Hsp90 在豬繁殖組織的表達特性與定位研究
2024-01-14 10:24:56陳麗圓楊鈺浩
韶關學院學報 2023年12期
關鍵詞:檢測
陳 云,李 晨,陳麗圓,鄧 晶,楊鈺浩
(1.韶關學院 生物與農業學院,廣東 韶關 512005;2. 韶關學院 廣東省粵北食藥資源利用與保護重點實驗室,廣東 韶關 512005)
Hsp90 即熱休克蛋白90,是熱休克蛋白家族的成員,它廣泛參與細胞的激素信號通路形成、細胞周期調控以及細胞發育等重要的生理過程,在雄性生殖過程中精子方面的變化以及分子層面的機制具有重要的意義[1]. 本課題組前期在高、低繁殖力公豬的精液iTRAQ 測序中,發現精子中Hsp90 蛋白的表達量,同精子膜的穩定性、與透明帶的結合密切相關[2]. 高、低繁殖力荷斯坦公牛的精子和精清蛋白質組學測序中發現高繁殖力公牛的精子中Hsp90 的豐度更高,Hsp90 表達量與公牛繁殖力相關[3]. 研究發現,Hsp90 的表達與豬精子品質有著很大關系,影響豬精子的活力、質膜完整率和存活率等[4-5],可能參與豬胚胎的發育過程[6]. Hsp90 可在多個物種的精子鞭毛中檢測到,調節激酶活性、減數分裂和精子成熟,Hsp90 的下調可能造成精子缺陷,從而導致生殖異常和不育[7]. Hsp90 蛋白在豬精液冷凍保存方面發揮重要作用,在豬冷凍精液品質相關基因多態性的研究中,發現Hsp90 基因是與豬冷凍精液品種相關的標記基因[8]. 在牛精液和雞精液中,均發現精子抗凍能力與Hsp90 基因的表達量相關,且Hsp90 基因的表達量可作為牛精子抗凍性的判斷標準[9-10]. 目前,Hsp90 蛋白在豬精液冷凍保存和胚胎發育方面的研究,特別是有關精子活力與基因、蛋白間的理論研究比較匱乏,對它們的保護機理尚未通曉. 因此,本研究分析Hsp90 在豬繁殖組織、生殖細胞中的定位及表達,為豬精液冷凍添加劑的研究提供依據.
1 材料及方法
1.1 試驗材料
1.樣品:母豬卵巢和輸卵管采于廣州市某屠宰場;公豬的組織樣品、精液采于廣東省某種豬場.
2.主要試劑:AlexaFluor 568 羊抗兔二抗購于Thermo 公司;GAPDH 鼠單克隆抗體、Hsp90 兔多克隆抗體購于Abcam 公司;HRP 標記羊抗IgG 抗體、HRP 標記羊抗鼠IgG 抗體購于Earthox 公司;核酸清除劑購自上海澤葉生物科技有限公司;動物組織RNA 提取試劑盒購自天根生化科技有限公司;2×Taq PCR Master Mix 和反轉錄試劑盒購自翌圣生物科技有限公司;Western blot 檢測中用到的試劑均購自北京鼎國昌盛生物技術有限公司;免疫組化檢測用試劑均購自北京雷根生物技術有限公司;全蛋白提取試劑盒和BCA 蛋白含量檢測試劑盒購自凱基生物有限公司;PBS、Hoechst 33342、礦物油和透明質酸酶購自Sigma公司.
1.2 方法
1.2.1 RNA 的提取
取豬的卵巢、輸卵管、睪丸、尿道球腺、前列腺、精囊腺、附睪頭、附睪體、附睪尾等新鮮組織分別加入液氮迅速研磨后,根據RNA 抽提試劑盒說明書用提取各組織總RNA. 用超微量核酸測定儀對其濃度和純度進行測定,用1%瓊脂糖凝膠對其完整性進行檢測,根據反轉錄試劑盒的操作說明合成cDNA.
1.2.2 引物設計與合成
根據NCBI 官網提供的豬源Hsp90 基因和內參基因GAPDH的mRNA 序列,利用Oligo 軟件設計引物(表1),引物在上海生工生物公司合成[11].
