蓖麻RcSRS家族鑒定及表達模式分析
2023-11-15 10:42:16徐興源李艷肖朱夢洋向殿軍
種子 2023年8期
關(guān)鍵詞:分析
徐興源, 李艷肖, 朱夢洋, 向殿軍, 劉 鵬
(內(nèi)蒙古民族大學(xué)農(nóng)學(xué)院/內(nèi)蒙古民族大學(xué)科爾沁沙地生態(tài)農(nóng)業(yè)國家民委重點實驗室, 內(nèi)蒙古 通遼 028000)
SRS(SHI-related sequence)基因,也被稱為短節(jié)間(STY)基因,編碼植物特有的一類轉(zhuǎn)錄因子,在應(yīng)對各種刺激信號的特異性反應(yīng)中有著至關(guān)重要的作用。第1個SRS基因是在擬南芥(Arabidopsisthaliana)中發(fā)現(xiàn)的,被命名為LRP1基因[1]。SRS基因擁有2個特殊的結(jié)構(gòu)域:環(huán)形鋅指結(jié)構(gòu)域(C-X2-C-X7-C-X-H-X2-C-X2-C-X7-C-X2-H)[2-3]和IGGH結(jié)構(gòu)域[4]。其中,環(huán)形鋅指結(jié)構(gòu)域是RING結(jié)構(gòu)域(C3H2C3或C3HC4)的一種,保守的RING結(jié)構(gòu)域最早在動物非洲爪蟾(Xenopuslaevis)中被發(fā)現(xiàn),其中有1個DNA結(jié)合基序[5],該結(jié)構(gòu)域能與RNA、蛋白質(zhì)等互作,參與細胞的生理和生化過程[6];而IXGH結(jié)構(gòu)域是同源異構(gòu)化所必需的[7]。除了這兩個保守結(jié)構(gòu)域之外,其余的蛋白質(zhì)是高度分化的[8]。
SRS基因在植物器官的生長[9]、心皮發(fā)育[10]和信號轉(zhuǎn)導(dǎo)[11]等方面均有重要作用。目前,SRS家族已在擬南芥、水稻(OryzasativaL.)、棉花(Gossypiumspp.)和大麥(HordeumvulgareL.)等不同植物中被鑒定和表征。擬南芥中有11個SRS家族成員被報道,其11個成員均擁有環(huán)形鋅指結(jié)構(gòu)域和IGGH結(jié)構(gòu)域,其中側(cè)根原基1(LRP1)會與SHI、SRS3和SRS6等形成復(fù)合體[4,12-14],從而影響側(cè)根和不定根原基的發(fā)育,還能影響染色質(zhì)的修飾和生長素信號傳導(dǎo)[15]。據(jù)報道,擬南芥SRS5基因是光形態(tài)建成的正調(diào)控因子,可直接與光形態(tài)建成基因(如HY5、BBX21和BBX22)的啟動子結(jié)合,激活其表達,促進光形態(tài)建成[15-19]。
