
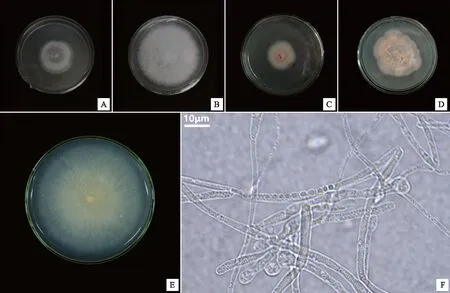
注:A~E分別為菌株HDL-1、HDL-3、HDL-4、HDL-5、HDL-2菌落形態,F為菌株HDL-2菌絲結構。圖2 鶴頂蘭原球莖中分離得到的菌株Fig.2 Fungi strains isolated from protocorms of Phaius tancarvilleae
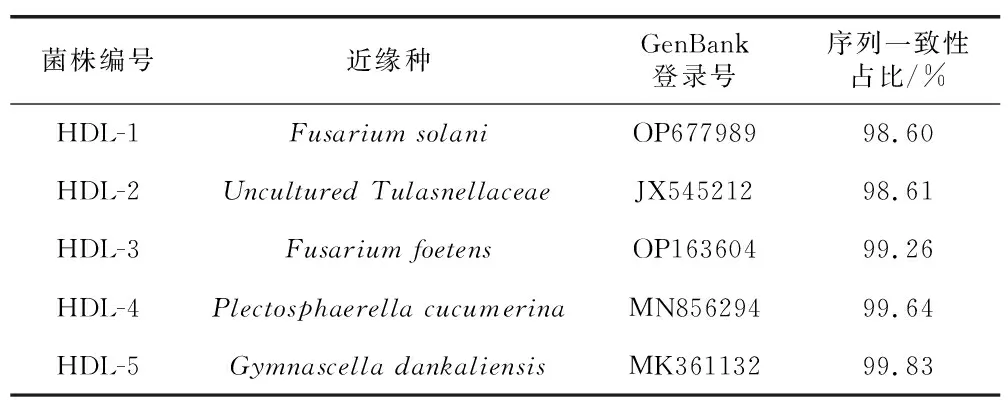
表2 鶴頂蘭原球莖中分離得到的真菌菌株的分子鑒定結果Table 2 Molecular identification of fungal strains isolated from protocorms of Phaius tancarvilleae
根據該菌株的ITS序列與GenBank上的比對結果,下載了相似性較高的蘭科菌根真菌的ITS序列構建系統發育樹。系統發育分析結果(圖3)顯示,從鶴頂蘭原球莖中分離所得的真菌菌株HDL-2與Uncul tured Tulasnellaceae(JX545212)聚為一支,支持率為99%,與其他膠膜菌屬菌株聚成一支較大的分支,支持率為95%,表明該菌株屬于膠膜菌屬。
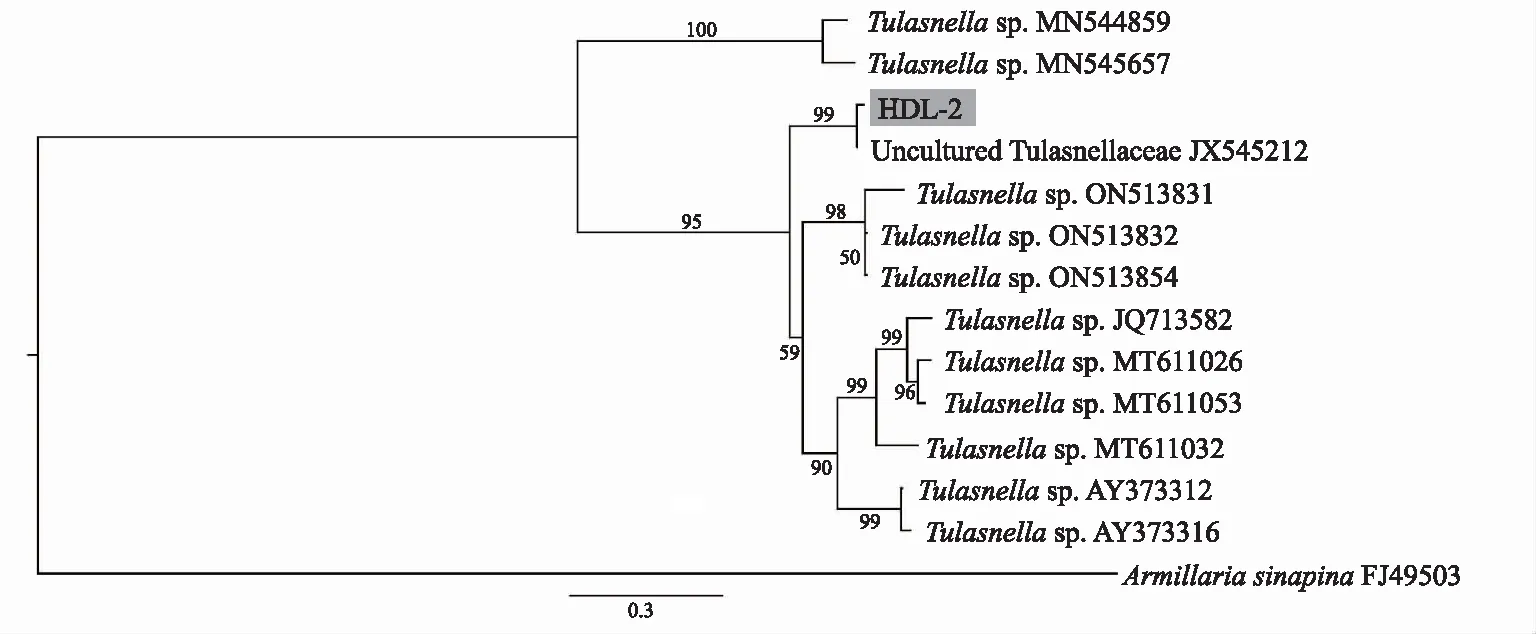
注:節點上的數字表示支持率,以Armillaria sinapina為外類群。圖3 基于核糖體ITS序列的最大似然法系統發育樹 Fig.3 Maximum-likelihood tree constructed based on rDNA-ITS sequences
2.3 鶴頂蘭種子共生萌發的發育過程
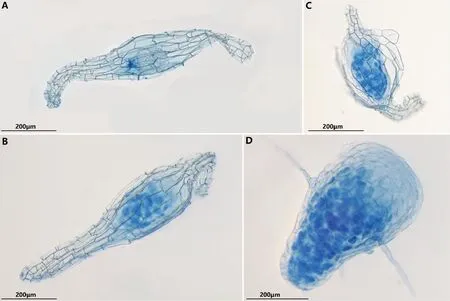
