硫辛酸對果蠅帕金森病模型神經(jīng)元的作用及其機制*
2023-10-18 13:37:00熊江福張祥明石彩群張春林
貴州醫(yī)科大學學報 2023年9期
熊江福, 張祥明, 石彩群, 張春林*
(貴州醫(yī)科大學 基礎(chǔ)醫(yī)學院 生物學系, 貴州 貴陽 550025)
帕金森病(Parkinson's disease,PD)是一種常見的中老年人神經(jīng)系統(tǒng)退行性疾病[1],主要病理學特征是多巴胺(dopamine,DA)神經(jīng)元的丟失及殘存神經(jīng)元中路易小體(Lewy body,LB)的出現(xiàn)[2-3]。PD發(fā)生的具體機制尚不完全清楚,但研究表明PD的發(fā)生會伴有DA神經(jīng)元中α-突觸核蛋白(α-synuclein,α-syn)異常聚集、氧化應激、線粒體功能障礙等[4]。目前,臨床上還沒有針對PD病因的治療藥物,PD傳統(tǒng)藥物主要是DA制劑和DA受體激動劑,長期使用會出現(xiàn)劑末現(xiàn)象和異動癥等嚴重副作用[5]。因此,尋找針對病因、療效好、副作用少、成本低廉及能夠?qū)D進行疾病修飾治療(disease modification therapy,DMT)的藥物在PD治療中具有重要意義[6]。α-硫辛酸(α-lipoic acid, α-LA)是線粒體能量代謝的輔助因子[7],具有抗氧化作用,且能夠透過血腦屏障對中樞神經(jīng)系統(tǒng)具有保護作用[8]。研究顯示,α-LA對6-OHDA以及魚藤酮誘導的PD細胞模型的細胞活性、氧化應激損傷、線粒體功能障礙等具有顯著的改善作用;此外,α-LA能夠改善魚藤酮誘導的PD模型大鼠的行為學,提高PD大鼠紋狀體的DA含量,降低活性氧(reactive oxygen species,ROS)水平,保護線粒體功能[9-11]。異常表達、聚集的α-syn能通過自噬-溶酶體途徑(autophagy-lysosome pathway,ALP)降解[12-14],α-LA除了具有顯著的抗氧化作用外,其還能夠上調(diào)細胞自噬水平,從而產(chǎn)生對細胞的保護作用[15-16]。果蠅是用于研究各種疾病的理想小型模式生物之一,過表達α-syn果蠅表現(xiàn)出上爬能力異常、DA神經(jīng)元丟失、ROS水平上升、線粒體功能障礙等類似人類PD的病理學特征[17-18]。本實驗利用過表達SNCA.A30P的PD果蠅模型,探討分析α-LA對PD果蠅模型的行為學以及DA神經(jīng)元的保護作用,檢測α-LA對PD果蠅模型氧化應激水平、線粒體功能、自噬以及α-syn表達的影響,進一步分析α-LA的神經(jīng)保護作用,并探索其降低α-syn聚集表達水平的細胞分子機制,為α-LA的臨床治療PD提供實驗依據(jù)。
1 材料和方法
1.1 材料
