長葉竹柏葉綠體基因組特征及系統發育分析
2023-09-24 07:27:06孔維杰楊雪蓮吳永飛田山君
種子 2023年7期
關鍵詞:物種
顏 麗, 孔維杰, 楊雪蓮, 吳永飛, 王 霞, 田山君
(貴州大學農學院, 貴陽 550025)
長葉竹柏(Nageiafleuryi)為羅漢松科、竹柏屬,主要分布在中國、越南、柬埔寨等東南亞國家,在《中國植物紅皮書》中被列為瀕危植物。長葉竹柏果仁可用作食用油、工業用油原料[1]以及木材原料,也是治療風濕病[2]和發燒[3]的中草藥,是具有較高經濟價值的珍稀樹種。近年來,隨著植物藥用成分分離技術水平的提高,國內外學者對竹柏科植物進行了大量的研究,結果顯示,該科植物含有二萜、二萜內酯、甾體、倍半萜、苯丙素、木脂素、三萜及雙黃酮等成分[4-5]。目前,對竹柏科植物所含的二萜內酯以及雙黃酮等成分進行了大量的生物活性研究,表明竹柏科植物具有抗腫瘤、抗病毒、抗氧化及抗炎活性功能[6]。
葉綠體主要是綠色植物進行光合作用的重要場所,在其他代謝途徑中也起關鍵作用[7],具有半自主遺傳性。葉綠體大多具有保守的環狀四分體結構,但不同物種間會存在一定差異,主要表現為IR區域長度和方向的不同[8]。葉綠體研究技術發展迅速,現已應用于植物系統發育[9]和種群分析[10]等方向的研究,對杧果(Mangiferaindica)[11]、香花枇杷(Eriobotryafragrans)[12]、柳葉芹(Czernaevialaevigataturcz.)[13]、燈盞花(Erigeronbreviscapus)[14]、蒜頭果(Malaniaoleifera)[15]、鐵皮石斛(Dendrobiumcandidum)[16]、大花君子蘭(Cliviaminiata)[17]、沙棗(Elaeagnusangustifolia)[18]和喜馬紅景天(Rhodiolahimalensis)[19]等的研究為植物的基因組學、種群分類及系統發育分析等提供了一定的理論依據。