將HDL-2菌株與鶴頂蘭種子在OMA培養基上進行共培養試驗,觀察發現,HDL-2菌株可以有效促進鶴頂蘭種子萌發。不接菌的對照組種子未見萌發跡象。種子接菌共培養20 d之前無明顯變化,保持未萌發狀態(圖4A);共培養20~30 d時,可以明顯觀察到種胚吸水膨脹,種皮仍然包裹在種胚周圍,達到萌發的第1階段(圖4B);30~40 d之后,種胚繼續膨脹和增大,種皮在一側破裂,頂端逐漸轉綠,形成原球莖,到達發育的第2階段(圖4C);40~60 d后,原球莖繼續長大,明顯呈現極性分化,發育形成頂部寬圓基部細尖的錐形原球莖,同時在原球莖除頂部外的位置生出細長絨毛狀的假根(圖4D);80~100 d后,原球莖頂部出現突起(圖4E),此時處于萌發的第3階段;100~120 d后,頂端的突起繼續伸長發育,形成葉芽,逐步發育形成第一片幼葉(圖4F),隨著頂端繼續生長,形成了莖及多數葉(圖4G),此時均處于萌發的第4階段;150~180 d后,從莖基部形成幼根(圖4H),到達萌發的第5階段,形成幼苗(圖4I)。
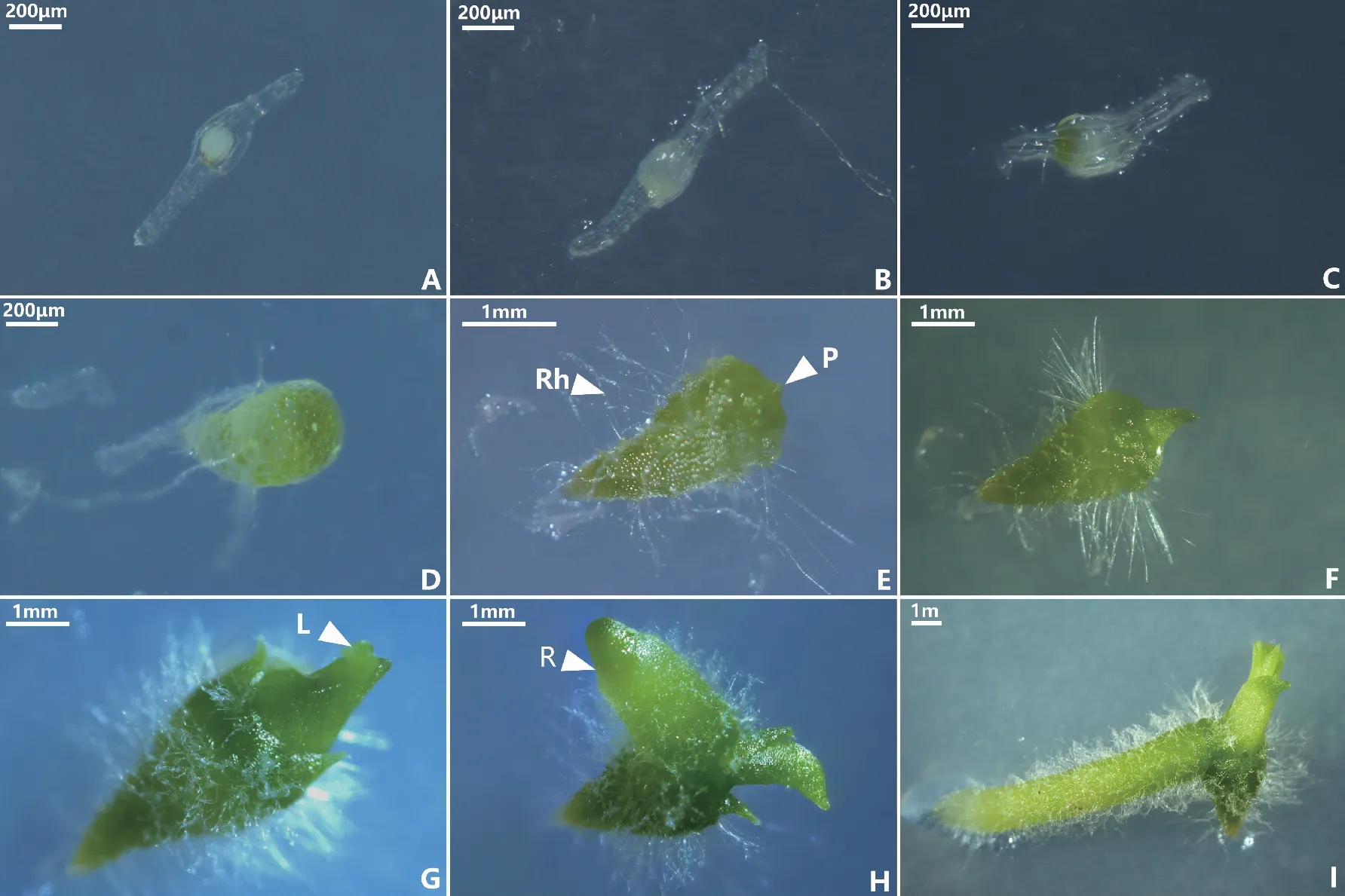
注:A為萌發第0階段,未萌發的種子;B為萌發第1階段,種胚吸水膨脹;C為萌發第2階段,種胚變綠,一側突破種皮; D為萌發第3階段,錐形原球莖;E為萌發第3階段,假根(Rh)和突起(P)產生;F為萌發第4階段,形成葉芽; G為萌發第4階段,莖及多數葉形成;H為萌發第5階段,幼根形成;I為幼苗形成。圖4 鶴頂蘭種子共生萌發過程 Fig.4 Symbiotic seed germination process of Phaius tancarvilleae
2.4 真菌侵染情況的觀察
將與HDL-2共培養的不同發育階段的種子或原球莖采用臺盼藍染色進行觀察,發現未萌發的種子真菌從種孔進入,聚集在胚柄端,但未侵入種胚(圖5A),說明此時共生關系尚未建成,而萌發的不同發育階段的種子或原球莖均被臺盼藍染色。處于吸水膨脹期間的種子可見種胚內基部細胞被染為藍色,說明鶴頂蘭種子在萌發的第1階段,真菌就開始侵入定殖(圖5B)。種胚突破種皮時(第2階段),原球莖細胞內可見大量被染為藍色的菌絲團(圖5C)。隨著原球莖體積不斷膨大、長出根毛(第3階段),原球莖細胞中仍存在大量菌絲團(圖5D)。臺盼藍染色結果表明,HDL-2菌株的菌絲在種子萌發階段可成功定植在種子及原球莖細胞內,表明共生關系已成功建立。
