我國不同地區草地貪夜蛾內共生菌感染和耐寒能力差異研究
2023-11-09 12:31:48牛洪濤張志春趙冬曉郭慧芳
環境昆蟲學報 2023年5期
關鍵詞:差異
胡 慧,王 娜,牛洪濤,張志春,趙冬曉,郭慧芳
(江蘇省農業科學院植物保護研究所,南京 210014)
草地貪夜蛾Spodopterafrugiperda是一種新入侵我國的重大害蟲,自2019年1月首次在云南發現以來,迅速擴散至全國大部分地區,給玉米安全生產造成嚴重威脅。遷飛能力強、繁殖力高是草地貪夜蛾快速擴散的重要原因(郭井菲等,2019;齊國君等,2020)。
昆蟲中普遍存在內共生菌,它們在調節宿主生殖和抗逆性等方面發揮著重要作用,是其進化的重要動力(Himleretal., 2011; van den Bosch and Welte, 2016; Lietal., 2020)。蚜蟲、煙粉虱Bemisiatabaci等重要刺吸類害蟲中的內共生菌已受到廣泛關注,除原生共生菌外,發現它們還含有Regiella、Hamiltonella、Arsenophonus、Rickettsia、Serratia和Wolbachia等多種次生共生菌,共生菌的功能各不相同,主要包括提高宿主生殖能力以及抗殺蟲微生物和寄生蜂能力等抗逆功能(Oliveretal., 2005; Himleretal., 2011;ukasiketal., 2013; Liu and Guo, 2019; Lietal., 2020)。此外,還發現共生菌也可削弱宿主的抗逆能力,如非洲粘蟲中Wolbachia能降低宿主對昆蟲核型多角體病毒的抵抗力(Grahametal., 2012),褐飛虱Nilaparvatalugens中共生菌Arsenophonus則降低了宿主對化學藥劑的抵抗力(Pangetal., 2018)。較刺吸類害蟲而言,鱗翅目害蟲體內共生菌研究相對較少。在非洲粘蟲和幻紫斑蛺蝶Hypolimnasbolina等中,發現了Wolbachia的感染(Dysonetal., 2002;Grahametal., 2012)。關于草地貪夜蛾體內微生物,僅有關于腸道微生物的研究,從幼蟲中篩選分離出14種腸道細菌菌株,包括2種纖維素降解細菌(李宏偉等,2020)。該害蟲中內共生菌感染情況如何?國內外尚未見報道。