1.1.1實驗動物 果蠅23868(w*;UAS-2xEGFP;elav-GAL4)、8147(w*;UAS-SNCA.A30P;購自Blomington Drosophila Stock center)、工具果蠅(pin/cyo;T3/T6)、Atg5 RNAi(FBgn0029943)、Atg8a RNAi(FBgn0052672)果蠅(浙江大學昆蟲所黃建華研究員惠贈)。
1.1.2試劑與儀器 α-LA(美國 Sigma),ROS、丙二醛(MDA)、谷胱甘肽(GSH)、超氧化物歧化酶(SOD)及三磷酸腺苷(ATP)檢測試劑盒(南京 建成),兔源酪氨酸羥化酶(tyrosine hydroxylase,TH)抗體(美國 Invitrogen),鼠源GAPDH抗體(中國 康城),鼠源ACTIN抗體(美國 ABclonal),鼠源α-syn抗體(美國 Life Techmologies) ,鼠源NADH脫氫酶(泛醌)鐵硫蛋白3(NDUFS3)抗體(英國 Abcam),鼠源自噬相關(guān)蛋白8a(autopahgy-related gene8a, ATG8a)抗體(美國 Santa Cruz Biotechnology),兔源自噬相關(guān)蛋白5(autopahgy-related gene5, ATG5)抗體(美國Novus Biologicals),羊抗兔二抗、羊抗鼠二抗(美國LI-COR公司),熒光羊抗兔二抗、antifade reagent(美國 Invitrogen);果蠅恒溫培養(yǎng)箱(寧波江南儀器),組織細胞破碎儀(美國Next Advance),多功能凝膠成像系統(tǒng)(美國 Bio-Rad),高速冷凍離心機(金壇金城碩華儀器),低速離心機(湖南湘儀實驗室儀器),恒溫磁力攪拌器(金壇富華儀器),水浴箱、瞬時離心機、立式壓力蒸汽滅菌器(金壇金城碩華儀器),微量加樣器(德國 Eppendof),轉(zhuǎn)膜儀、電泳儀(美國 Bio-Rad);Odyssey雙色紅外熒光成像系統(tǒng)(美國LI-COR),激光共聚焦顯微鏡(日本 Olympus)。
1.2 方法
1.2.1果蠅培養(yǎng)基配制 取1 L去離子水加熱至100 ℃,取瓊脂4.25 g、酵母26.5 g、玉米面50 g、白糖7.25 g、紅糖60 g溶于300 mL水中攪拌均勻后加入1 L熱水中,加入的同時繼續(xù)攪拌直至再次沸騰;加入1.75 g對羥基苯甲酸甲酯,繼續(xù)加熱3~5 min,待冷卻至70 ℃,加入丙酸6 mL攪拌,使用果蠅食物加料機將食物裝入不同果蠅培養(yǎng)管中。在正常果蠅培養(yǎng)基的基礎(chǔ)上,加入2.15 mmol/L α-LA制成加藥果蠅培養(yǎng)基。