注:A為種子未萌發,真菌聚集在胚柄端,尚未侵染胚;B為萌發第1階段,膨大的胚中已有真菌侵染; C為萌發第2階段,原球莖被菌絲團侵染;D為萌發第3階段,錐形原球莖中下部被菌絲團侵染。圖5 臺盼藍染色顯示鶴頂蘭種子不同萌發階段真菌侵染情況 Fig.5 Trypan Blue Staining showed fungal infection in different stages of seed germination of Phaius tancarvilleae
2.5 HDL-2促進鶴頂蘭種子萌發的效率
本研究的3組種子不同處理間的種子萌發效率存在顯著差異(表3,圖6)。不接菌的陰性對照組沒有觀察到種胚明顯膨脹,所有種子均在第0階段(圖7C)。培養30 d時,種子在接種HDL-2的OMA培養基上的萌發率為22.09%,19.51%處于第1階段,有2.59%進入第2階段,種子在不含真菌的改良1/2MS培養基上的萌發率為17.18%,均處于第1階段,兩者之間差異不顯著(圖7A)。60 d時,共生培養和無菌培養的萌發率差異顯著,接菌的OMA培養基的種子總萌發率為38.56%,并且有13.64%的種子發育到第3階段,少數種子進入到第4階段,1/2MS培養基中種子的總萌發率為29.56%,種子的發育僅處于第1、第2階段。80 d時,接菌的OMA培養基上種子萌發率達到(58.33±3.48)%,有25.96%的種子處于第3階段,改良的1/2MS培養基中種子的總萌發率為40.90%,到達第3階段的種子僅為1.83%,兩者之間差異極顯著。110 d時,接菌培養的OMA培養基的種子總萌發率為86.69%,已有5.57%的原球莖發育成幼苗,到達第4階段(圖7A),此時改良的1/2 MS培養基上種子的總萌發率為52.04%,僅有(0.55±0.34)%的種子發育到第4階段,大部分種子停留在第2階段或第3階段(圖7B),共生萌發和無菌萌發存在極顯著差異。