迄今為止,對長葉竹柏的相關研究主要是其化學成分、栽培引種及病蟲害等方面,關于長葉竹柏的完整葉綠體基因組研究尚未見報道。本研究利用高通量測序對長葉竹柏葉綠體進行基因測序、組裝和注釋,分析其系統發育關系,旨在為鑒定該物種、豐富其葉綠體遺傳信息、進一步探索其個體水平的遺傳差異及遺傳改良奠定理論基礎。
1 材料與方法
1.1 試驗材料
長葉竹柏葉片來自貴州省植物園(26°37′20″N,106°43′29″E),將新鮮、無病蟲害樣本經液氮速凍后于-80 ℃冷藏保存。
1.2 試驗方法
1.2.1長葉竹柏DNA提取及測序
采用CTAB法[20],從150 mg新鮮葉片樣本中提取全基因組DNA,送至惠通生物有限公司檢測,DNA檢測合格后將其打斷生成300~500 bp的小片段,在序列末端修復、3′端加A、加測序接頭,再經過純化、瓊脂糖凝膠電泳,PCR擴增等一系列操作構建好整個長葉竹柏的測序文庫,經質檢合格的文庫,使用Illumina高通量測序平臺Nova Seq6000進行測序。
1.2.2長葉竹柏葉綠體全基因組的組裝、注釋
測序數據采用SPAdes軟件進行基因組拼接[21],使用已公布的近緣葉綠體數據與蛋白編碼基因序列作為參考,利用Blastn與Exonerate軟件將結果與參考基因組進行比對,確定序列組裝。使用PRICE和MITObim軟件[22]對目標序列進行延伸、合并及拼接,盡量減少Scaffold數目。迭代拼接的結果使用Bowtie2軟件[23],將原始測序Reads進行回帖,挑出匹配成對的,利用軟件SPAdes進行重拼接,直至完成環狀基因組。然后利用PGA(Plastid Genome Annotator)軟件[24]進行基因組注釋,設置默認參數,完成注釋。采用Chloroplot軟件[21]繪制長葉竹柏葉綠體基因組圖譜。
1.2.3長葉竹柏葉綠體密碼子偏好性和簡單重復序列分析
