AICAR對不同腫瘤細胞系c-Myc表達及細胞增殖影響的差異研究
2023-12-12 08:18:34趙艷麗
中國藥理學通報 2023年12期
趙艷麗,尚 曼,王 霆,傅 源
(天津醫科大學基礎醫學院藥理學系,天津 300070)
5-氨基咪唑-4-羧基酰胺核糖核苷(5-aminoimidazole-4-carboxamide 1-β-D-ribofuranoside ,AICAR)是一種細胞內源的代謝物,其結構類似腺苷,通常用作腺苷酸激活蛋白激酶(adenosine 5′ monophosphate-activated protein kinase,AMPK)的激動劑。AMPK在細胞能量代謝調控中發揮核心作用,AICAR則能夠通過激活AMPK信號通路調節細胞代謝, 刺激肌肉纖維活動, 抑制癌細胞生長[1]。AICAR在很多腫瘤學研究中被廣泛使用,許多研究者都通過使用AICAR來證明AMPK參與了的某些特定的生物學過程。作為AMPK的激動劑,AICAR可以抑制細胞內許多消耗ATP的生物過程,因而被認為是一種有前途的抗癌分子。然而,在應用AICAR治療非小細胞肺癌和急性髓系白血病的臨床試驗中,AICAR顯現出嚴重的副作用[2-3]。因此,有必要對AICAR在細胞內的功能展開充分研究,特別是要關注其不依賴于AMPK的生物學功能。
近期很多研究表明,AICAR在細胞內具有不依賴于AMPK的功能。例如,AICAR可以通過誘導Cdc25c降解阻滯細胞周期進程[4];AICAR能夠通過阻止核因子κB(nuclear factor kappa-B, NF-κB)結合于其靶基因啟動子區減弱NF-κB介導的炎癥反應[5]。本研究也揭示了一項AICAR不依賴于AMPK的新功能,即AICAR可以促進原癌基因MYC蛋白(Myc proto-oncogene protein,c-Myc)的表達。因此,在將AICAR作為AMPK激動劑應用于各種臨床場景時,不能簡單地認為AICAR的生物學效應全部是由AMPK激活導致的,有必要對其效應進行全面深入的分析。
c-Myc是一個轉錄因子,在調節癌細胞增殖中發揮著重要作用。在超過70%的人類癌細胞中都可以檢測到c-Myc的異常高表達。大量研究表明,c-Myc是腫瘤發生和發展的重要標志物[1]。既往研究顯示,抑制c-Myc表達是一種有效阻止腫瘤進展的手段[6-7]。作為細胞增殖相關的主要調節因子,c-Myc也會促進一些核苷酸合成相關代謝酶的轉錄[8-9]。因此,有理由認為c-Myc可能會受到一些內源性核苷酸代謝物的反饋調節。然而,到目前為止,還沒有發現任何與核苷酸相關的內源性代謝物可以調節c-Myc。另外,c-Myc也是一個與AMPK有密切聯系的蛋白。兩者雖然屬于相對獨立的信號通路,但都直接調控細胞內能量代謝。兩者之間也存在密切的信號溝通[10-11]。考慮到c-Myc與AMPK的密切聯系以及c-Myc能夠調節核苷酸合成,我們猜想,c-Myc可能也會類似AMPK受某些核酸衍生物的調節。
本研究探討了AICAR對c-Myc表達及細胞增殖的影響。我們發現,AICAR能以細胞特異地方式上調促癌因子c-Myc。這些研究結果完善了我們對AICAR功能的理解,并提示了基于腫瘤細胞異質性個性化地分析AICAR抗癌作用的必要性。
1 材料與方法
1.1 材料
1.1.1細胞 人胰腺癌細胞SW1990、人肝癌細胞HepG2、人乳腺癌細胞MCF-7、人骨肉瘤細胞U2OS來自于國家實驗細胞資源共享平臺。人非小細胞肺癌細胞A549由天津市腫瘤醫院李悅國課題組饋贈。人腎透明細胞腺癌細胞786-O來源于ATCC。