注:A為與菌株HDL-2共培養的OMA培養基;B為1/2MS培養基;C為無真菌參與的OMA培養基。圖7 培養110 d時,不同處理下的鶴頂蘭原球莖發育Fig.7 At culture of 110 days, protocorm development of Phaius tancarvilleae under different treatments
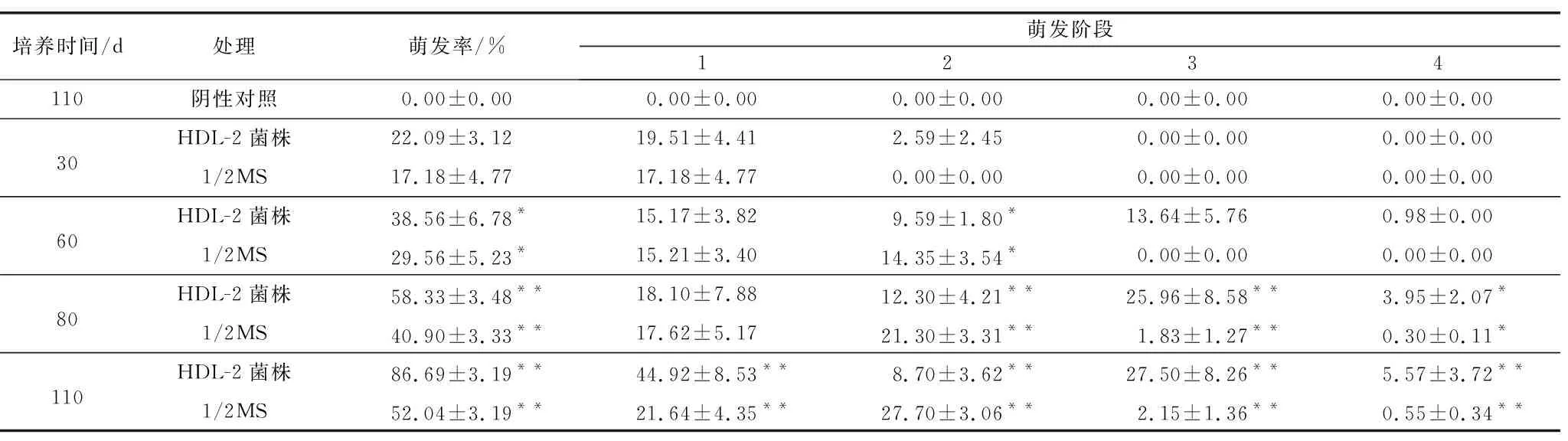
表3 不同處理下鶴頂蘭種子的萌發率、萌發各階段的比率Table 3 The seed germination rate and the ratio of germination stages of Phaius tancarvilleae under different treatments
3 討 論
在自然條件下,蘭科植物的種子必須與真菌產生共生關系,必須依賴共生真菌提供營養才能萌發[5-8]。在沒有共生真菌協助的情況下,蘭科種子基本無法完成萌發過程,多數只能吸水膨脹至種皮破裂階段[25]。篩選可促進蘭科種子萌發的有效共生真菌,在蘭花資源的保護和可持續利用上具有重要的意義。鶴頂蘭是廣泛應用的園藝觀賞花卉,規模化生產技術并不成熟。Cheng等[26]從臺灣本土蘭科植物中分離出4種絲核菌屬(Rhizoctonia)真菌,分別接種于鶴頂蘭的組培幼苗,發現菌株R15可提高幼苗的成活率、增加植物高度和葉片長度,菌株R19可促進鶴頂蘭開花,菌株R02則可顯著增強光合作用的速率。這些研究均是基于鶴頂蘭的幼苗展開的,而關于鶴頂蘭種子的真菌共生萌發目前未見報道。本研究通過原地共生萌發技術,首次從鶴頂蘭的原球莖中分離獲得的真菌HDL-2,可成功實現鶴頂蘭種子的共生萌發,該真菌屬于膠膜菌科膠膜菌屬真菌,屬于典型的蘭科菌根真菌。目前已報道的對蘭科植物種子有促萌發作用的真菌主要集中在絲核類真菌,包括蠟殼菌屬(Sebacina)、膠膜菌屬和角擔菌屬(Ceratobasidium)[27-28]。膠膜菌屬真菌是最常見的蘭科植物菌根真菌[29-31],可以有效促進蘭科植物種子萌發。如Youm等[32]從蝦脊蘭(CalanthediscolorLindl.)