1.2.2果蠅雜交 將8147(UAS-SNCA.A30P)處女果蠅與23868(elav-Gal4)雄性果蠅置于同一果蠅瓶內(nèi),果蠅雜交的后代即是PD果蠅。根據(jù)果蠅產(chǎn)卵情況,2~3 d將果蠅倒入新的果蠅瓶內(nèi)。本實驗所使用的果蠅GAL4/UAS系統(tǒng)是一種轉(zhuǎn)基因技術(shù)體系,其原理是利用特定的啟動子或增強子,在果蠅神經(jīng)元激活酵母轉(zhuǎn)錄激活子GAL4的表達,GAL4又以同樣的方式引起GAL4反應元件UAS-SNCA.A30P的轉(zhuǎn)錄。只有將GAL4轉(zhuǎn)基因品系果蠅與UAS-靶基因品系果蠅進行雜交,才可能產(chǎn)生表達靶基因的后代。UAS-SNCA.A30P和elav-Gal4分別與工具果蠅(pin/cyo,T3/T6)雜交,得到+/cyo;elav-GAL4/T6和UAS-SNCA.A30P/pin;+/T3,再將兩品系雜交得到UAS-SNCA.A30P/cyo;elav-GAL4/T3,該品系最后分別與UAS-Atg5RNAi和UAS-Atg8aRNAi雜交最后得到Atg5 RNAi和Atg8a RNAi的PD果蠅。
1.2.3分組 以Elav-GAL4>UAS-SNCA.A30P果蠅品系作為模型(PD)組,Elav-GAL4果蠅品系作為對照(control)組,Elav-GAL4>UAS-SNCA.A30P果蠅品系培養(yǎng)于α-LA加藥培養(yǎng)基中作為干預實驗(LA)組;敲降A(chǔ)tg5(Atg5 RNAi)或Atg8a(Atg8a RNAi)后,Atg5 RNAi和Atg8a RNAi的PD果蠅培養(yǎng)于α-LA加藥培養(yǎng)基中作為Atg5降低+LA組和Atg8a降低+LA組,挑選雜交后羽化0~2 d的雄性果蠅培養(yǎng)于不同培養(yǎng)基中,25 ℃培養(yǎng)箱中培養(yǎng)。
1.2.4果蠅壽命實驗 為了研究α-LA對PD果蠅模型壽命的影響,control組、PD組以及LA組果蠅分別培養(yǎng)在含有不同果蠅食物的果蠅管中,收集不同組剛羽化的雄性果蠅各150只,每組3管,每管50只果蠅,25 ℃培養(yǎng)箱中培養(yǎng),每3 d換新的果蠅食物并記錄死亡果蠅數(shù)。
1.2.5果蠅上爬能力檢測 取在不同培養(yǎng)基中培養(yǎng)的果蠅,二氧化碳麻醉,隨機挑選20只放入20 cm長、15 cm寬的果蠅管中,室溫放置30 min,讓果蠅充分蘇醒,在果蠅管距離管底8 cm處劃線,輕輕敲擊果蠅管底,使果蠅全部掉在管底,記錄10 s內(nèi)爬過8 cm處果蠅數(shù),每組重復10次。上爬率(%)=爬過8 cm處果蠅數(shù)/總果蠅數(shù)×100%。
1.2.6果蠅腦部氧化應激指標(ROS、MDA、GSH、SOD)含量檢測 取在不同培養(yǎng)基中培養(yǎng)25 d的不同組果蠅40只,PBS中取下果蠅腦部組織破碎,10 000 r/min離心20 min,取上清。BCA試劑盒檢測蛋白濃度,2′,7′-二氯二氫熒光素(2′,7′-dichlorodihydrofluorescein,DCFH) 法檢測ROS含量,5,5′-二硫代-雙-(2-硝基苯甲酸)(5,5'-dithiobis-2-nitrobenoicacid,DTNB)法檢測GSH含量,硫代巴比妥酸(thiobarbituric acid)法檢測MDA含量,氯化硝基四氮唑藍(nitrobluetetrazolium,NBT)法檢測SOD含量。