SRS家族可參與并影響植物的生物合成。如:擬南芥的STY1和STY2基因可以調(diào)節(jié)生長素的生物合成,并以劑量依賴的方式協(xié)同調(diào)控擬南芥雌蕊、雄蕊和葉片的發(fā)育[8],其中STY1基因主要在發(fā)育的雌蕊中表達[20-21]。另外,STY1基因突變后會造成雌蕊發(fā)育不良,葉片的發(fā)育也會受到影響[8,22]。
SRS家族還參與多種非生物脅迫誘導(dǎo)[23-24]。例如,在紫花苜蓿(Medicagosativa)的莖組織中,有27個MsSRS基因的表達受到冷脅迫和鹽脅迫的誘導(dǎo),呈現(xiàn)出不同的表達模式,表明MsSRSs可能在冷脅迫和鹽環(huán)境下在苜蓿的莖組織中發(fā)揮作用[23]。大豆(Glycinemax)SRS家族的21個成員在干旱、鹽和外源ABA誘導(dǎo)下也表現(xiàn)出不同的表達模式,其中GmSRS18在鹽脅迫下顯著表達[24]。然而,蓖麻(Ricinuscommunis)的SRS家族成員的鑒定和相關(guān)的研究卻鮮有報道。
蓖麻是大戟科蓖麻屬植物,在工業(yè)[25]、醫(yī)藥[26]和航天[27]等諸多領(lǐng)域都有應(yīng)用,在環(huán)境保護和修復(fù)等方面也發(fā)揮著積極作用。蓖麻適應(yīng)環(huán)境能力強,在鹽堿、貧瘠和污染嚴重的環(huán)境下依然能正常生長[28-30],而SRS家族在應(yīng)對低溫、鹽和干旱脅迫的過程中起著重要的作用[23-24]。本研究從蓖麻全基因組數(shù)據(jù)中檢索出蓖麻的SRS基因,并對篩選出的蓖麻SRS基因進行系統(tǒng)的生物信息學(xué)分析。根據(jù)課題組在蓖麻轉(zhuǎn)錄組數(shù)據(jù)中檢索出的RcSRS基因FPKM表達值,分析其在不同環(huán)境脅迫下的表達模式,為后續(xù)深入研究RcSRS基因在蓖麻中的功能提供理論依據(jù)。
1 材料與方法
1.1 蓖麻SRS家族成員篩選與理化性質(zhì)分析