1.1.2主要試劑 AICAR(貨號:HY-13417)和放線菌素D(actinomycin D,貨號:HY-17559)購自MCE公司。放線菌酮(CHX,貨號:14126)來源于Cayman。DAPI(貨號:A606584)購自上海生工生物工程有限公司。TRIzol試劑購自Thermo Fisher(貨號:15596026)。DMEM(貨號:11965092)、RPMI 1640(貨號:11875119)和Opti-MEM(貨號:31985070) 購自Gibco。青霉素-鏈霉素混合液(貨號:P1400)和非必需氨基酸(貨號:N1250)購自北京索萊寶科技有限公司。胎牛血清(貨號:13011-8611)購自四季青。轉染試劑LipofectamineTMRNAiMAX(貨號:13778500)購自Thermo Fisher。兔抗c-Myc購自Proteintech(貨號:10828-1-AP),兔抗AMPKɑ1購自Abcam(貨號:ab32047)。兔抗p-AMPKα Thr172 購自Cell Signaling Technology(貨號:2535S)。β-actin 購自Sigma Aldrich(貨號:A3854)。靶向c-Myc、ADK、ATIC的siRNAs由Synbio technologies公司合成。靶向AMPKα1/2的siRNA購自Santa Cruz Biotechnology(貨號:sc-45312)。
1.1.3主要儀器 生物安全柜購自力康生物醫療科技控股集團。細胞培養箱購自Thermo Fisher Scientific。電泳儀、轉膜儀、普通PCR儀和熒光定量PCR儀購自Bio-Rad。化學發光成像分析儀購自上海天能生命科學有限公司。干式恒溫儀購自江蘇海門市其林貝爾儀器制造有限公司。酶標儀購自Bio-Tek。
1.2 方法
1.2.1藥物的配制 AICAR使用DMSO配制成濃度為1 mol·L-1的母液備用。使用時用培養基稀釋至所需濃度。
1.2.2細胞培養 細胞系SW1990、786-O使用RPMI 1640培養基(含10%胎牛血清、1%青霉素-鏈霉素混合液),HepG2、A549、MCF-7、U2OS使用DMEM培養基(含10%胎牛血清、1%青霉素-鏈霉素混合液和1%非必需氨基酸)。在37 ℃,5% CO2的環境中于細胞培養箱內培養。
1.2.3細胞轉染 取對數生長期的SW1990細胞,接種1×105個細胞至12孔板每孔。待細胞密度達到50%~70%時進行轉染。按照體積比100 ∶2.5 ∶1.5的比例混合Opti-MEM、LipofectamineTMRNAiMAX和siRNA(10 nmol·L-1),并依據說明書所述的操作方法加入至SW1990細胞培基中。轉染時間為48 h。
1.2.4Western blot 向12孔板的細胞中加入100 μL裂解液(含蛋白Laemmli加樣緩沖液)。收集細胞,于98 ℃高溫煮10 min。SDS-PAGE凝膠電泳分離變性蛋白,然后將蛋白轉移至PVDF膜。用含有10%脫脂奶粉的TBST(Tris-buffer saline with Tween-20 ∶20 mmol·L-1Tris, 150 mmol·L-1NaCl, 0.1%w/vTween-20)溶液中封閉2 h。加入一抗后4 ℃過夜孵育。第2天用TBST洗膜,然后室溫孵育二抗2 h。之后再次用TBST溶液洗膜。使用化學發光成像系統對蛋白質進行顯影。
1.2.5qRT-PCR SW1990細胞接種于12孔板中。用AICAR(0,1,2 mmol·L-1)處理細胞16 h。之后收集細胞,用TRIzol試劑提取細胞總RNA。使用Hifair?Ⅱ第一鏈cDNA合成試劑盒(YEASEN)逆轉錄合成cDNA。使用SYBR Master Mix (Yeasen)完成qRT-PCR。引物序列如下:β-actin正向引物序列:5-CGTACCACTGGCATCGTGAT-3,反向引物序列:5-AGGTAGTCAGTCAGGTCCCG-3;c-Myc正向引物序列:5-TCCCTCCACTCGGAAGGAC-3,反向引物序列:5-CTGGTGCATTTTCGGTTGTTG-3。