1.2.7果蠅腦部PAL、PPM1/2、PPM3、PPL1、PPL2 PA神經(jīng)元簇檢測 顯微鏡下PBS中取解剖果蠅大腦,4%多聚甲醛中固定10 min,吸取固定液加入PBST洗3次,每次10 min,1%胎牛血清封閉1 h,PBST洗5 min,1∶1 000一抗孵育(24 h),PBST洗3次,每次10 min,1∶2 000二抗孵育(1.5 h),加1滴DAPI于載玻片上,將果蠅大腦放于其中,蓋片。20倍鏡下激光共聚焦顯微鏡觀察結(jié)果,藍色熒光為DAPI染色,標記細胞核作為背景;綠色熒光為TH染色,標記DA經(jīng)元;最后兩結(jié)果合并(MERGE)觀察。
1.2.8果蠅腦部ATP含量檢測 隨機取各組40只果蠅,PBS中取頭部,加入裂解液100 μL,組織破碎后4 ℃離心20 min,10 000 r/min,取上清。不透明96孔板中加入100 μL ATP檢測工作液,室溫放置5 min。在對應孔中加入樣品20 μL,輕輕吹打混勻,間隔2 s后用化學發(fā)光檢測儀檢測相對光單位(RLU),根據(jù)標準曲線計算出樣品中ATP濃度。
1.2.9Western blot檢測果蠅腦部α-syn、NUDFS3、TH、Atg5、Atg8a蛋白表達水平 不同組果蠅隨機取40只,剪取頭部,提取蛋白,BCA法檢測各組蛋白濃度,定量上樣。SDS-PAGE膠電泳分離蛋白,將蛋白轉(zhuǎn)印至PVDF膜,5%脫脂牛奶封閉1 h,分別加入1∶2 000的一抗。4 ℃孵育過夜,TBS洗3次,加1∶4 000的二抗室溫孵育2 h,TBS洗3次后,Odyssey雙色紅外熒光成像系統(tǒng)掃描成像,Image studia軟件分析結(jié)果。
1.3 統(tǒng)計學分析
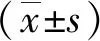
2 結(jié)果
2.1 α-LA對PD果蠅壽命的影響
生存曲線分析結(jié)果表明,與contro組比較,contrL+LA組果蠅的生存率變化不明顯,差異無統(tǒng)計學意義(P>0.05);與control組相比,PD組果蠅生存率明顯降低(P<0.001);與PD組相比,LA組果蠅生存率延長(P<0.05)。見圖1。以上結(jié)果提示α-LA可延長PD果蠅的壽命。
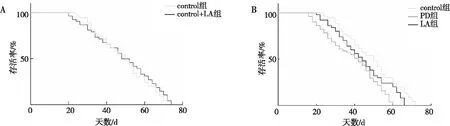
注:A為正常果蠅在加入α-LA培養(yǎng)基與未加入α-LA培養(yǎng)基壽命比較;B為PD果蠅在加入α-LA培養(yǎng)基與未加入α-LA培養(yǎng)基壽命比較。
2.2 α-LA對PD果蠅上爬能力的影響