從油料作物數(shù)據(jù)庫(http://oilplants.iflora.cn/)獲取蓖麻的基因組序列文件、蛋白質(zhì)序列文件和基因組注釋文件。在Rice Geneme Annotation Project 數(shù)據(jù)庫(http://rice.uga.edu/)中獲得的日本晴水稻SRS基因家族蛋白質(zhì)序列作為靶標序列進行本地Blastp比對;并使用hmmer軟件,以PF05142模型檢索出含有該結(jié)構(gòu)域的蛋白質(zhì)序列。結(jié)合BLASTP和hmmer的結(jié)果,刪除冗余序列后將獲取的蛋白序列上傳到NCBI-CD SEARCH 和SMART(https://smart.embl-heide-lberg.de/)網(wǎng)站,確保所篩選的蓖麻SRS蛋白質(zhì)序列準確,最終得到5個蓖麻RcSRS家族成員。蓖麻RcSRS家族成員的理化性質(zhì)分析在ExPASy網(wǎng)站 (https://web.expasy.org/)完成。用WoLF PSORT(https://wolfpsort.hgc.jp/)來預(yù)測RcSRS的亞細胞位置。RcSRS蛋白質(zhì)的磷酸化位點預(yù)測在NetPhos3.1 (https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)網(wǎng)站完成。
1.2 蓖麻RcSRS家族成員的高級結(jié)構(gòu)分析
RcSRS蛋白質(zhì)的二級和三級結(jié)構(gòu)預(yù)測分別使用SOPMA (https://npsa-prabi.ibcp.fr/cgibin/npsa_automat.pl? page=npsa_sopma.html)和SWISS-MODEL(https://www.swissmodel.expasy.org/)在線網(wǎng)站完成,再由SAVES v6.0(https://saves.mbi.ucla.edu/)在線工具對三級結(jié)構(gòu)模型進行打分。
1.3 蓖麻RcSRS家族成員系統(tǒng)進化和基因結(jié)構(gòu)分析
使用MEGA11.0軟件對RcSRS蛋白質(zhì)進行進化樹的構(gòu)建。使用MEME(https://meme-suite.org/meme/doc/meme.html)在線軟件預(yù)測RcSRS蛋白質(zhì)的保守基序,最大Motif數(shù)量設(shè)置為10,其他參數(shù)默認。使用TBtools軟件完成基因結(jié)構(gòu)和保守基序的可視化。
1.4 蓖麻RcSRS家族在染色體上的定位和共線性分析