運用Codon W1.4.2軟件[25]統計分析長葉竹柏葉綠體基因組密碼子的偏好性RSCU(Relative Synonymous Codon Usage);利用MISA(MIcroSAtellite identification tool)提供的Perl程序[26]分析簡單重復序列(Simple Sequence Repeat,SSR)位點,程序在Windows命令窗口運行。其中,設置單核苷酸最低重復次數為10,二核苷酸為5,三核苷酸、四核苷酸、五核苷酸、六核苷酸的均為4。
1.2.4長葉竹柏葉綠體基因組系統進化分析方法
從NCBI數據庫下載與24個與長葉竹柏近緣物種的葉綠體全基因組序列,構建最大似然(ML)樹。利用MAFFT7.037[27]和FFT-NS-2策略將24個不同物種完整葉綠體基因組中的共有蛋白編碼基因與長葉竹柏的基因進行比對。運行Model-finder var1.6來選擇最佳模型,并選擇TVM++F+I+G4模型[28]。使用RAxML var8.2.9構建系統發育樹,自舉檢測值設置1 000[29]。
2 結果與分析
2.1 長葉竹柏葉綠體基因組基本特征
本研究獲得的長葉竹柏葉綠體基因組序列數據在NCBI(https://www.ncbi.nlm.nih.gov/)的GenBank中的登錄號為OL435123[30]。該葉綠體基因組呈雙螺旋環狀結構(圖1),大小為133 870 bp,因缺乏反向重復序列(IR)結構,所以它不屬于典型的四分體結構。
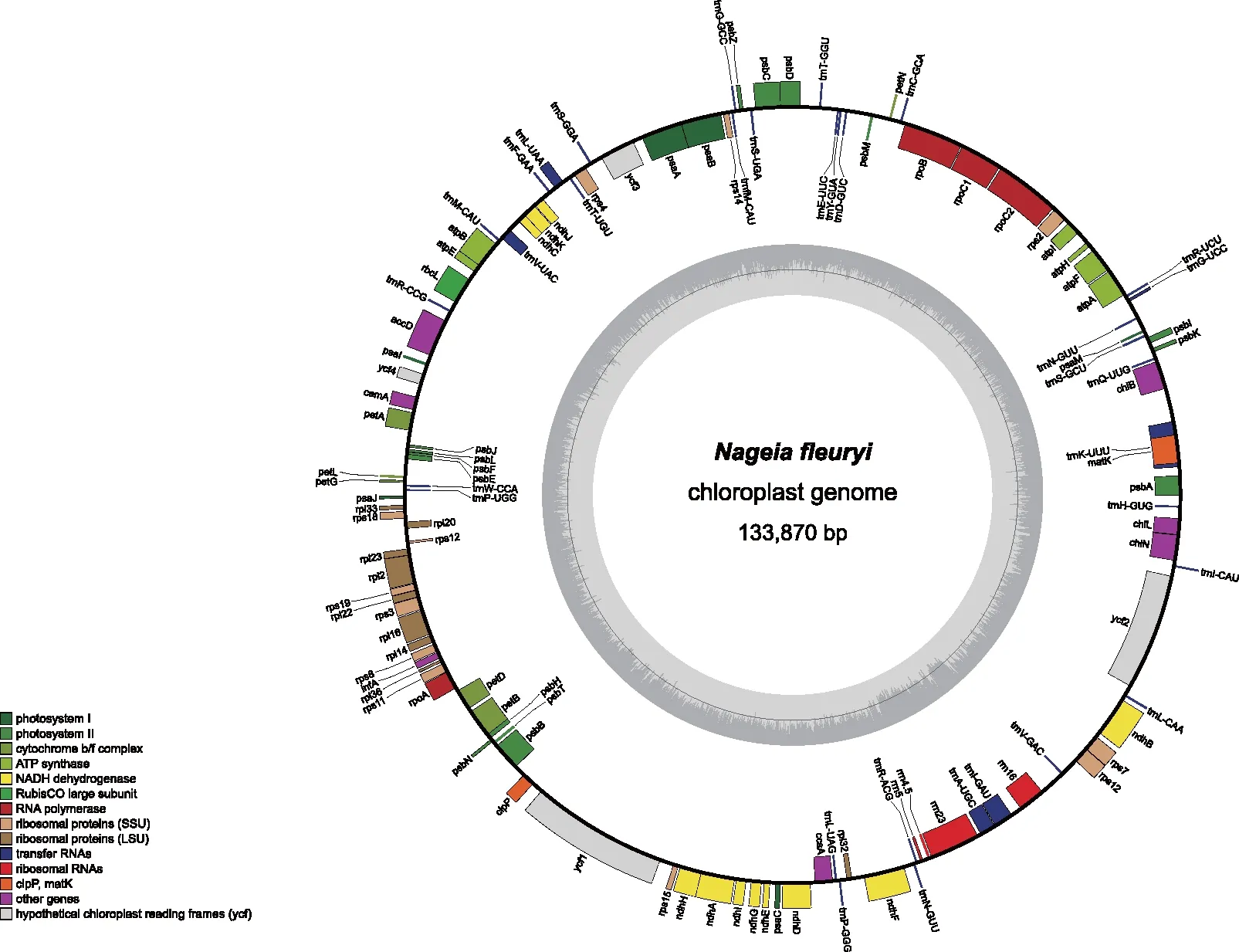
圖1 長葉竹柏葉綠體基因組環形圖譜 Fig.1 Circularized map of the chloroplast genome of Nageia fleuryi