與control組比較,control+LA組果蠅的上爬率無明顯變化,差異無統(tǒng)計學意義(P>0.05)。在培養(yǎng)20 d后,與control組相比,PD組果蠅上爬率明顯下降(P<0.001);與PD組相比,LA組果蠅上爬率明顯升高(P<0.001)。見圖2。提示α-LA可提高PD果蠅上爬能力。

注:A為正常果蠅在加入α-LA培養(yǎng)基與未加入α-LA培養(yǎng)基不同時間點的上爬率比較;B為PD果蠅在加入α-LA培養(yǎng)基與未加入α-LA培養(yǎng)基不同時間點的上爬率比較; (1)與control組相比,P>0.05;(2)與control組相比,P<0.001;(3)與PD組相比,P<0.001。
2.3 α-LA對PD果蠅腦內(nèi)α-syn表達水平的影響
與PD組比較,LA組果蠅腦內(nèi)α-syn表達水平明顯減少(P<0.01),見圖3。以上結(jié)果提示α-LA可降低PD果蠅腦內(nèi)α-LA的表達。
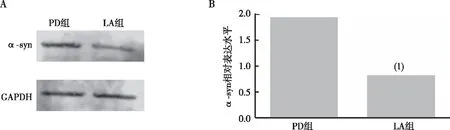
注:A、B分別為PD組與LA組果蠅腦部α-syn蛋白的Western blot結(jié)果及相對含量統(tǒng)計;(1)與PD組相比,P<0.01。
2.4 α-LA對PD果蠅腦部ATP水平的影響
與control組比較,PD組果蠅腦內(nèi)ATP含量明顯減少(P<0.001);與PD組相比,LA組果蠅腦內(nèi)ATP水平明顯增加(P<0.01)。見圖4。以上結(jié)果提示α-LA可增加PD果蠅腦部ATP水平。
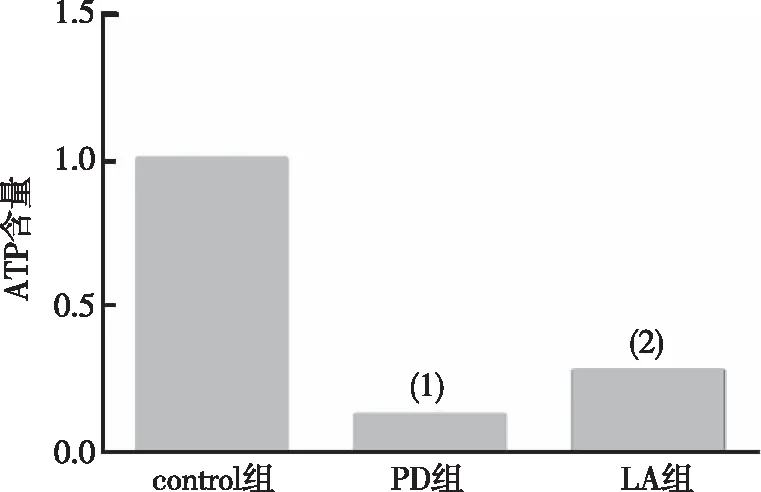
注:(1)與對control組比較,P<0.001;(2)與對PD組比較,P<0.01。
2.5 α-LA對PD果蠅腦部NUDFS3表達的影響
與contro組比較,PD組果蠅腦內(nèi)NUDFS3表達水平下降(P<0.05);與PD組比較,LA組果蠅腦內(nèi)NUDFS3表達增高(P<0.05)。見圖5。以上結(jié)果提示α-LA提高PD果蠅腦部NUDFS3表達。

注:A、B為各組果蠅NDUFS3蛋白的Western blot結(jié)果及相對含量統(tǒng)計;(1)與control組比較,P<0.05;(2)與對PD組比較,P<0.05。
2.6 α-LA對PD果蠅腦部ROS、MDA、GSH及SOD的影響
與control組比較,PD組果蠅腦內(nèi)ROS水平明顯上升(P<0.01);與PD組相比,LA組果蠅腦內(nèi)ROS水平下降(P<0.05)。見圖6A。與control組比較,PD組果蠅腦內(nèi)MDA含量明顯升高(P<0.001);與PD組比較,LA組果蠅腦內(nèi)MDA水平下降(P<0.05);見圖6B。與control組比較,PD組果蠅腦內(nèi)GSH含量降低(P<0.05);與PD組比較,LA組果蠅腦內(nèi)GSH水平下降(P<0.05);見圖6C。與control組比較,PD組果蠅腦內(nèi)SOD含量明顯降低(P<0.01);與PD組比較,LA組果蠅腦內(nèi)SOD含量明顯上升(P<0.01);見圖6D。以上結(jié)果提示α-LA降低PD果蠅腦部氧化應激水平。