利用TBtools軟件提取出RcSRS家族成員在染色體上的位置信息和共線性關(guān)系信息,最終使用Advanced Circos程序可視化結(jié)果。
1.5 蓖麻RcSRS啟動子的順式作用元件分析
先從蓖麻的基因組中獲取RcSRS基因轉(zhuǎn)錄起始上游1 500 bp的序列,提交至PlantCare(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)預(yù)測RcSRS啟動子順式作用元件的組成。
1.6 蓖麻與日本晴水稻物種間SRS蛋白質(zhì)的聚類分析
把日本晴水稻的OsSRS蛋白質(zhì)序列和蓖麻的RcSRS蛋白質(zhì)序列整合到同一個文件中,然后用MEGA11.0軟件構(gòu)建進化樹,將結(jié)果提交到iTol(https://itol.embl.de)工具中進行美化處理。
1.7 蓖麻RcSRS家族成員的表達模式分析
以蓖麻品種“通篦5號”為試驗材料,選取籽粒飽滿,大小、光澤和斑紋一致的蓖麻籽種子。后續(xù)種子的培養(yǎng),植株的脅迫處理,總RNA的提取,cDNA文庫的構(gòu)建等參照李艷肖等[31]的方法。從RNA-Seq數(shù)據(jù)中篩選出RcSRS基因的脅迫表達量數(shù)據(jù);并下載Brown等[32]蓖麻的RNA-Seq原始數(shù)據(jù),經(jīng)質(zhì)控后篩選出RcSRS基因的組織表達量數(shù)據(jù)。將上述最終得到的脅迫表達量數(shù)據(jù)和組織表達量數(shù)據(jù)提交到TBtools軟件進行熱圖可視化處理。
2 結(jié)果與分析
2.1 蓖麻RcSRS家族成員信息和理化性質(zhì)分析
根據(jù)基因在染色體上的位置,把從蓖麻中篩選到的5個RcSRS家族成員依次命名為RcSRS1、RcSRS2、RcSRS3、RcSRS4、RcSRS5,并對其蛋白質(zhì)進行基本理化性質(zhì)分析(表1)。RcSRS蛋白質(zhì)的氨基酸數(shù)量為199~350個;分子質(zhì)量為22.11~37.87 kD;理論等電點在6.65~9.20之間,除了RcSRS3外均是堿性蛋白質(zhì)(pI>7);脂肪酸介于47.97~61.54之間;除RcSRS1之外,其余均是不穩(wěn)定性蛋白質(zhì);RcSRS蛋白質(zhì)為親水性蛋白質(zhì);RcSRS蛋白質(zhì)的亞細胞預(yù)測結(jié)果顯示,定位于細胞核的有4個(RcSRS2、RcSRS3、RcSRS4、RcSRS5)、定位于細胞膜的有1個(RcSRS1)。