長葉竹柏葉綠體基因組編碼119個基因(表1),包括蛋白編碼基因(80個)、tRNA基因(35個)和rRNA基因(4個),其中,有11個基因(ndhF、ndhA、ndhB、rpl2、rpoC1、tRNA-Lys、tRNA-Gly、tRNA-Leu、tRNA-Val、tRNA-Ile、tRNA-Ala)含有1個內含子,2個基因(clpP和ycf3)含有2個內含子,rps12具有反式剪接。因基因功能各不相同,長葉竹柏葉綠體基因組可分為光合作用基因(43個)、自我復制基因(64個)、其他編碼蛋白基因(8個)和未知功能基因(4個),占比分別為36%,54%,7%和3%。
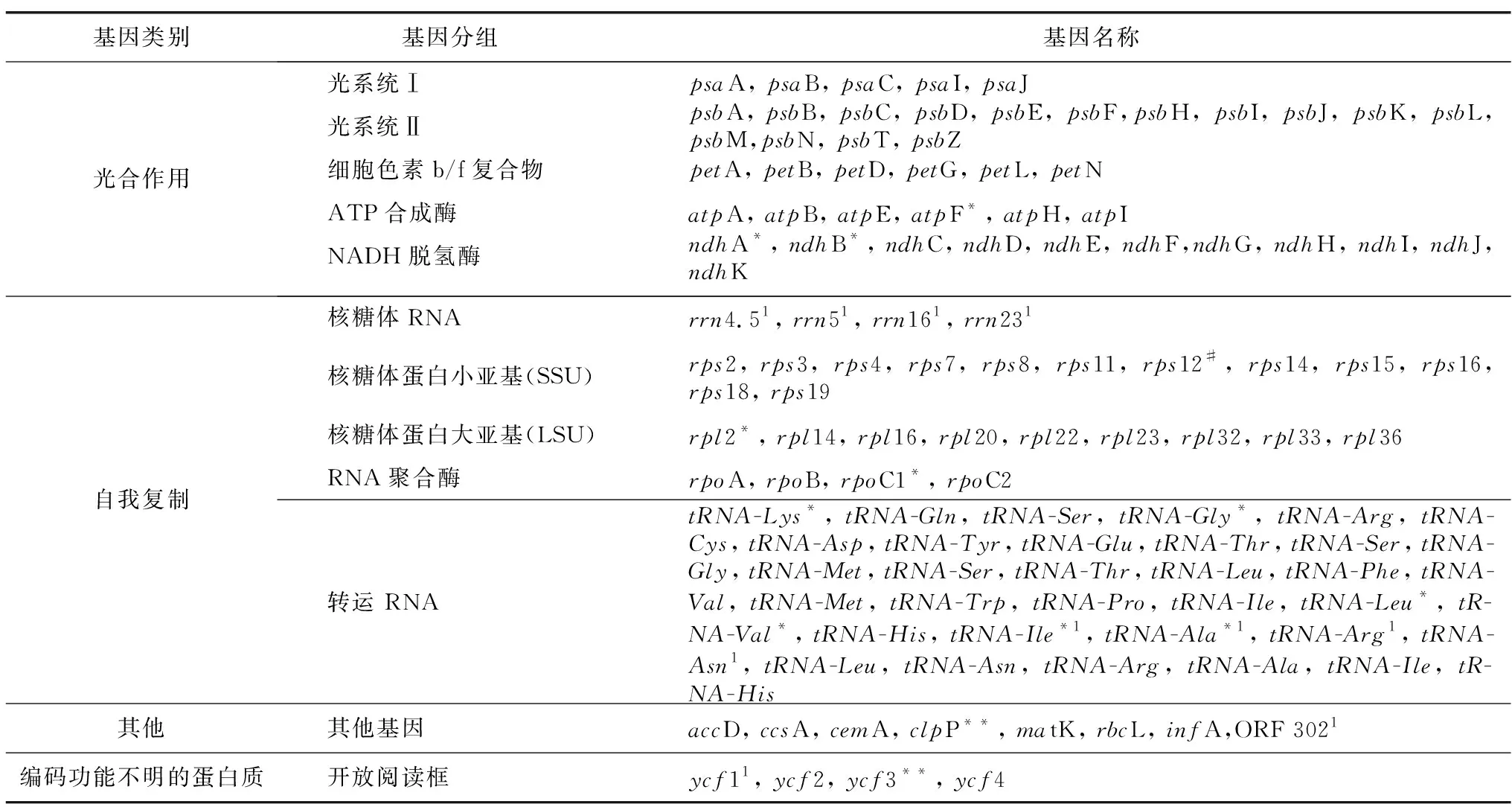
表1 長葉竹柏葉綠體基因組注釋信息Table 1 Gene annotation information of the chloroplast genome of Nageia fleuryi
2.2 長葉竹柏葉綠體偏好密碼子分析
真核生物基因組存在64個密碼子,通常編碼20種不同氨基酸和3個終止密碼子,其中,編碼同一氨基酸的不同密碼子被使用的頻率也不一樣,被頻繁使用的密碼子是“偏好密碼子”,而其他密碼子則是“非偏好密碼子”,這種現象被稱為“密碼子偏好性”。一般認為,密碼子偏好性的成因是不同密碼子對應的tRNA在細胞里的豐度不一樣,tRNA豐度越高,對應的密碼子的使用頻率也會越高。偏好密碼子分析結果(表2)表明,長葉竹柏葉綠體基因組共檢測出44 623個密碼子,編碼亮氨酸(Leu)的密碼子數量最多,為4 703個,占總密碼子數的10.54%;色氨酸(Trp)的密碼子數量最少,為629個,占總密碼子數的1.41%。長葉竹柏葉綠體測序結果顯示,密碼子具有明顯的偏好性,RSCU等于1的氨基酸有2個,分別為甲硫氨酸(Met)和色氨酸(Trp),RSCU大于1的密碼子種類有28個,其中只有2個密碼子的堿基以G結尾,其余所有密碼子均以A/U結尾。
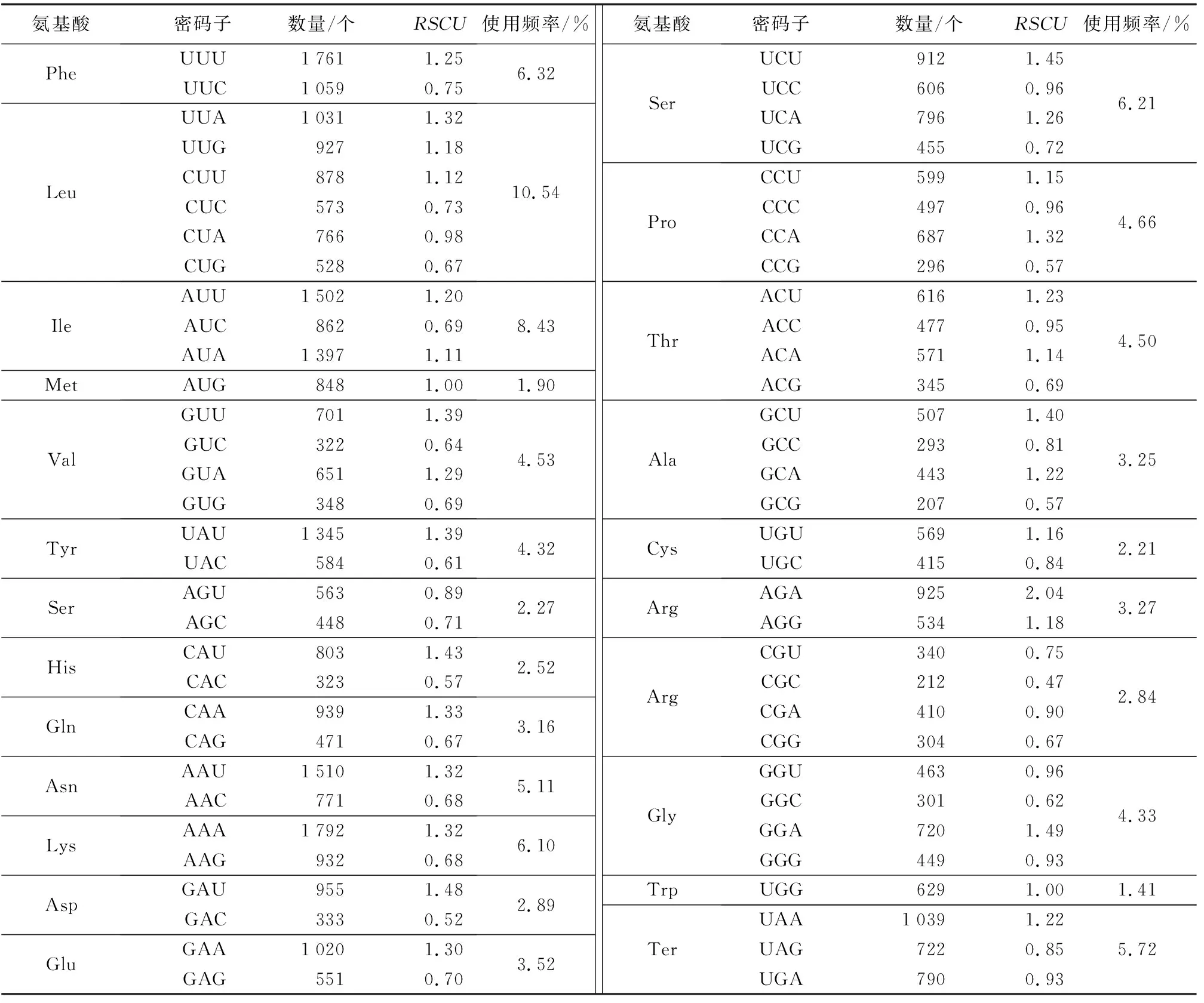
表2 長葉竹柏各氨基酸相對同義密碼子使用度Table 2 The RSCU for amino acid of Nageia fleuryi