昆蟲的生長發育和繁殖有著適宜的溫度,溫度過高或者過低都會影響其正常的生命周期,嚴重時導致昆蟲死亡(Broufasetal.,2009)。昆蟲在長期進化過程中為適應惡劣的生境條件,表現耐寒和耐熱能力多樣性,如二化螟Chilosuppressalis不同地理種群耐寒能力有差異,在相同的低溫下,北京種群的半致死時間較福州種群更長(張珺等,2005)。在15℃~30℃范圍內,草地貪夜蛾在15℃下的死亡率最高,不能完成整個世代發育(劉小宇等,2022)。不同地區草地貪夜蛾耐寒能力是否存在差異?其是否與害蟲休內生菌感染存在相關性,均未見研究報道。本文將比較不同地區采集草地貪夜蛾共生菌感染的差異以及耐寒能力差異,以為進一步分析共生菌功能奠定基礎,也為我國草地貪夜蛾遷飛路線分析提供依據。
1 材料與方法
1.1 供試昆蟲
2019年5-7月分別在我國不同地區玉米田采集草地貪夜蛾幼蟲,每個地區采集35~128頭(采集具體信息見表1)。草地貪夜蛾采回后在室內用人工飼料飼養,按地區分別進行群體飼養,飼養條件:溫度27±1℃,相對濕度70%±5%,光周期L∶D=14∶10。飼養1~3代后,在各種群中隨機選用3齡以上幼蟲進行共生菌檢測,待檢測幼蟲單頭分裝到離心管中,保存于-80℃冰箱。
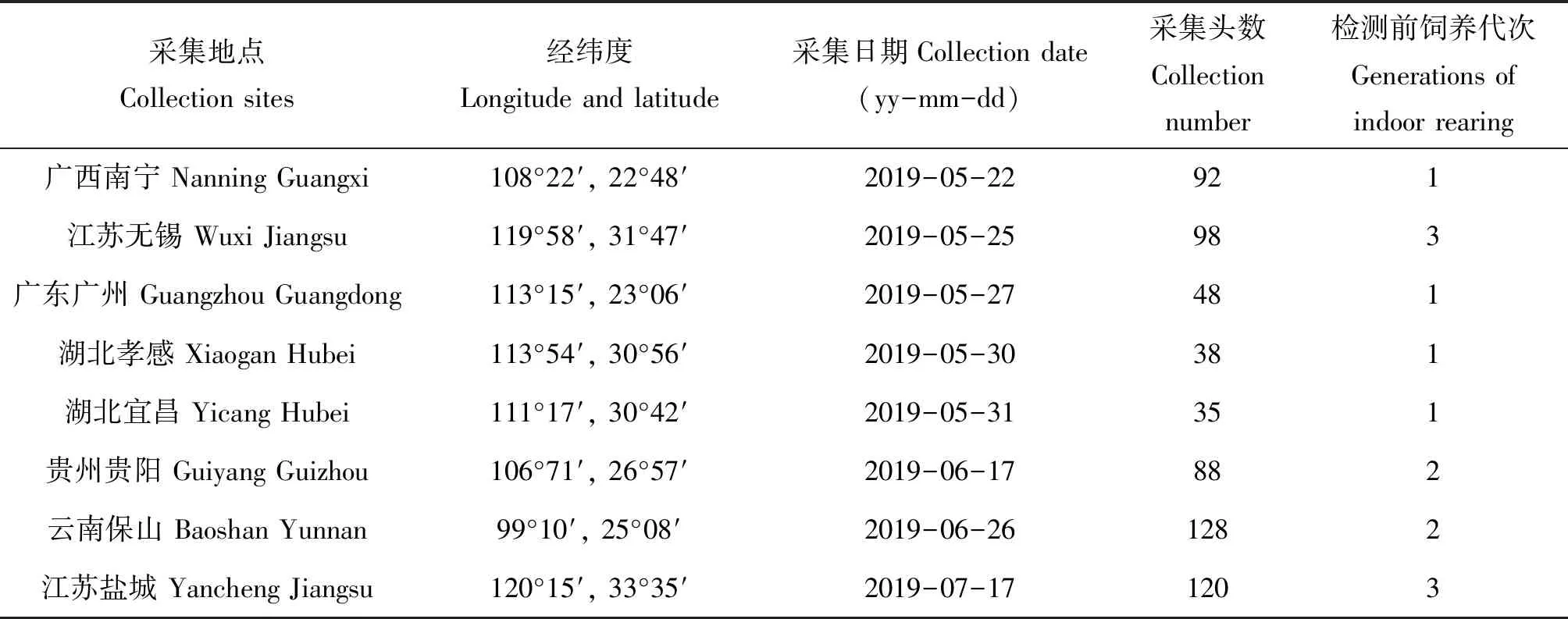
表1 草地貪夜蛾的采集信息
1.2 草地貪夜蛾基因組DNA提取
草地貪夜蛾單頭幼蟲放置于1.5 mL離心管中,先用75%酒精清洗一次,再用無菌水清洗一次,液氮中速凍后研磨至粉狀后,立即加入350 μL PBS溫和研磨30 s,用DNA提取試劑盒(Promega A1125)按說明書進行DNA提取,-20℃冰箱中保存備用。
1.3 草地貪夜蛾夜蛾體內共生菌檢測
除廣西南寧種群收集84頭試蟲外,其余各種群均收集60頭試蟲用于檢測,共檢測Portiera、Hamiltonella、Wolbachia、Cardinium、Arsenophonus、Rickettsia、Fritschea、Serratia和Acinetobacter9種共生菌,引物詳細信息見表2(上海生工生物工程公司合成);PCR擴增反應體系為25 μL,包括 2×Master Mix 13 μL,上游引物和下游引物各1 μL,滅菌超純水8 μL,DNA模板2 μL。擴增程序因共生菌不同而異,共9個程序,分別為:
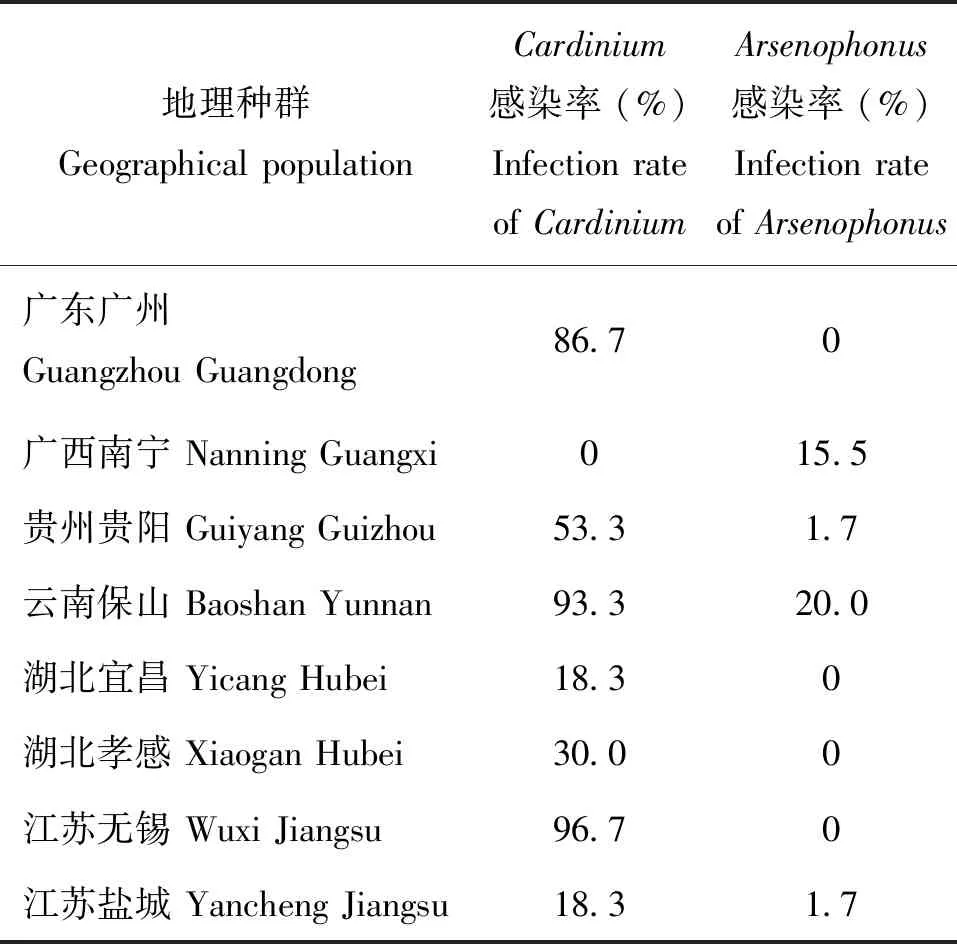
表2 草地貪夜蛾不同地理種群共生菌感染率
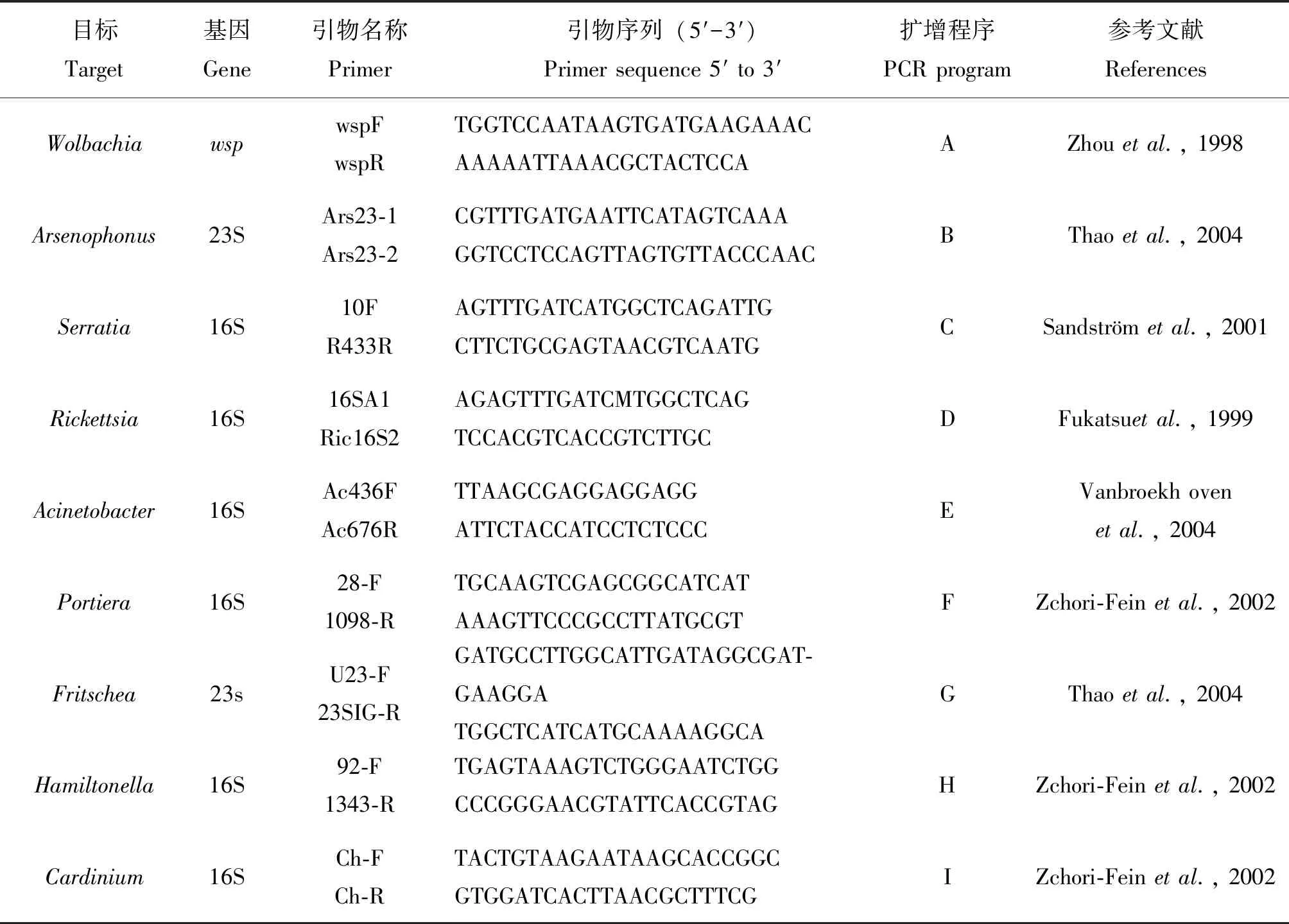
表2 共生菌檢測用引物序列
(A)95℃ 5 min;94℃ 30 s,56℃ 1 min ,72℃ 1 min,循環35次;72℃ 20 min。
(B) 95℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 45 s,循環35次;72℃ 10 min。
(C) 94℃ 2 min;94℃ 1 min,65℃→55℃,1 min,72℃ 2 min,循環10次;94℃ 1 min,55℃ 1 min,72℃ 2 min,循環25次;72℃ 6 min 。
(D) 95℃ 3 min;94℃ 1 min,58℃ 1 min,72℃ 90 s,循環35次;72℃ 5 min。
(E) 95℃ 5 min;95℃ 1 min,58℃ 1 min,72℃ 1 min,循環30次;72℃ 20 min。
(F) 95℃ 5 min;94℃ 1 min,64℃ 1 min,72℃ 90 s,循環35次;72℃ 5 min。
(G) 95℃ 3 min;94℃ 1 min,56℃ 1 min,72℃ 1 min,循環5次;95℃ 1 min,58℃ 1 min,72℃ 1 min,循環30次;72℃ 20 min。
(H) 95℃ 3 min;94℃ 1 min,56℃ 1 min,72℃ 90 s,循環35次;72℃ 10 min。
(I)95℃ 3 min;94℃ 1 min,56℃ 1 min,72℃ 90 s,循環35次;72℃ 10 min。
PCR產物的純化、連接轉化與序列測定:使用普通瓊脂糖凝膠DNA回收試劑盒回收PCR產物(TIANgel Midi Purification Kit),具體步驟參照說明書。連接轉化反應按照pEASY-T3 Cloning Kit試劑盒說明書進行。挑斑、搖菌后,將陽性克隆用于測序,所有測序均由南京金斯瑞生物科技有限公司完成,測序結果在NCBI網站中進行BLAST比對。