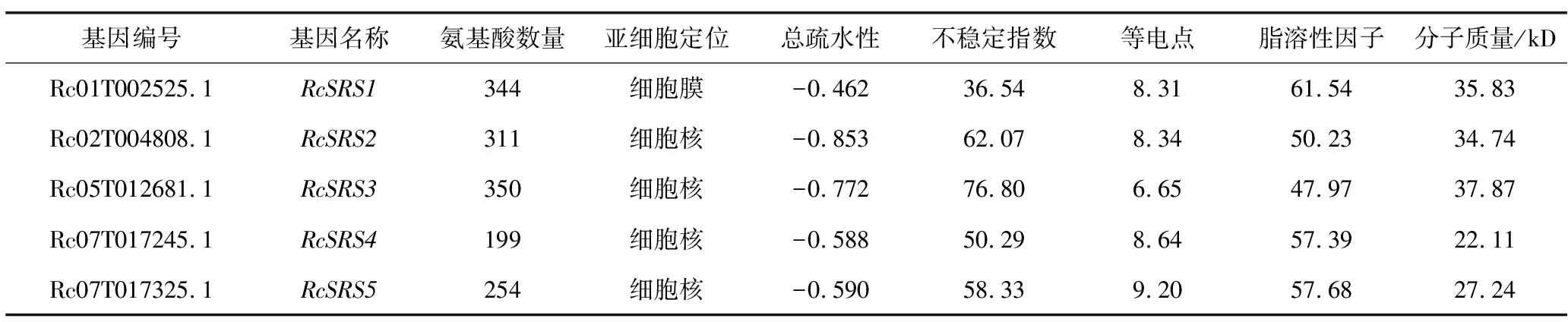
表1 蓖麻RcSRS家族成員的基本信息
RcSRS蛋白質(zhì)的磷酸化位點預(yù)測結(jié)果(圖1)表明,潛在的絲氨酸磷酸化位點數(shù)為12~30個,蘇氨酸磷酸化位點數(shù)目為8~17個,酪氨酸磷酸化位點數(shù)為2~5個。
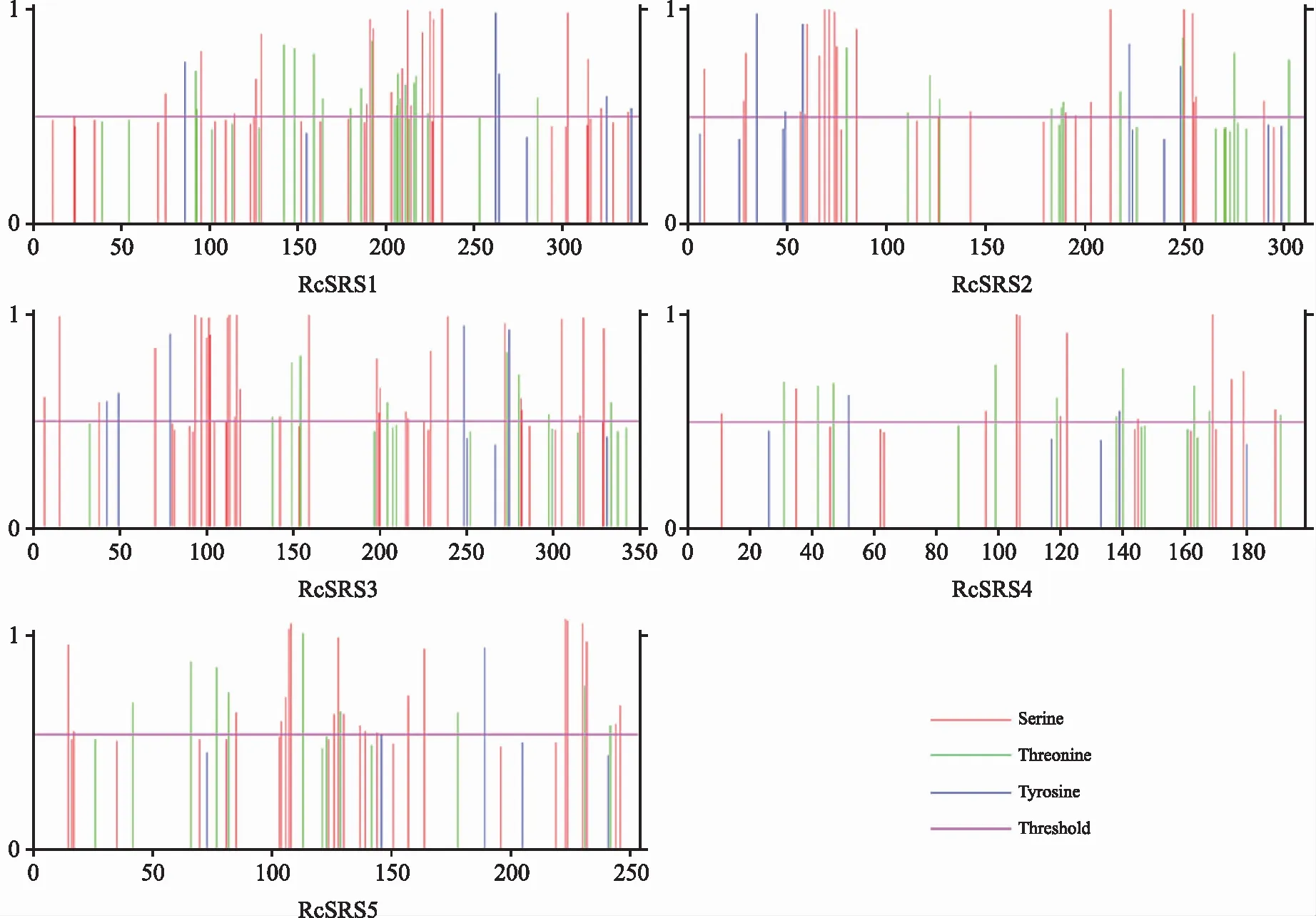
圖1 蓖麻RcSRS蛋白質(zhì)的磷酸化位點
2.2 蓖麻RcSRS家族成員的高級結(jié)構(gòu)分析
RcSRS蛋白質(zhì)的二級和三級結(jié)構(gòu)預(yù)測結(jié)果(表2,圖2)顯示,RcSRS蛋白質(zhì)主要以不規(guī)則卷曲為主,除RcSRS1外,其余RcSRS蛋白質(zhì)均無β折疊結(jié)構(gòu);SAVES v6.0 模型檢測結(jié)果(圖2右)顯示,RcSRS1、RcSRS2和RcSRS3蛋白質(zhì)的氨基酸殘基中有66.7%位于core區(qū)域,22.2%位于次允許區(qū)域;編碼RcSRS4和RcSRS5蛋白的氨基酸殘基中有57.1%位于core區(qū)域,28.6%位于次允許區(qū)域,表明RcSRS蛋白質(zhì)的三級結(jié)構(gòu)模型構(gòu)建結(jié)果合理。
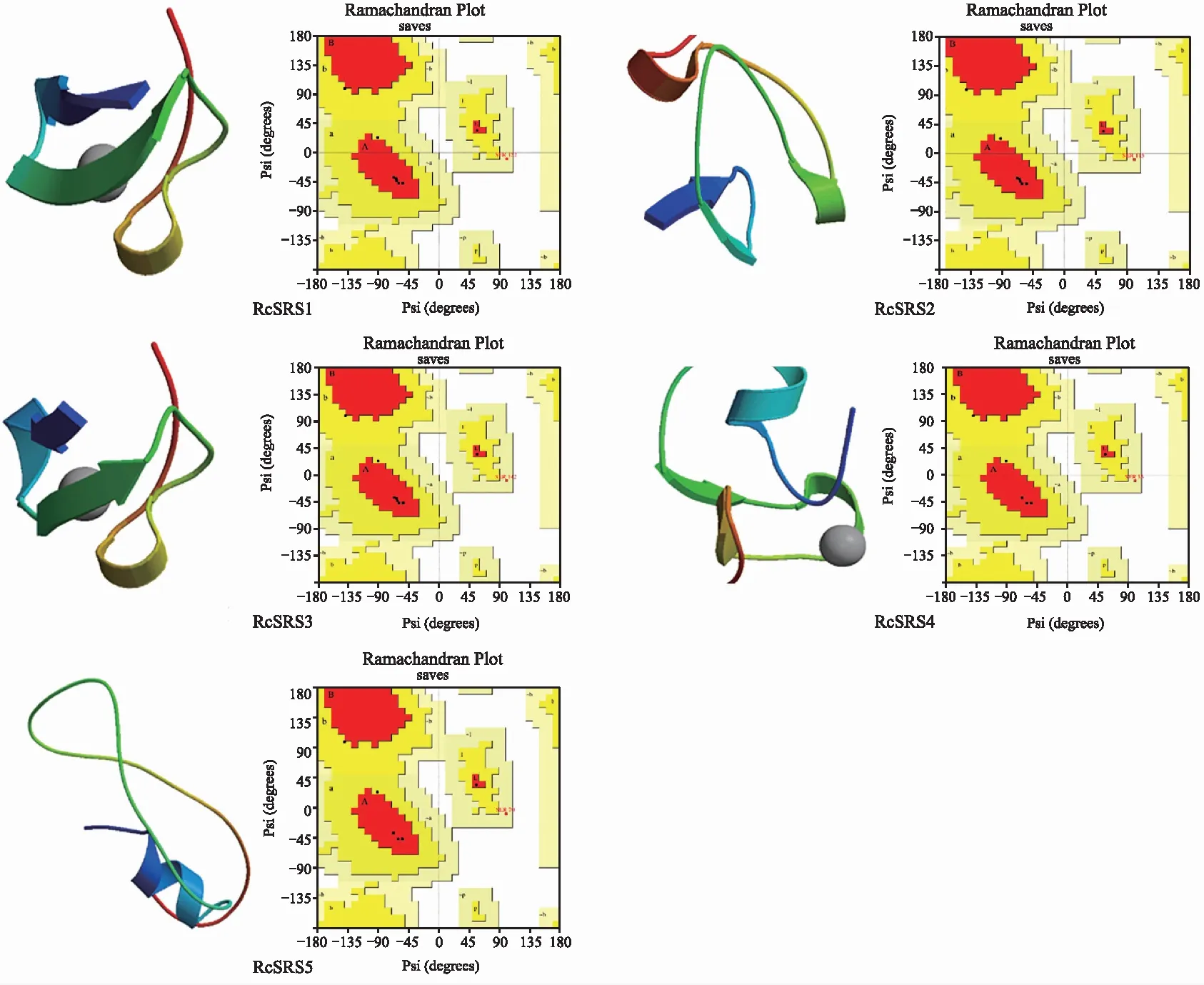
注:左為蛋白質(zhì)的三級結(jié)構(gòu)預(yù)測;右為蛋白質(zhì)的三級結(jié)構(gòu)檢測。

表2 蓖麻RcSRS蛋白質(zhì)的二級結(jié)構(gòu)
2.3 蓖麻RcSRS家族成員系統(tǒng)進化、保守基序和基因結(jié)構(gòu)分析
進化樹結(jié)果(圖3)表明,5個蓖麻RcSRS蛋白被聚為3組。其中RcSRS2和RcSRS3在類群Ⅰ中,RcSRS4在類群Ⅱ中,RcSRS1和RcSRS5則在類群Ⅲ中。保守基序預(yù)測結(jié)果顯示,RcSRS蛋白質(zhì)的保守基序的種類和數(shù)目均不相同,其中RcSRS1和RcSRS5基序數(shù)目最少,均有4個;RcSRS2和RcSRS3的保守基序數(shù)目最多,均有8個。基因結(jié)構(gòu)預(yù)測結(jié)果顯示,RcSRS基因結(jié)構(gòu)較為簡單,僅RcSRS5含有3個外顯子,RcSRS1、RcSRS2、RcSRS3、RcSRS4均僅有2個外顯子,說明RcSRS基因在結(jié)構(gòu)上具有保守性。