1.2.6mRNA降解速率檢測 準備兩組細胞,分為對照組和AICAR(2 mmol·L-1)處理組。待AICAR處理完成后,加入放線菌素D(5 mg·L-1)抑制mRNA合成。孵育0、40、60、80 min,100、120 min后收集細胞,提取RNA。使用qRT-PCR的方法檢測mRNA水平。
1.2.7蛋白降解速率檢測 準備細胞,分為對照組和AICAR(2 mmol·L-1)處理組。待AICAR處理完成后,加入放線菌酮(CHX,50 mg·L-1)抑制蛋白合成,孵育0、30、60、90、120、150 min后收集細胞,提取蛋白樣本,使用Western blot方法檢測蛋白水平。
1.2.8MTT檢測細胞活力 SW1990、A549、HepG2細胞分別接種于96孔板內。對照組和實驗組均設置10個復孔,每孔接種8 000個細胞。siRNA的轉染時間為48 h。然后用AICAR(0、2 mmol·L-1)處理細胞16 h。細胞處理完成后,棄去細胞培養基,每孔加入100 μL無血清培養基和10 μL MTT溶液(0.5%)。在37 ℃繼續孵育3~4 h。然后棄去含有MTT的培養基,每孔加入150 μL DMSO。用分光光度法測定了490 nm波長的吸收值變化。
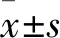
2 結果
2.1 AICAR促進S19990細胞中c-Myc基因表達胰腺癌細胞系SW1990用不同濃度AICAR(0、1、2 mmol·L-1)處理后,c-Myc的蛋白水平升高(Fig 1A)。AICAR(2 mmol·L-1)處理組的c-Myc mRNA水平明顯高于對照組(P<0.01);AICAR(1 mmol·L-1)處理組與對照組間差異無統計學意義(P=0.25;Fig 1B),因此,后續實驗選擇2 mmol·L-1作為AICAR的工作濃度。用AICAR(2 mmol·L-1)處理SW1990細胞0、4、6、8、12、24 h后,c-Myc蛋白水平逐漸升高,在處理約12 h后達到c-Myc蛋白最大表達水平(Fig 1C),因此,確定后續實驗中AICAR處理時間均大于12 h。
2.2 AICAR促進c-Myc基因表達不依賴于AMPKAICAR是一種常用的AMPK激動劑,可以導致AMPK催化亞基于Thr172位點磷酸化(Fig 2A)。為了檢驗其他AMPK激動劑是否也能誘導c-Myc增加,我們用腺苷處理SW1990細胞。結果發現,腺苷抑制c-Myc的表達,與AICAR的作用完全相反 (Fig 2B)。這一結果提示,AICAR促進c-Myc上調可能與其激活AMPK信號通路的功能無關。為了驗證這一假設,我們在SW1990細胞中通過siRNA敲低了AMPK的催化亞基。結果表明,敲低AMPK并不影響AICAR對c-Myc的上調作用(Fig 2C)。

Fig 1 AICAR promoted c-Myc gene expression in SW1990 n=3)

Fig 2 AICAR promoted c-Myc expression in an AMPK-independent manner
2.3 AICAR調節c-Myc表達不依賴于其下游代謝產物在細胞中,AICAR能夠被腺苷激酶(ADK)磷酸化生成5-氨基咪唑-4-羧基酰胺核糖核苷單磷酸(5-aminoimidazole-4-carboxamide-1-β-D-ribofuranosyl 5′-monophosphate, ZMP),并被AICAR轉化酶(ATIC)進一步轉化為次黃嘌呤核苷單磷酸(IMP),腺苷單磷酸(AMP)及鳥苷單磷酸(GMP)(Fig 3A)。為了研究c-Myc表達上調是AICAR的直接效應還是其下游代謝產物的效應,我們在SW1990中敲低了ADK或ATIC (Fig 3B,C)。結果發現這兩個基因的敲低都沒有影響AICAR上調c-Myc的作用,這表明AICAR本身,而不是其下游代謝物上調c-Myc。

Fig 3 The regulatory effect of AICAR on c-Myc expression was not mediated by its downstream metabolites