1.4 草地貪夜蛾夜蛾耐寒能力測定
通過測定-6℃低溫下草地貪夜蛾的死亡率來比較害蟲的耐寒能力。將經過室內連續20代人工飼養的各地區草地貪夜蛾2齡幼蟲放入裝有人工飼料的培養皿中(直徑35 mm),每個培養皿20頭作為一個重復,每個種群5個重復,放置于人工氣候培養箱中。培養箱溫度從27℃以0.25℃/min速度降至0℃,在0℃保持3 h后,再以0.25℃/min的速度降至-6℃,在-6℃保持5 h后,以0.25℃/min的速度升溫至27℃,在27℃下保持48 h。每隔24 h加入新鮮人工飼料,觀察記錄各培養皿中死亡蟲數,計算死亡率。
1.5 數據處理與分析
應用MEGA 6軟件對草地貪夜蛾幼蟲內共生菌進行同源性分析,鄰接法(Neighbor-Joining(NJ))構建基于16S rDNA序列或23S rDNA序列的系統發育樹。應用SPSS 19.0軟件對不同地區草地貪夜蛾共生菌感染率差異進行χ2檢驗分析,對低溫下害蟲死亡率采用單因素方差分析和Tukey多重比較法進行不同地區間的差異顯著性分析,對共生菌感染率與低溫死亡率進行相關性分析。
2 結果與分析
2.1 我國草地貪夜蛾攜帶共生菌種類
對來自廣西、廣東、湖北、云南、貴州和江蘇等8個地區草地貪夜蛾進行共生菌檢測,共檢測包括Portiera、Cardinium、Arsenophonus、Hamiltonella、Wolbachia、Rickettsia、Fritschea、Acinetobacter、Serratia等9種共生菌。發現各地草地貪夜蛾中只檢測到Cardinium和Arsenophonus這兩種共生菌,Cardinium16S rDNA序列號為MN864662.1,Arsenophonus23S rDNA序列號為MW282126.1。
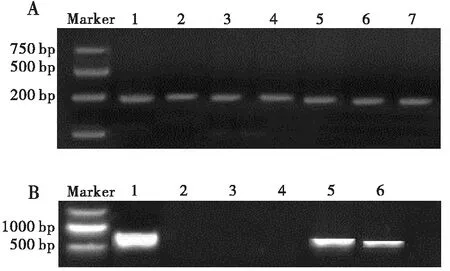
圖1 草地貪夜蛾體內共生細菌擴增產物電泳圖
2.2 草地貪夜蛾體內Arsenophonus和Cardinium的同源性分析