圖3 蓖麻RcSRS家族成員系統(tǒng)進化和基因結(jié)構(gòu)分析
2.4 蓖麻RcSRS家族在染色體上的定位和共線性分析
圖4結(jié)果表明,蓖麻RcSRS基因分布在4條染色體上,其中Chr7上擁有兩個RcSRS基因。RcSRS家族成員存有3對共線性關(guān)系,分別是RcSRS2~RcSRS3、RcSRS2~RcSRS5和RcSRS3~RcSRS5,說明RcSRS基因在進化過程中存在基因間的復(fù)制現(xiàn)象。
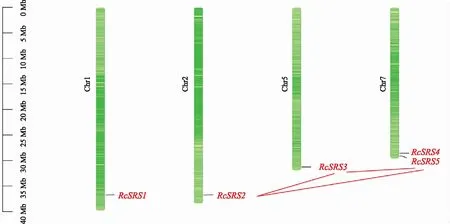
圖4 蓖麻RcSRS家族在染色體上的定位和共線性分析
2.5 蓖麻RcSRS家族順式作用元件分析
對RcSRS啟動子區(qū)域的順式作用元件進行預(yù)測,結(jié)果(圖5)表明,RcSRS擁有多個與逆境脅迫響應(yīng)、防御反應(yīng)和應(yīng)激反應(yīng)等相關(guān)的順式作用元件。其中RcSRS2的順式作用元件數(shù)量和種類都是最多的;光響應(yīng)元件在RcSRS啟動子中數(shù)量最多,說明光在RcSRS基因表達調(diào)控的過程中發(fā)揮著重要作用;RcSRS4除了光響應(yīng)元件外僅有1個厭氧誘導(dǎo)相關(guān)元件,說明不同的RcSRS基因?qū)δ婢趁{迫和激素處理的應(yīng)答模式存有差異;另外,RcSRS2、RcSRS3、RcSRS5啟動子中都含有干旱脅迫響應(yīng)元件,說明它們可能均響應(yīng)干旱脅迫。RcSRS的順式作用元件分析說明,RcSRS家族基因在調(diào)控蓖麻的生長和發(fā)育以及在脅迫響應(yīng)等方面均起著重要的調(diào)控作用。
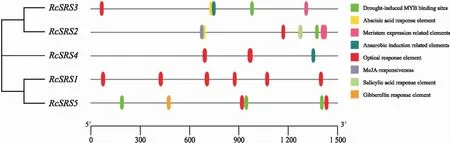
圖5 蓖麻RcSRS家族順式作用元件分析
2.6 蓖麻與日本晴水稻SRS家族成員間的聚類分析
蓖麻(5個)與日本晴水稻SRS(5個)家族成員的聚類結(jié)果(圖6)表明,10個SRS蛋白質(zhì)被聚為三類(類群Ⅰ~類群Ⅲ)。每個亞家族的SRS成員數(shù)不等,RcSRS蛋白質(zhì)只分布于類群Ⅰ和類群Ⅲ中,在類群Ⅲ中分布最多(有3個),分別是RcSRS2、RcSRS3、RcSRS5,在類群Ⅰ中分布最少(只有2個),分別是RcSRS1和RcSRS5。以上結(jié)果表明,RcSRS和OsSRS在進化關(guān)系上比較近,有較高的同源性。