2.3 長葉竹柏葉綠體簡單重復序列(SSR)分析
表3顯示,長葉竹柏葉綠體基因組共49個SSR(包含單核苷酸、二核苷酸和三核苷酸)。其中,單核苷酸重復序列最多,共26個,主要有A(10個)、C(1個)、G(3個)和T(12個),共占53.06%;二核苷酸重復序列共22個,主要是AT(14個)、CA(1個)、TA(6個)和TC(1個)4種重復類型,占44.90%;三核苷酸最少,只有1個,為AAT,占2.04%。
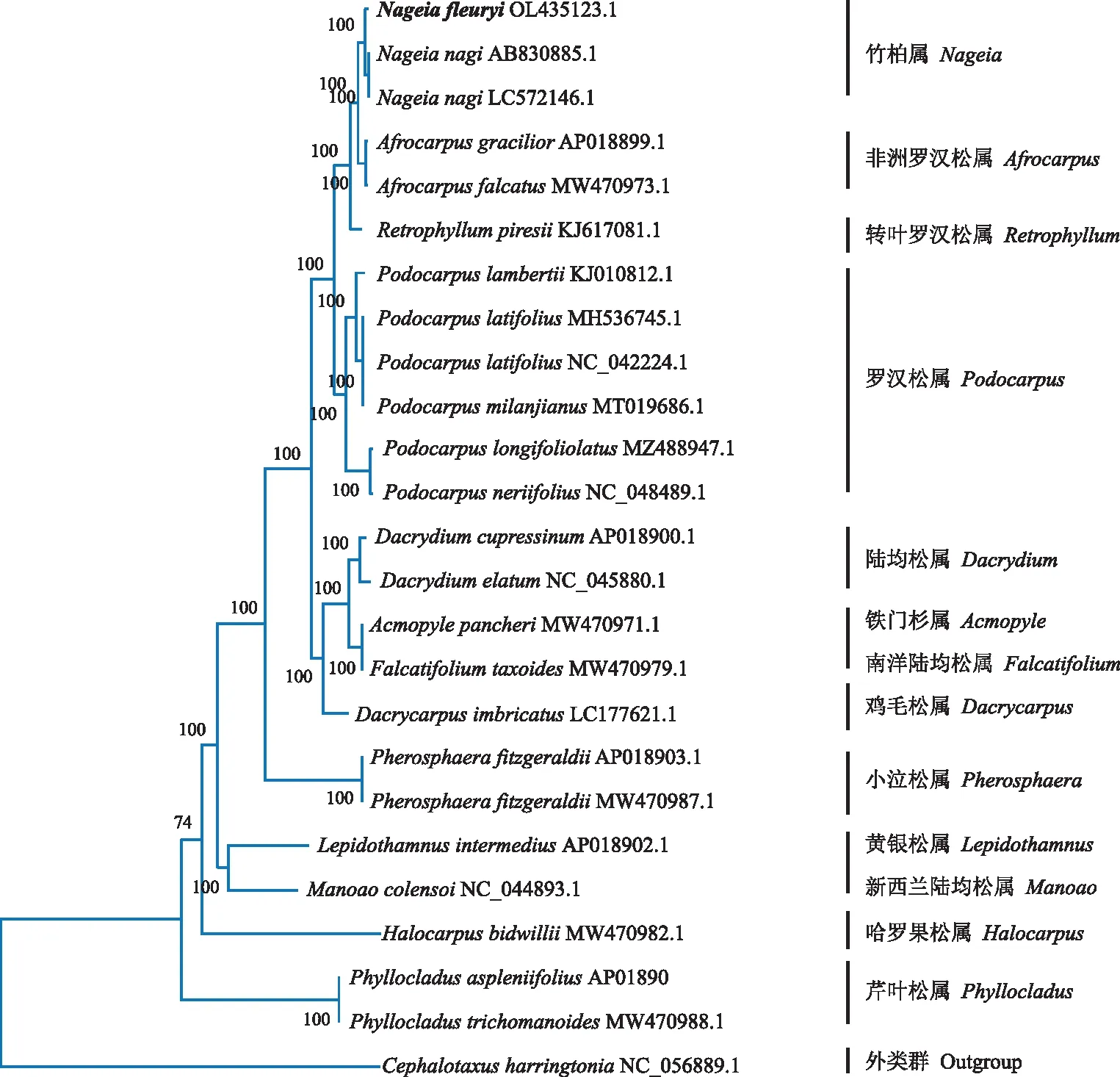
圖2 25個物種葉綠體基因組系統發育樹 Fig.2 Phylogenetic tree based on chloroplast genomic of 25 species
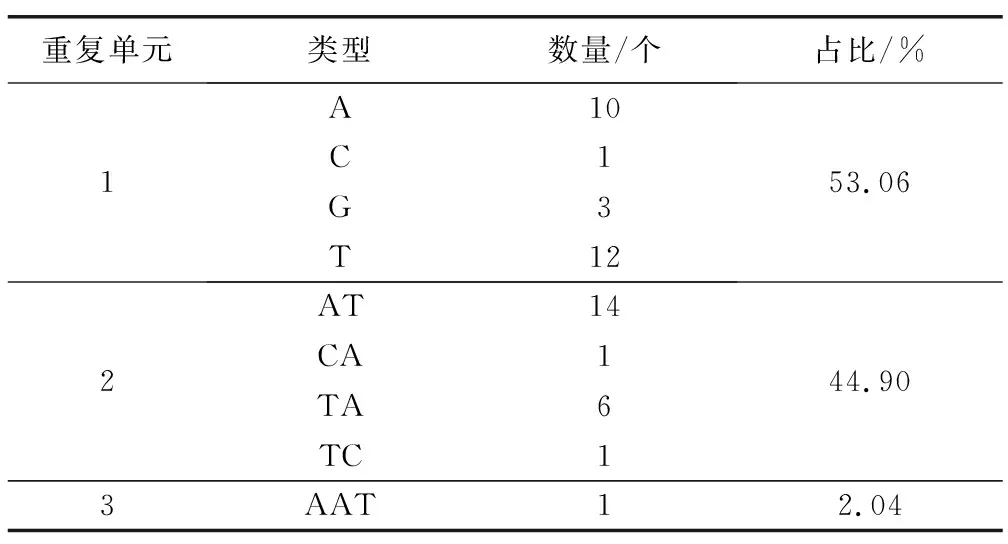
表3 長葉竹柏葉綠體簡單重復序列(SSR)信息Table 3 The SSR information for chloroplast of Nageia fleuryi
2.4 長葉竹柏系統進化樹分析
葉綠體基因組聚類分析對研究植物發育進化有重要意義。本研究選用日本粗榧(Cephalotaxusharringtonia)為外類群及24個羅漢松科物種,構建最大似然(ML)樹。系統發育樹(圖2)顯示,不同科的植物分布不同的進化分支,長葉竹柏與同為羅漢松科的23個物種構成一個單系類群。長葉竹柏與竹柏聚為一支,自展值為100,同為羅漢松科竹柏屬,說明長葉竹柏與竹柏親緣關系最為密切。此外,與其他屬相比,非洲羅漢松屬(Afrocarpus)和轉葉羅漢松屬(Retrophyllum)與竹柏屬(Nageia)親緣關系密切。
3 討 論