注:A~D分別為各組果蠅腦部ROS、MDA、GSH及SOD含量統(tǒng)計;(1)與control組比較,P<0.01;(2)與PD組比較,P<0.05; (3)與control組比較,P<0.001; (4)與control組比較,P<0.05;(5)與對PD組比較,P<0.01。
2.7 α-LA對PD果蠅大腦DA神經(jīng)元的影響
2.7.1果蠅腦內(nèi)TH表達水平 與control組比較,PD組果蠅腦內(nèi)TH水平降低(P<0.05);與PD組相比,LA組果蠅腦內(nèi)TH水平升高(P<0.05)。見圖7。
2.7.2α-LA對DA神經(jīng)元的影響 免疫熒光結(jié)果顯示,PD組果蠅腦內(nèi)PPL1和PPL2 DA神經(jīng)元及總DA數(shù)量明顯低于control組(P<0.01);LA組果蠅腦內(nèi)PPL1和PPL2 DA神經(jīng)元及總DA神經(jīng)元數(shù)量多于PD組(P<0.05)。其他神經(jīng)元簇并無明顯影響(P>0.05)。見圖8。
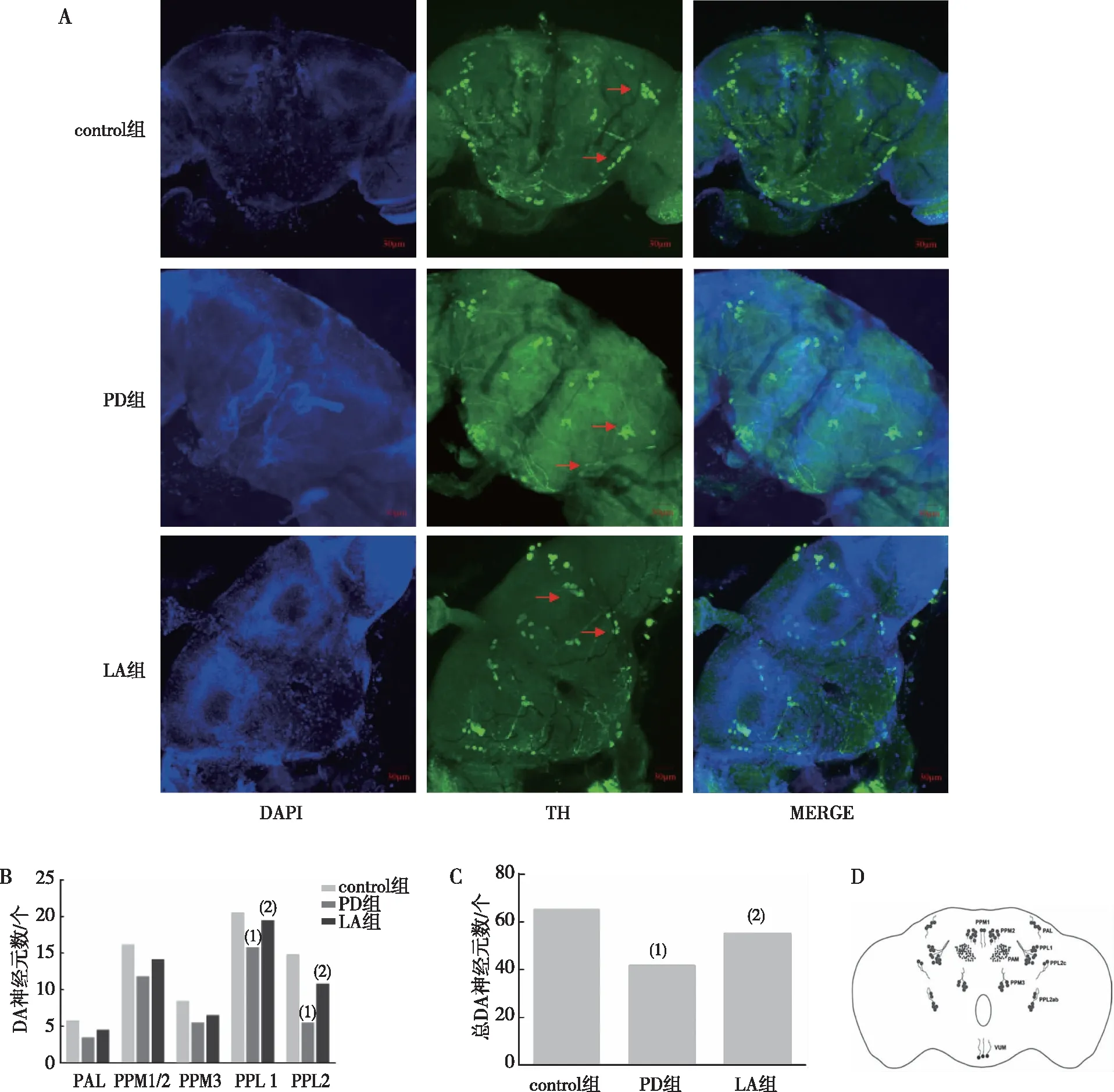
注:A為各組別果蠅腦內(nèi)DA神經(jīng)元分布(20×),紅色箭頭所指為PPL1和PPL2神經(jīng)元簇;B為各組別果蠅腦內(nèi)不同DA神經(jīng)元簇數(shù)量統(tǒng)計;C為不同組果蠅腦內(nèi)總DA神經(jīng)元數(shù)量統(tǒng)計;D為果蠅腦內(nèi)DA神經(jīng)元分布標準圖;(1)與control組比較,P<0.05;(2)與對PD組比較,P<0.05。
2.8 α-LA對PD果蠅腦內(nèi)Atg5及Atg8a的影響
與control比較,PD組果蠅腦內(nèi)Atg5、Atg8a表達水平下降(P<0.05、P<0.01);與PD組比較,LA組果蠅腦內(nèi)Atg5、Atg8a表達水平上升(P<0.05、P<0.05);見圖9A—D。α-LA對Atg5 RNAi或Atg8a RNAi后PD果蠅腦內(nèi)α-syn蛋白的影響,如圖9E—F所示,與PD組比較,LA組果蠅腦內(nèi)α-syn含量下降(P<0.05);與PD組比較,Atg5降低+LA組和Atg8a降低+LA組果蠅腦內(nèi)α-syn含量無明顯變化(P>0.05)。以上結(jié)果提示α-LA可提高PD果蠅腦內(nèi)自噬水平、降低α-LA聚集。
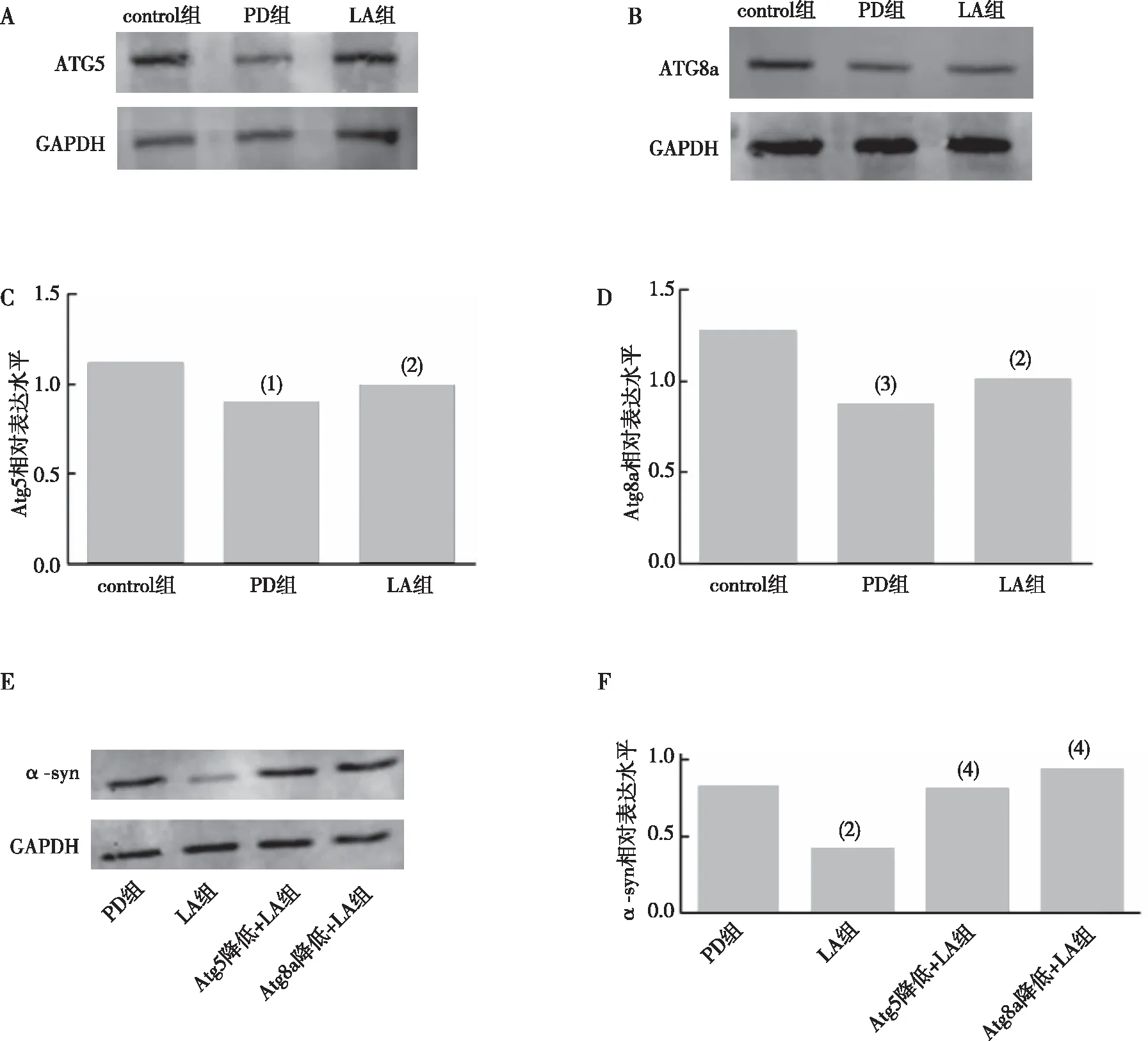
注:A、C為各組果蠅Atg5蛋白的Western blot結(jié)果及相對表達統(tǒng)計;B、D為各組果蠅Atg8a蛋白的Western blot結(jié)果及相對表達統(tǒng)計;E、F為各組果蠅腦內(nèi)α-syn蛋白的Western blot結(jié)果及相對表達統(tǒng)計;(1)與control組比較,P<0.05,(2)與PD組比較,P<0.05;(3)與control組比較,P<0.01;(4)與PD組比較,P>0.05。