比較分析了草地貪夜蛾與其它昆蟲體內共生菌Arsenophonus的同源性,基于23S rDNA序列通過NJ法構建的系統發育樹顯示(圖2),草地貪夜蛾體內Arsenophonus與同屬鱗翅目的二化螟中Arsenophonus親緣關系最近,在同一分支上,與煙粉虱、麗蠅蛹集金小蜂Nasoniavitripennis、大豆蚜Aphisglycines中Arsenophonus的親緣關系相對較遠,在另一分支上,而與白蠟蚧Ericeruspela和褐飛虱的親緣關系最遠,后兩者在另一獨立分支上。
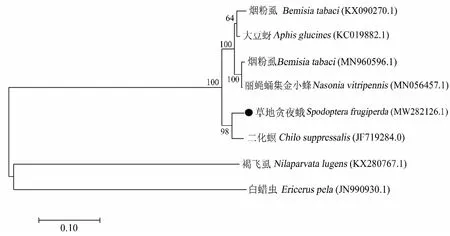
圖2 基于23S rDNA序列構建的草地貪夜蛾共生菌Arsenophonus系統發育樹
對草地貪夜蛾與其它昆蟲體內共生菌Cardinium的同源性進行了分析,基于16S rDNA序列分析構建系統發育樹表明(圖3),草地貪夜蛾體內共生菌Cardinium與果蠅Drosophilabifurca、螫斑鼓翅蠅Sepsispunctum、恩蚜小蜂Encarsiainaron最近,在同一分支上;與煙粉虱體內Cardinium較為接近,而與Hamaliasirokata和白背飛虱Sogatellafurcifera中Cardinium親緣關系相對較遠,與螨Microzetorchestesemeryi中Cardinium的親緣關系最遠。

圖3 基于16S rDNA 序列構建的草地貪夜蛾共生菌Cardinium系統發育樹
2.3 不同地區草地貪夜蛾共生菌感染率差異
對采自江蘇、廣東、廣西、貴州、云南和湖北等6個省份8個地區草地貪夜蛾共生菌感染率進行了比較分析,結果發現,Arsenophonus和Cardinium這兩種共生菌感染率在不同地區間均存在顯著差異(表2)。從Arsenophonus的感染來看,各地理種群中總體較低,以云南保山地區草地貪夜蛾Arsenophonus的感染率最高,為20.0%,其次是廣西南寧地區,感染率為15.5%,貴州貴陽和江蘇鹽城的感染率均為1.7%,而廣東廣州、江蘇無錫以及湖北宜昌和孝感這4個地理種群中均未發現Arsenophonus的感染(χ2=78.658, df=7,P<0.001)。共生菌Cardinium的感染總體較高,感染率最高的為江蘇無錫種群,高達96.7%,其次為云南保山種群,感染率為93.3%,廣東廣州種群感染率為86.7%,貴州貴陽種群為53.3%,湖北孝感和宜昌種群感染率則分別為30.0%和18.3%,江蘇鹽城種群感染率也為18.3%,而廣西南寧種群中未發現有Cardinium感染(χ2=296.727, df=7,P<0.001)。