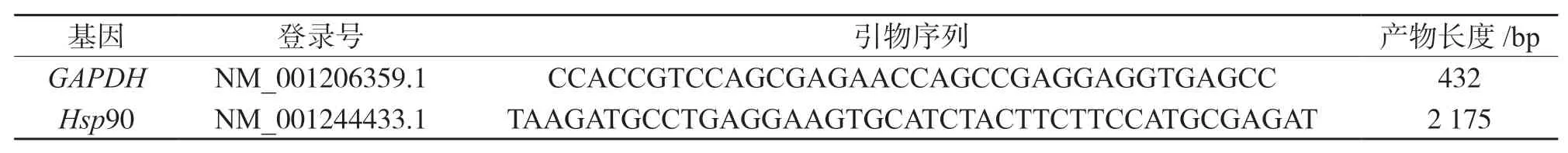
表1 RT-PCR 引物列表
1.2.3 RT-PCR 檢測
RT-PCR 擴增體系(20 μL) :上下游引物(10 μmol·L-1)各0.4 μL,2×Taq PCR Mix 10 μL,模板1 μL,加ddH2O 至20 μL. 擴增采用三步法,即94 ℃30 s,94 ℃15 s,58 ℃2 min,72 ℃10 s,共30 個循環.
1.2.4 免疫印跡蛋白檢測
取10~20 mg 動物組織,參考說明書提取組織蛋白,再參考BCA 蛋白含量檢測試劑盒說明書檢測總蛋白含量. 將組織蛋白按比例加入5×SDS 上樣緩沖液沸水煮10 min 變性,在12%SDS-PAGE 凝膠上上樣20 μg·孔-1,80 V 電泳3 h;將蛋白轉至PVDF 膜,90 V 轉膜1 h,脫脂奶粉室溫封閉1 h;Hsp90 一抗孵育(1∶1 000 稀釋),4 ℃過夜,TBST 洗膜2 次,TBS 洗膜1 次;二抗孵育,37 ℃搖床孵育2 h,洗膜同上;ECL 超敏發光液顯色,曝光顯影. 以內參GAPDH 進行目的蛋白表達量校正[12].
1.2.5 免疫組化檢測
獲得成年公豬和母豬的新鮮樣本,立即固定在10%的福爾馬林中,常規石蠟包埋、切片,厚度為4 μm.二甲苯脫蠟,酒精梯度脫水,抗原修復[12]. 添加兔源Hsp90 一抗(1∶500 稀釋),4 ℃過夜,陰性對照為兔血清. 加HRP 標記的羊抗鼠二抗,37 ℃下孵育1 h,PBS 洗滌,DAB 顯色,終止[12]. 蘇木精復染細胞核,脫水封片,顯微鏡下觀察切片顏色,陽性為棕黃色,自來水沖洗切片終止顯色[12].
1.2.6 精子免疫熒光檢測
取1 mL 精液,用PBS 洗滌3 次. 將50μL 精子懸浮液放在載玻片上,干燥60 min. 在室溫下用3.7%甲醛固定20 min,PBS 洗滌,用10%正常兔血清封閉1 h,然后與稀釋的Hsp90 抗體(82 μg·mL-1)或正常兔IgG 抗體(陰性對照,1 μg·mL-1)共同孵育,4 ℃過夜. PBS 洗滌,與熒光素偶聯的山羊抗兔IgG(1∶1 000 稀釋)在37 ℃孵育1 h,然后與Hoechst 33342(1∶100 稀釋)避光室溫孵育10 min. 通過激光掃描共聚焦顯微鏡檢測Hsp90 蛋白.
1.2.7 卵母細胞免疫熒光檢測
采集成年母豬的卵巢,抽取卵泡液,PBS 沉降3 次后,將卵母細胞的卵丘復合物用透明質酸酶去除,進行免疫熒光染色. 將卵母細胞吸入3.7%甲醛固定,37 ℃培養箱固定30 min 以上;PBS 洗滌3 次,37 ℃培養箱中放置2 h;一抗孵育,將卵母細胞放入提前做好的液滴中,37 ℃培養箱孵育4 h,清洗3 次;二抗孵育,步驟同上,室溫孵育1 h,清洗卵母細胞;將Hoechest 33342 進行稀釋(1∶100),在載玻片上做個液滴,放入卵母細胞,室溫避光放置10 min. 封片,激光掃描共聚焦顯微鏡觀察和拍照.
1.2.8 軟件分析
RT-PCR 結果和Western blot 結果使用Image J 圖像分析軟件測定,采用SPSS 19.0 軟件對數據進行統計和顯著性分析,結果用“平均值±標準誤差”表示,*表示差異顯著,0.01≤P<0.05,**表示差異極顯著,P<0.01.