Ohyama等[32]和Shinozak等[33]首次測出地錢(MarchantiapolymorphaL.)和煙草(NicotianatabacumL.)葉綠體全基因組序列,為后續無數學者提供了新線索,解決了許多植物分類[34]、葉綠體基因數據庫完善[35]及物種鑒別[36]等方面的關鍵問題。如張慧等[37]研究獲得了完整的益母草屬葉綠體基因組,系統發育樹結果表明其與水蘇屬(Stachys)物種的親緣關系較近;Zhou等[38]通過對大黃屬(Rheum)的研究,開發了超級DNA條形碼。可見,研究植物葉綠體基因組信息對生物進化分析、分類及開發利用具有重要的意義。
物種進化過程中,往往會出現一些基因組重排和葉綠體基因組基因丟失的少量事件。本研究發現,長葉竹柏葉綠體基因組在基因種類和結構上與大多數裸子植物(如羅漢松屬[39]、松科[40]、柏科[41]等)的葉綠體基因組結構類似,都缺失IR區;Chen等[42]研究水杉葉綠體全基因組發現,由于水杉葉綠體基因組的一個大IR拷貝丟失,并且推測其IRA已丟失,但IRB已保留在水杉葉綠體基因組中。魯華等[43]對華山松葉綠體基因組分析結果表明,南系華山松葉綠體基因組也缺乏反向重復區。此外,還有其他少數被子植物(如牻牛兒苗屬植物[44],蒺藜苜蓿、鷹嘴豆、三葉草等豆科植物[45])也因丟失了一個反向重復(IR)區而不具有四分體結構,無法區分其大單拷貝區和小單拷貝區。一般情況下認為,反向重復序列大可起到穩定葉綠體基因組的作用[46],使之能應對大型的結構重排。因此,大反向重復序列損失的植物葉綠體基因組會發生一系列的多變問題,從而導致基因缺失或基因組重排的現象。另外,長葉竹柏及其他缺少IR區物種中的IRA和IRB是否同時缺失或缺失其中之一以及其中的結構變化引起的生物特異性等有待進一步研究探討。
植物葉綠體DNA分子一般長120~170 kb[8],本研究中長葉竹柏葉綠體基因組大小為133 870 bp,屬于一般長度范圍內。葉綠體基因組通常含有100~120個編碼基因,且大多蛋白編碼基因占比較大,能達80%以上,tRNA、rRNA占比較少。