2.4 AICAR通過促進c-Myc mRNA穩定性上調c-Myc表達為了研究AICAR上調c-Myc表達的機制,我們用RNA合成抑制劑放線菌素D檢測了AICAR對c-Myc mRNA的穩定性的影響,結果發現,AICAR處理后,c-Myc mRNA的穩定性明顯增強(Fig 4A)。然而,使用蛋白質合成抑制劑放線菌酮(CHX)對c-Myc蛋白的穩定性進行測試,結果顯示,AICAR對c-Myc蛋白的穩定性沒有影響(Fig 4B)。這表明,AICAR上調c-Myc基因表達主要是通過促進其mRNA穩定性實現的。

Fig 4 AICAR enhanced stability of c-Myc mRNA n=3)
2.5 AICAR對c-Myc的調節具有細胞選擇性我們選擇了幾種常用的人類癌細胞系進行測試,分別用AICAR(0、1、2 mmol·L-1)處理16 h。結果發現,AICAR在腎透明癌細胞786-O和非小肺癌細胞A549中上調c-Myc蛋白表達(Fig 5A);相比之下,在肝細胞癌細胞HepG2、乳腺癌細胞MCF-7和骨肉瘤細胞U2OS中AICAR下調c-Myc蛋白表達(Fig 5B)。
鑒于AICAR對c-Myc的調控作用具有細胞選擇性,我們選取HepG2作為AICAR不能上調的c-Myc的細胞系代表,測試了AICAR是否會影響這類細胞中c-Myc的mRNA的穩定性。(Fig 5C)結果表明,AICAR不會影響HepG2細胞內c-Myc的mRNA的穩定性。

Fig 6 Effect of AICAR on cell viability
2.6 AICAR對細胞活力的影響SW1990、A549和HepG2細胞分別用AICAR(0、2 mmol·L-1)處理16 h。MTT法測定細胞活力表明,在HepG2細胞中,AICAR(2 mmol·L-1)處理組細胞活力明顯低于對照組(P<0.01;Fig 6C),這與AICAR下調HepG2細胞中c-Myc表達可能引起的預期效應一致。然而,在SW1990細胞和A549細胞中AICAR(2 mmol·L-1)處理組的細胞活力與對照組之間無統計學差異(Fig 6A,B)。在較高濃度(4 mmol·L-1)下,AICAR會略微抑制A549細胞的增殖;而在較低濃度(1 mmol·L-1)下,AICAR會略微促進SW1990細胞的增殖。c-Myc是經典的促細胞增殖基因,AICAR雖然能夠上調c-Myc的表達,但在某些濃度劑量下,AICAR不能上調細胞的增殖速率甚至會下調細胞的增殖速率。這可能是由于AICAR除了影響c-Myc還會影響AMPK等其他信號通路。事實上,AICAR能夠通過激活AMPK抑制細胞增殖,在不同的濃度條件下,不同的信號通路可能會占主導。因此,在多種信號通路的共同作用下,特別是由于c-Myc和AMPK相互拮抗,細胞的表觀增殖速率可能不會發生明顯變化。
鑒于c-Myc在SW1990和A549細胞中被AICAR上調,我們推測抑制c-Myc與AICAR處理聯合治療可能成為抑制這類細胞增殖的有效策略。為了驗證這一猜想,我們用RNA干擾敲低了SW1990、A549和HepG2細胞中的c-Myc,再用AICAR(0、2 mmol·L-1)處理細胞16 h(Fig 6D-H)。結果顯示,在SW1990和A549細胞中,AICAR(2 mmol·L-1)處理組的細胞活力與對照組細胞活力差異無統計學意義,表明單獨AICAR處理不能抑制這兩種細胞的增殖。然而,在敲低c-Myc后,SW1990細胞和A549細胞中AICAR(2 mmol·L-1)處理組的細胞活力明顯低于對照組,差異有統計學意義(P值分別為<0.01和0.014;Fig 6D,E)。表明在AICAR上調c-Myc的細胞中,必須同時抑制c-Myc的表達,AICAR才能有效抑制細胞增殖。相比之下,在HepG2細胞中,當c-Myc正常表達時,AICAR(2 mmol·L-1)處理組的細胞活力低于對照組,差異具有統計學意義(P<0.01;Fig 6F)。在此基礎上敲低c-Myc,AICAR(2 mmol·L-1)處理組的細胞活力依然低于對照組,差異仍具有統計學意義(P<0.01;Fig 6F)。綜上,對于AICAR上調c-Myc表達的細胞,抑制或敲低c-Myc是AICAR發揮抗增殖作用的前提條件;而對AICAR不能上調c-Myc的細胞,無需抑制或敲低c-Myc,AICAR即可發揮抗增殖作用。