2.4 不同地區草地貪夜蛾耐寒能力比較
為明確不同地區草地貪夜蛾耐寒能力差異,比較了廣西南寧等8個地區草地貪夜蛾在-6℃低溫脅迫下的死亡率。結果發現,不同地區草地貪夜蛾耐寒能力存在顯著性差異,湖北宜昌地區草地貪夜蛾耐寒能力最強,低溫脅迫后死亡率最低,為35%,顯著低于江蘇鹽城和湖北孝感地區死亡率(76%和69%),其它6個地區害蟲低溫下死亡率也較低,在46%~49%之間,同樣顯著低于鹽城和孝感地區,但6個地區間無顯著差異(F7, 39=4.653,P=0.0011;圖4)。
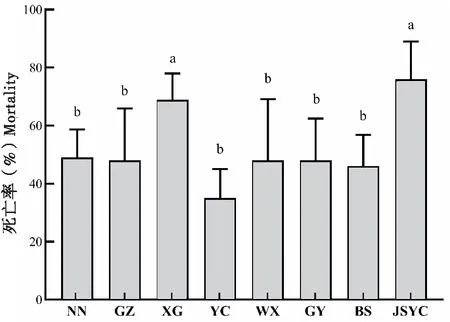
圖4 不同地區草地貪夜蛾-6℃脅迫下的死亡率比較
2.5 草地貪夜蛾共生菌感染與耐寒能力相關性分析
對各地區草地貪夜蛾共生菌感染率和低溫下害蟲死亡率相關性分析表明,Cardinium感染率與-6℃低溫脅迫下的死亡率沒有顯著相關性(線性回歸相關系數=0.3971,P>0.05),Arsenophonus感染率與-6℃低溫脅迫下的死亡率也沒有顯著相關性(線性回歸相關系數=0.5023,P>0.05),說明草地貪夜蛾中Cardinium和Arsenophonus感染與宿主耐寒能力均無直接相關性。
3 結論與討論
本研究首次發現了草地貪夜蛾體內有內共生細菌Cardinium和Arsenophonus的感染,兩種共生菌的感染在我國2019年采集的不同地理種群中存在差異。Cardinium在我國江蘇、云南以及廣東各地草地貪夜蛾中感染率較高,而在廣西南寧種群中未被發現,而Arsenophonus在被檢測種群中有一半未被發現,已發現地區感染率也不高。作為一種新入侵我國的害蟲,草地貪夜蛾一入侵后就表現出了共生菌感染在不同地區間的差異,說明草地貪夜蛾在入侵我國的初期尚未受到本地其它害蟲共生菌的顯著影響,可能與遷入蟲源相關。