3 討論
α-LA具有抗氧化作用,能保護細胞和細胞內(nèi)線粒體不受氧化應激的損傷,并且能夠透過血腦屏障,從而對神經(jīng)元產(chǎn)生保護作用[17]。相關(guān)研究表明,α-LA能夠提高魚藤酮誘導的PD大鼠模型紋狀體DA量以及GSH含量,并且降低脂質(zhì)過氧化和蛋白質(zhì)羰基含量;同時,α-LA能夠改善PD模型大鼠行為學,并且延緩DA神經(jīng)元的丟失[9-11]。這些結(jié)果都證明α-LA能保護PD中多巴胺神經(jīng)元,從而維持多巴胺含量,減緩PD的運動癥狀。PD患者最主要的運動癥狀包括行動遲緩、肌肉僵硬、靜止性震顫、步態(tài)不穩(wěn)等。果蠅是理想的模式生物,其大腦中存在DA能神經(jīng)元簇,DA神經(jīng)遞質(zhì)的合成與人類同源,并且果蠅的多巴神經(jīng)系統(tǒng)同樣參與運動的控制[18-19]。在果蠅DA神經(jīng)元過表達突變的α-syn時,果蠅表現(xiàn)出壽命明顯降低、上爬能力丟失嚴重等類PD表現(xiàn)[20-22];本研究在PD果蠅中通過上爬實驗、繪制生存曲線實驗證明α-LA能夠抑制突變α-syn造成的運動障礙。
α-syn在中樞神經(jīng)系統(tǒng)中高表達,主要定位于神經(jīng)元的細胞質(zhì)和突觸前膜末端。相關(guān)研究表明α-syn聚集會導致線粒體功能障礙[13-16]。過表達α-syn或者α-syn的突變會導致線粒體損傷,線粒體損傷使ROS水平上升,ROS水平升高加劇α-syn聚集,形成惡性循環(huán)[20-21]。在果蠅DA神經(jīng)元過表達突變的α-syn,果蠅同樣表現(xiàn)出腦內(nèi)線粒體損傷、氧化應激水平升高等類癥狀[20-22]。α-LA作為抗氧化劑,具有顯著的抗氧化功能。相關(guān)研究表明α-LA在多種PD模型中發(fā)揮抗氧化作用,保護細胞和線粒體不受ROS的損傷[9-11];同時,α-LA還能夠降低α-syn的水平[22]。本研究通過檢測果蠅腦部α-syn的蛋白含量驗證了α-LA降低α-syn的作用,同時通過對氧化應激標志物ROS、MDA、SOD、GSH等的檢測,證實了α-LA的抗氧化功能,通過ATP水平和NDUFS3表達水平證實α-LA能夠降低α-syn對線粒體的損傷。本研究在轉(zhuǎn)基因PD果蠅中證實了α-LA具有線粒體保護和降低α-syn水平的雙重作用,其機制可能是α-LA抑制了α-syn對線粒體的破壞,從而降低果蠅腦部氧化應激水平,減輕α-syn與氧化應激相互加劇的惡性循環(huán)。α-syn的異常聚集具有神經(jīng)元毒性,DA神經(jīng)元是其主要目標[23]。在過表達α-syn突變的果蠅中,受損的神經(jīng)元也是DA神經(jīng)元[17],與人類PD特征一致。果蠅腦部TH表達水平檢測與免疫熒光染色果蠅大腦DA神經(jīng)元結(jié)果一致,進一步結(jié)果顯示PD果蠅受損的DA神經(jīng)元簇主要是PPL1和PPL2,兩結(jié)果說明α-LA具有神經(jīng)保護作用,能夠降低α-syn對果蠅DA神經(jīng)元的損傷。