3 討論
AICAR是一個經典的AMPK激動劑,AMPK是調控細胞內各種生物能量代謝的關鍵因子。既往研究表明,AICAR能夠通過激活AMPK抑制腫瘤細胞增殖或誘導癌細胞凋亡[12-14]。因此,在大多數研究AICAR的文章中,AICAR的生物學效應往往與AMPK相互關聯。然而,在本研究中,我們發現了AICAR在胰腺癌細胞SW1990中的新作用。AICAR能夠以一種不依賴于AMPK的方式上調轉錄因子c-Myc的表達。AICAR對c-Myc的上調作用不局限于胰腺癌細胞SW1990。通過測試一系列腫瘤細胞系,我們發現AICAR同樣能夠上調腎透明癌細胞786-O和非小細胞肺癌細胞A549中的c-Myc表達。
有趣的是,在肝癌細胞HepG2、乳腺癌細胞MCF-7和骨肉瘤細胞U2OS中,AICAR會下調c-Myc表達,表明AICAR對c-Myc的調控效應具有細胞特異性。
c-Myc是一個在腫瘤發生發展中起重要作用的轉錄因子。在幾乎所有的腫瘤組織中,c-Myc都呈現高表達[1, 6]。c-Myc能夠促進腫瘤細胞的增殖,直接抑制c-Myc可以快速消退小鼠的腫瘤[6]。既往研究表明,AICAR能夠以多種方式發揮抗癌作用。一方面,AICAR是AMPK激動劑,通過激活AMPK進而下調mTOR信號通路抑制細胞內蛋白質合成[15-16]。另一方面,AICAR可以通過抑制核苷酸合成和阻滯細胞周期起到抗腫瘤細胞增殖的作用[17]。但是,本研究顯示,AICAR能夠在某些腫瘤細胞內上調c-Myc的表達水平。由于c-Myc本身是一個促癌因子,因此可以預見,c-Myc表達水平的上調會削弱AICAR的抗癌作用。在某些類型的腫瘤細胞中,AICAR甚至有可能通過c-Myc依賴的方式起到促癌的作用。
MTT實驗表明,AICAR會明顯降低HepG2細胞的增殖速率,而對SW1990和A549的細胞增殖無明顯影響。巧合的是,AICAR會下調HepG2細胞中c-Myc的表達,而上調SW1990和A549細胞中c-Myc的表達。這些現象提示,AICAR對細胞增殖的影響與其對c-Myc的調控作用有關。進一步研究表明,敲低SW1990和A549細胞的c-Myc能夠促使AICAR發揮抑制細胞增殖的效應。因此,我們推測:在SW1990和A549中,AICAR介導了c-Myc的上調,c-Myc的上調抵消了AICAR通過激活AMPK信號通路所帶來的抗增殖效應。而在HepG2細胞中,由于AICAR無法上調c-Myc,所以AMPK信號通路帶來的抗增殖效應占據主導地位。
基于本研究所嘗試的幾種細胞系,AICAR調控c-Myc的作用方向與腫瘤細胞的組織來源無明確關聯。事實上,由于腫瘤細胞具有高度異質性,即便是相同組織來源的腫瘤細胞,由于所攜帶的基因變異不同,其表達譜系也可能會有很大差別[18]。AICAR在哪些腫瘤細胞會上調c-Myc取決于AICAR的直接作用靶點的表達水平。然而,本研究的一個局限是我們并沒有鑒定出AICAR上調c-Myc的直接作用靶點。AICAR結合靶點的鑒定及AICAR上調c-Myc具體的分子機制,還有待進一步探索。
綜上所述,AICAR能夠選擇性地在某些類型的腫瘤細胞內以AMPK非依賴的方式上調c-Myc表達,進而影響細胞增殖。這些實驗結果表明,雖然AICAR是一個很有應用前景的抗癌小分子,但在將其應用于抗癌治療時,需考慮到其對c-Myc等促癌轉錄因子的調控作用。應用AICAR前,需要有針對性地具體分析AICAR對特定類型細胞帶來的特定效應。