的根中分離出了來自膠膜菌屬的促萌發真菌;Zi等[33]從兜唇石斛[Dendrobiumaphyllum(Roxb.) C. E. Fishc.]原球莖中分離出有效促進種子萌發的膠膜菌屬真菌(Tulasnellasp.)FDaI7;Shao等[34]從鼓槌石斛(DendrobiumchrysotoxumLindl.)原球莖中分離出的膠膜菌可以促進鼓槌石斛種子萌發。除此之外,小菇屬(Mycena)、瘤菌根菌屬(Epulorhiza)、鬼傘屬(Coprinellus)等其他屬真菌也有報道。如徐錦堂和郭順星[13]從天麻的原球莖中分離出了促萌發真菌紫萁小菇(Mycenaosmundicola);孫曉穎等[35]從帶葉兜蘭的原球莖及小幼苗中分離出的共生真菌屬于瘤菌根菌屬;Gao等[36]從杜鵑蘭[Cremastraappendiculata(D. Don) Makino]根中分離出的共生真菌屬于鬼傘屬。
大量研究表明,從成年植株根中分離獲得的真菌種類多樣,但往往未必能夠促進種子萌發或形成幼苗。現在普遍認為最有效的手段是采用原地誘捕法獲得蘭科植物的共生萌發真菌[11,27]。Zi等[33]、孫曉穎等[35]、孟元元[37]均使用了該方法,在不同石斛屬蘭花的原生地埋下種子,當種子萌發形成原球莖后,從原球莖中成功分離獲得促進種子萌發的有效真菌。本研究也采用了原地共生萌發的技術,但誘導鶴頂蘭種子萌發的環境基質并非從野外環境獲得,而是使用了人工栽培的基質。從鶴頂蘭的人工栽培基質中獲得了原球莖,從中分離到了膠膜菌屬真菌能促進鶴頂蘭種子萌發,表明該真菌可能在自然環境中廣泛存在。而該菌株是否也存在于鶴頂蘭的野生環境中,自然環境中的鶴頂蘭種子萌發機制還需進一步的研究探明。
共生萌發通常具有較高的萌發效率[38]。陳婭婭等[39]使用了21株菌根真菌對鵝毛玉鳳花[Habenariadentata(Sw.) Schltr.]種子進行了共生萌發研究,發現其中有8株可以顯著提高種子的萌發效率。