長葉竹柏葉綠體共編碼119個基因,其中蛋白編碼基因有80個,約占總數的67%,相較于其他物種,長葉竹柏葉綠體的蛋白編碼基因較少。本研究根據基因的功能將注釋基因分為四大類,分別是光合作用、自我復制、其他編碼蛋白及未知功能,這與黃桃[47]、薄殼山核桃[48]、扁果草[49]等葉綠體基因組功能分類結果相似,可再次證實葉綠體大部分基因主要起到光合作用及基因復制等功能。因此,可通過進一步探究葉綠體基因組表達調控機理,分析葉綠體光合作用及生物合成的相關具體機制。
密碼子偏好性反映基因乃至物種的起源和進化方式,且對基因功能及其編碼蛋白的表達有一定影響。通過密碼子偏好性分析研究發現,長葉竹柏葉綠體基因組共檢測出44 623個密碼子,RSCU等于1的氨基酸有2個(甲硫氨酸和色氨酸),RSCU大于1的密碼子共有28個,其中只有兩個密碼子的堿基以G結尾,其余所有密碼子均以A/U結尾,說明長葉竹柏葉綠體基因具有明顯的AU偏好性。該結果與前人研究結果相似,張雨等[19]研究紅景天葉綠體基因組密碼子偏好性也發現其具有明顯的AU偏向性。此外,在劍麻[50]、翠雀[51]、橄欖[52]、秋茄[53]等葉綠體基因組研究中發現,其密碼子偏好是A或T。說明不同物種的密碼子偏好性存在差異,而這通常是自然選擇和突變等因素影響所致。還有研究表明,不同物種密碼子偏好性所受的主導因素也各不相同,如刺榆[54]受自然選擇的影響;涼粉草[55]則同時受自然選擇和突變壓力的共同影響;紫菜[56]除自然選擇和突變壓力外,可能還受堿基組成等因素的影響。本研究沒有具體探討長葉竹柏葉綠體密碼子偏好性的影響因素,下一步可對其密碼子使用規律及其影響因素進行深入研究。
簡單重復序列(SSR)是指由1~6個核苷酸組成的基本單元,經多次串聯形成重復的一段DNA序列[57],有幾十至幾百條堿基序列,它們分布在葉綠體基因組的不同部位,影響著細胞的多項生命活動,而位于非編碼區的SSR比位于編碼區的變化更大[58]。