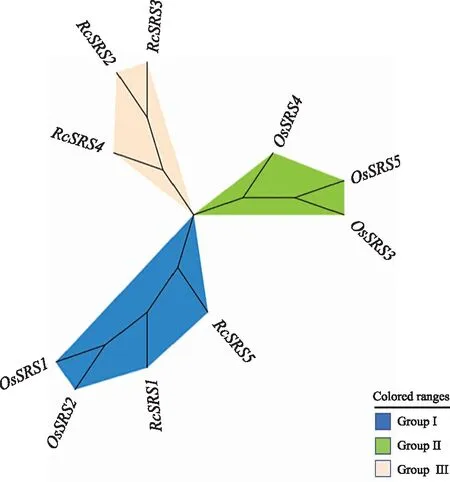
圖6 蓖麻與日本晴水稻SRS家族成員間的聚類分析
2.7 蓖麻RcSRS家族成員的表達模式分析
2.7.1蓖麻RcSRS家族成員在不同組織中的表達模式分析
蓖麻RcSRS基因的表達模式分析(圖7)表明,RcSRS1基因在胚乳、萌芽的種子、葉片中都表達;RcSRS5基因在雄花中表達水平最高,表明RcSRS5基因可能參與并影響雄花的發(fā)育;RcSRS2、RcSRS3、RcSRS4基因在不同組織的表達量都很低或不表達。綜上,RcSRS基因擁有組織表達的特異性。
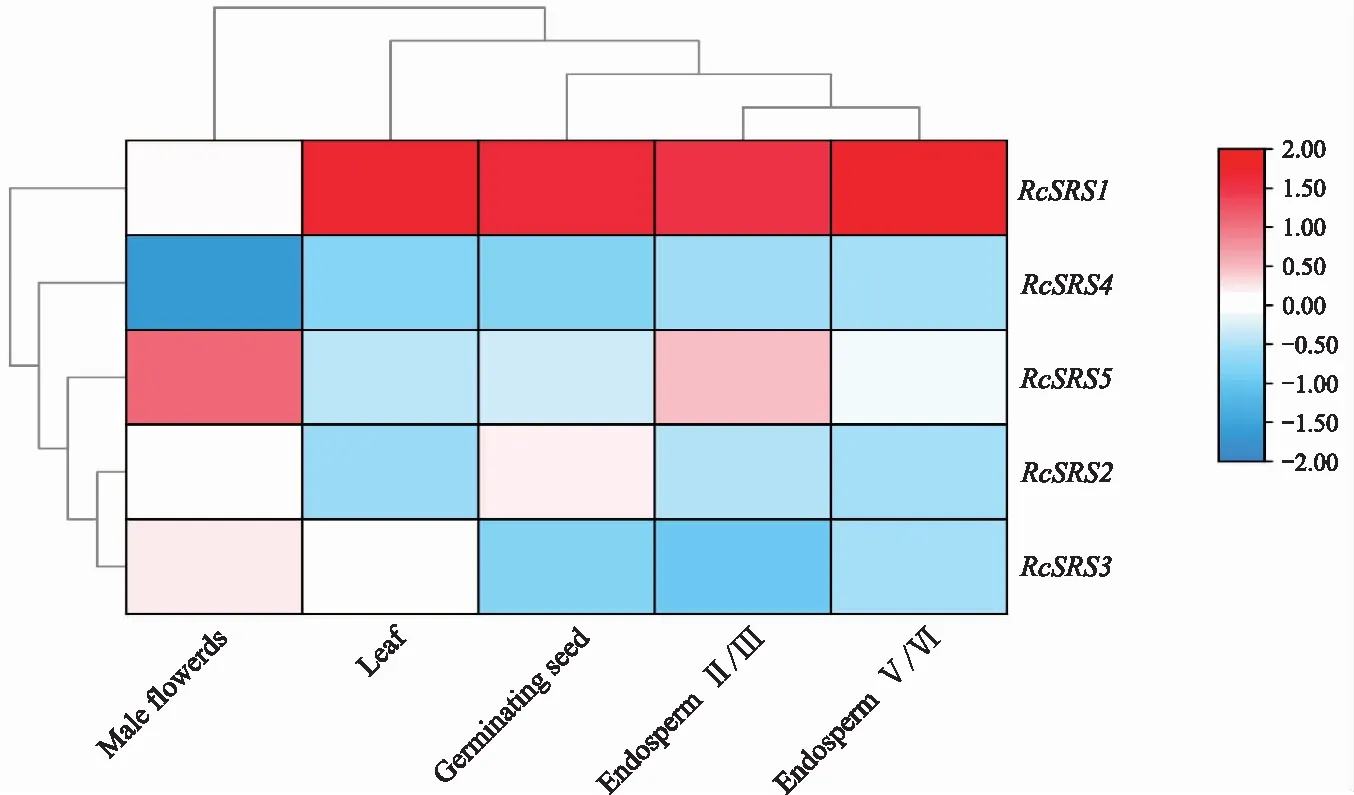
圖7 蓖麻RcSRS基因在不同組織器官中的表達分析
2.7.2蓖麻RcSRS家族成員在非生物脅迫下的表達分析
蓖麻RcSRS家族成員的非生物脅迫表達分析(圖8)顯示,在干旱脅迫和鹽脅迫處理下只有4個基因表達,分別是RcSRS1、RcSRS2、RcSRS4、RcSRS5,其中RcSRS5在干旱脅迫和鹽脅迫處理下12 h和24 h的表達量較0 h處理下呈上調(diào)表達模式。在低溫脅迫處理下只有3個基因表達,分別是RcSRS1、RcSRS2、RcSRS3,他們在低溫脅迫處理下12 h和24 h的表達量較0 h處理下均呈下調(diào)表達模式。綜上,RcSRS基因在蓖麻抵御非生物脅迫過程中具有重要作用。
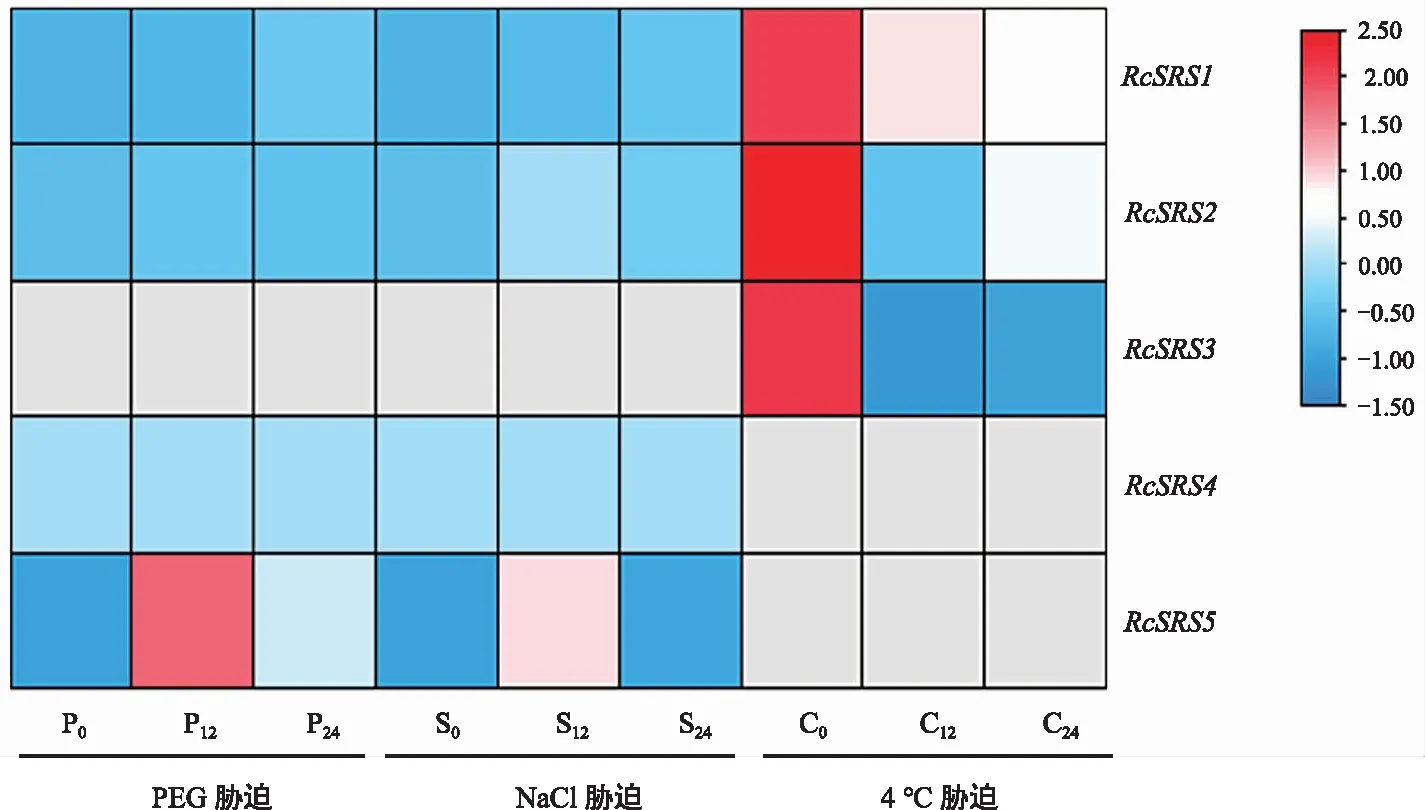
注:0,12和24代表非生物脅迫時長,灰色表示未檢測到。
3 討 論
SRS基因在進化的過程中發(fā)生了變化,不同物種中SRS基因的數(shù)量不盡相同,在擬南芥[2]、蘋果[30]和陸地棉[34-35]中分別含有11,11,27 個成員。本研究在蓖麻中共鑒定到5個RcSRS成員,除RcSRS1之外,其余編碼的蛋白質(zhì)均是不穩(wěn)定性蛋白;不同的RcSRS家族成員所編碼的蛋白質(zhì)理化性質(zhì)均存在差異,可以推測出他們在植物體內(nèi)的功能也不盡相同;對基因結(jié)構(gòu)分析表明,位于同一亞家族的基因家族成員具有相似的基因結(jié)構(gòu),說明同一類群的成員具有相近的進化關(guān)系或功能;對水稻和蓖麻的SRS蛋白質(zhì)進行發(fā)育樹分析,結(jié)果顯示,RcSRS蛋白質(zhì)成員只在類群Ⅰ和類群Ⅱ,表明該家族在進化過程中較為保守,同一聚類中的成員在進化上同源,在結(jié)構(gòu)上表現(xiàn)出相似性。