對幽靈蘭[Dendrophylaxlindenii(Lindl.) Benth. ex Rolfe]種子萌發的研究中,Dlin-394菌株(角擔菌屬)可以顯著提高種子萌發率,在真菌浸染9周后種子萌發率最高,為84%,而使用改良的1/4MS培養基進行無菌萌發的種子萌發率僅為40%左右,且共生萌發的種子形成幼苗的速率也較無菌萌發的高[40]。Zhou和Gao[41]的研究發現,在相同培養時間下,鳳蝶蘭[Papilionantheteres(Roxb.) Schltr.]和Epa-01(瘤菌根菌屬)共培養時的種子萌發率要顯著高于在AGS培養基上進行無菌培養的種子萌發率,且所處的萌發階段也更高,分別在15,30,45 d時,共生培養的原球莖的鮮重均極顯著高于無菌培養。Gao等[36]研究發現,杜鵑蘭在與DJF-10(鬼傘屬)菌株共生培養6周后的種子萌發率超過80%,相比之下,在無菌的MS培養基上種子萌發率僅9.68%,遠低于共生培養,且共生萌發條件下13周后就能形成幼苗,但在MS培養基上無菌萌發的原球莖需要5個多月才能發育成幼苗。與前人的研究相似,本研究中鶴頂蘭種子與HDL-2菌株(膠膜菌屬)共生萌發的效率要顯著高于無菌萌發。鶴頂蘭種子在無菌萌發培養基上培養110 d后萌發率為(52.04±3.19)%,大部分種子處于0,1,2階段;在接種了HDL-2菌株的OMA培養基上,110 d時萌發率可達(86.69±3.19)%,大部分種子處于第3階段。在不加激素和其他物質的情況下,鶴頂蘭種子在MS、Kyato、KC三種基本培養基中的萌發率為20%~30%[42]。由此可見,HDL-2真菌對鶴頂蘭種子萌發的促進作用明顯,且效率顯著高于無菌萌發培養基。本研究建立的共生萌發手段為鶴頂蘭作為園藝資源在生產實踐中的開發和利用提供了新的技術和方法。
蘭科植物種子的共生萌發機制極其復雜,相關的研究具有挑戰性。缺乏高效的促萌發真菌限制了對蘭科植物資源的保護和利用。本研究通過原地萌發技術獲得的一種膠膜菌屬真菌對鶴頂蘭的種子萌發和幼苗形成有促進作用,可用于鶴頂蘭的人工繁育生產和野外回歸的保護實踐,并可為真菌與蘭科植物的互作關系研究提供科學資料。