關于草地貪夜蛾在我國的遷飛路徑,Li等(2020)利用軌跡分析方法預測了草地貪夜蛾在我國有東線和西線兩條由南向北的遷飛路線,其中東線源于泰國、老撾、越南,進入我國華南地區后,逐步向北遷入長江流域、黃淮以及黃河以北地區;西線則源于緬甸,進入我國云南省后,經貴州、四川后,遷入山西、陜西、甘肅等地。從我國各地草地貪夜蛾中共生菌Cardinium感染情況比較來看,我國廣東廣州、云南保山和江蘇無錫地區害蟲體內Cardinium感染率均很高,而廣西南寧地區未見Cardinium感染,說明江蘇地區草地貪夜蛾來源于廣東的可能性很大,廣西和廣東的境外蟲源可能存在一定的差異。在草地貪夜蛾中發現的另一種共生菌Arsenophonus在云南種群中有一定的感染率,而在廣東以及長江中下游的湖北和江蘇地區均接近零感染。草地貪夜蛾在各地共生菌的差異為用軌跡分析方法預測出來的害蟲東西線兩條遷飛路徑初步提供了一定的現實依據,但同一地區或者模擬同一路徑上害蟲共生菌感染的差異性,也反映出遷飛性害蟲遷飛路徑的復雜性,要進一步通過不同地區草地貪夜蛾共生菌的差異分析來明確害蟲的詳細遷飛路徑,今后還需要對更多地區害蟲共生菌進行具體分析。再從各地區草地貪夜蛾抗寒能力差異來看,尚未表現出與害蟲東西遷飛路線的相關性。
關于共生菌Cardinium在昆蟲中的功能,已報道它主要與宿主生殖調控相關。Cardinium和Wolbachia共感染能誘導煙粉虱雄性后代減少和細胞質不親和,在寄生蜂Encarsiasuzannee中Cardinium也引起宿主昆蟲的細胞質不親和(Lvetal.,2020;Pilgrimetal., 2021)。而就Arsenophnus的功能,已發現其具有調控生殖和影響化學藥劑和殺蟲微生物敏感性等多種功能。已報道Arsenophnus在麗蠅蛹集金小蜂中可殺死雄性胚胎,導致其雌性化(Hugeretal., 1985),在褐飛虱中幫助宿主抗殺蟲真菌綠僵菌(朱歡歡等,2017),還降低了褐飛虱對吡蟲啉的敏感性(Pangetal., 2018)。為發掘共生菌新功能,本研究在比較發現我國不同地區草地貪夜蛾耐寒能力存在顯著差異基礎上,初步分析了Arsenophnus和Cardinium與宿主耐寒能力的相關性,但未發現其存在關聯。Arsenophnus和Cardinium這兩種共生菌在草地貪夜蛾中是否具有抗殺蟲微生物以及調控生殖的功能?除此外,有無其它功能?均有待進一步的研究。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