He等[36]研究發(fā)現(xiàn),SRS家族在葉和花的生長和發(fā)育過程中起著重要的作用。在藜麥(Chenopodiumquinoa)中,除CqSRS08外所有的CqSRSs基因在花中都高表達,CqSRS08則只在葉片中高表達[19]。在擬南芥中,STY1和STY2在花的發(fā)育階段高表達,可以通過調(diào)節(jié)生長素合成的方式來影響雌蕊的發(fā)育,并影響柱頭的形成[8,15]。蓖麻RcSRS家族的不同組織和器官中的表達分析發(fā)現(xiàn),RcSRS家族成員主要在葉和花中高表達,這與張雪等[34]和趙坤宇,陳甜甜[35]在棉花中的研究結(jié)果相一致。在擬南芥中,SRSs已經(jīng)被證明在葉和花的生長發(fā)育中起到了關(guān)鍵的調(diào)控作用[8]。RcSRSs在葉和花中的表達遠高于其他組織,表明RcSRS主要參與了葉和花的生長和發(fā)育。
Ma等[37]研究表明,在植物抵御非生物脅迫的過程中,SRS家族基因同樣起著重要的作用。在蘋果中,11個SRS成員在鹽脅迫、ABA脅迫下均有明顯響應(yīng)[34]。在白香草木樨(Melilotusalbus)中有39個MaSRS基因含有ABRE作用元件,其與植物耐鹽脅迫密切相關(guān),其中MaSRS4和MaSRS32在鹽脅迫下相對表達量均高于未處理的植株,在低溫脅迫下表達量較低,而MaSRS48在低溫下則顯著表達[38]。在蓖麻研究中發(fā)現(xiàn),RcSRS1在低溫脅迫下下調(diào)表達,表明該基因可能在蓖麻抵御低溫脅迫的過程中發(fā)揮作用,可作為后續(xù)研究RcSRS家族基因在蓖麻抵御低溫脅迫作用中的候選基因。分析發(fā)現(xiàn)RcSRS1在啟動子順式作用元件中含有較多的光響應(yīng)元件,其在低溫脅迫下的下調(diào)表達情況可能與之相關(guān),這與李艷肖等[31]的研究結(jié)果相一致。綜上可以看出,RcSRSs基因能響應(yīng)非生物脅迫,但其作用機理尚不明確,還待進一步研究。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