有研究發(fā)現(xiàn),PD患者的大腦神經(jīng)元中可檢測到異常溶酶體以及自噬體的存在,且抑制溶酶體形成,α-syn的水平會急劇上升,反之自噬水平的上升會降低α-syn的聚集。而過表達正常或A30P、A53T突變都會抑制自噬的發(fā)生[24-25]。這些結(jié)果表明自噬和α-syn異常聚集具有非常密切的關(guān)系,因此自噬溶酶體通路可能是PD病理學機制以及藥物靶點開發(fā)的一個關(guān)鍵點。果蠅存在與哺乳動物同源的自噬過程以及調(diào)控基因,如Atg13、Atg14、Atg5、Atg8等[26]。本實驗通過檢測α-syn含量,結(jié)果顯示α-LA干預后,PD果蠅腦內(nèi)α-syn含量降低。進一步檢測果蠅自噬溶酶體標志性蛋白Atg5和Atg8a,結(jié)果顯示α-LA干預后Atg5和Atg8a的表達水平上升;利用果蠅雜交分別得到Atg5 RNA i和Atg8a RNAi 果蠅品系,自噬RNAi果蠅品系與PD果蠅雜交得到自噬RNAi PD果蠅,α-LA干預后α-syn含量無變化。以上結(jié)果說明,α-LA可能通過提高自噬水平降低α-syn聚集。
綜上所述,本研究利用SNCA.A30P突變轉(zhuǎn)基因帕金森病果蠅模型證實了,α-LA不僅能夠降低果蠅腦內(nèi)氧化應激水平對神經(jīng)元線粒體的損傷,從而產(chǎn)生保護神經(jīng)保護作用,還能提升果蠅腦內(nèi)自噬水平,加強自噬對突變α-syn的降解,減輕α-syn對DA神經(jīng)元的毒性,最終減少DA神經(jīng)元數(shù)量的損失,改善果蠅行動能力、延長果蠅壽命。本研究結(jié)果對α-LA的臨床應用提供了實驗理論依據(jù)。
猜你喜歡
美與時代·美術(shù)學刊(2022年3期)2022-04-27 01:18:15
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
世界科學技術(shù)-中醫(yī)藥現(xiàn)代化(2020年2期)2020-07-25 02:05:56
人大建設(shè)(2019年12期)2019-05-21 02:55:32
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
西南軍醫(yī)(2016年6期)2016-01-23 02:21:19
西南軍醫(yī)(2015年2期)2015-01-22 09:09:37
食品科學(2013年15期)2013-03-11 18:25:48