與核基因組相比,葉綠體基因組較小,SSR數量相對較少,可用于物種鑒定、遺傳多樣性分析和分子輔助育種等[59]。基于長葉竹柏葉綠體基因組SSR分析,發現其大多數單核苷酸由A/T組成,大多數二核苷酸由AT/TA組成,而其余SSR的A/T含量較高,該研究結果與SSRs通常由短聚腺嘌呤(poly A)或聚胸腺嘧啶(poly T)重復序列組成的論點一致[60]。葉綠體基因組SSR中較高的A/T含量也有助于堿基組成的偏差,從而使A/T在長葉竹柏葉綠體基因組中富集。
Zhang等[61]研究指出,葉綠體基因組長度是普通條形碼序列的幾百倍,有足夠的變異位點,完整的葉綠體基因組可作為超級DNA條形碼,應用于物種鑒定及系統發育的關系判定[62]。為了研究羅漢松科中長葉竹柏的系統發育位置,本研究選擇了24個物種,這些物種包括了竹柏科、羅漢松科和紅豆杉科,以紅豆杉科的日本粗榧列為外類群,將這些物種與長葉竹柏葉綠體基因組數據構建系統發育樹。結果表明,長葉竹柏與竹柏的同源性最高,親緣關系最近,該結果確定了長葉竹柏的系統位置及其發育關系。此外,還可對長葉竹柏葉綠體基因組進行其他分析和應用,如SNP位點、數字PCR等分析技術,從而進一步精準鑒別竹柏科植物。
4 結 論
葉綠體基因組注釋表明,長葉竹柏葉綠體基因組全長為133 870 bp,缺乏反向重復區,總GC含量為37%,共119個基因,包括80個蛋白編碼基因、35個tRNA基因和4個rRNA基因,其絕大多數的基因與光合作用及基因轉錄翻譯相關。密碼子偏好性分析表明,亮氨酸(Leu)使用頻率最高(10.54%),色氨酸(Trp)使用頻率最低(1.41%),密碼子偏向使用A和U兩種堿基。共檢測到49個SSR位點,以單堿基(53.06%)重復為主。此外,系統發育分析顯示,長葉竹柏與竹柏親緣關系最密切。該研究為進一步研究羅漢松科分子標記開發、種間鑒定以及該物種的演化過程提供科學依據。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