2 結果
2.1 Hsp90 基因在大白豬繁殖組織中的表達
利用RT-PCR 方法,以GAPDH基因為內參,檢測豬繁殖組織Hsp90 基因的表達,結果顯示,Hsp90基因在睪丸和附睪尾表達,在卵巢中微量表達,見圖1.
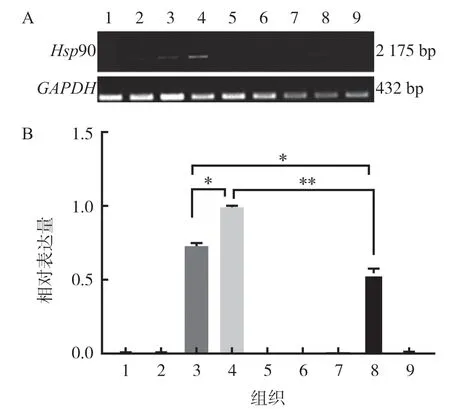
圖1 Hsp90 mRNA 在豬繁殖組織中的表達
2.2 Hsp90 蛋白在大白豬繁殖組織中的表達
利用Western blot 方法,以GAPDH 為內參,檢測豬繁殖組織Hsp90 蛋白的表達,見圖2,結果表明,Hsp90 蛋白在睪丸、卵巢、輸卵管和精囊腺中表達[12].
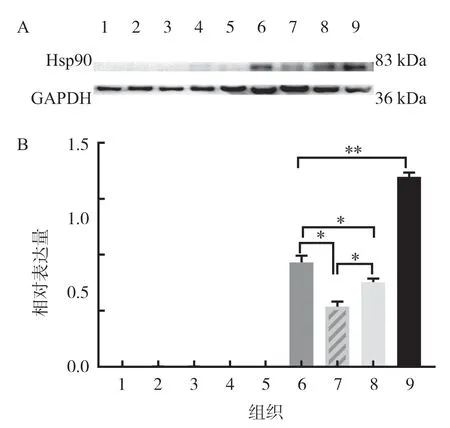
圖2 Hsp90 蛋白在豬繁殖組織中的表達
2.3 Hsp90 蛋白的免疫組化
取新鮮的睪丸和卵巢組織,制作切片. 如圖3,通過免疫組化方法,發現Hsp90 蛋白在睪丸的次級精母細胞、精子細胞和間質細胞中表達(圖3B),也在卵巢的顆粒細胞、卵丘細胞和卵母細胞中表達(圖3D).
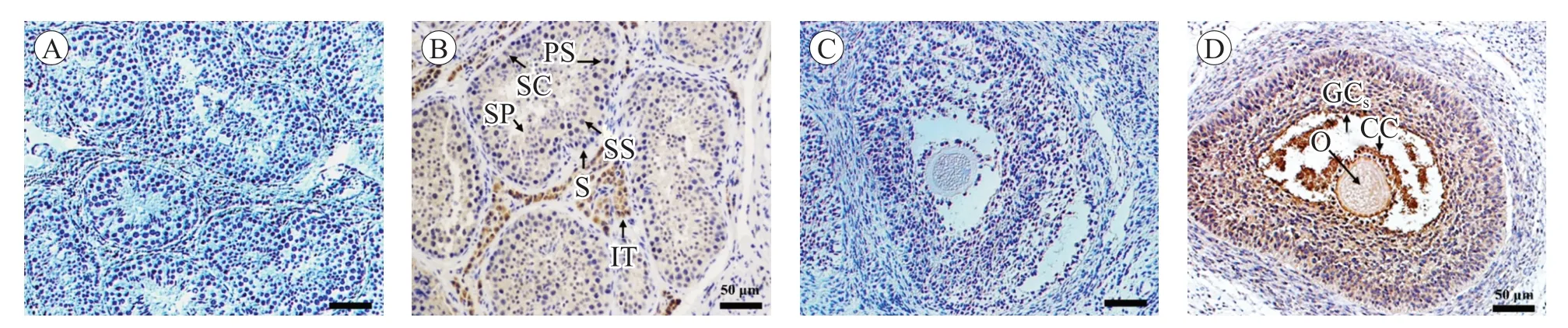
圖3 Hsp90 蛋白在豬睪丸和卵巢中的定位
2.4 Hsp90 蛋白在豬精子和卵母細胞中的表達
使用免疫熒光染色法,檢測Hsp90 在精子和卵母細胞復合體中的表達. 如圖4 及圖5,Hsp90 在精子的頸部和尾巴中部表達(圖4),也在卵丘細胞的細胞質和卵母細胞中表達(圖5).
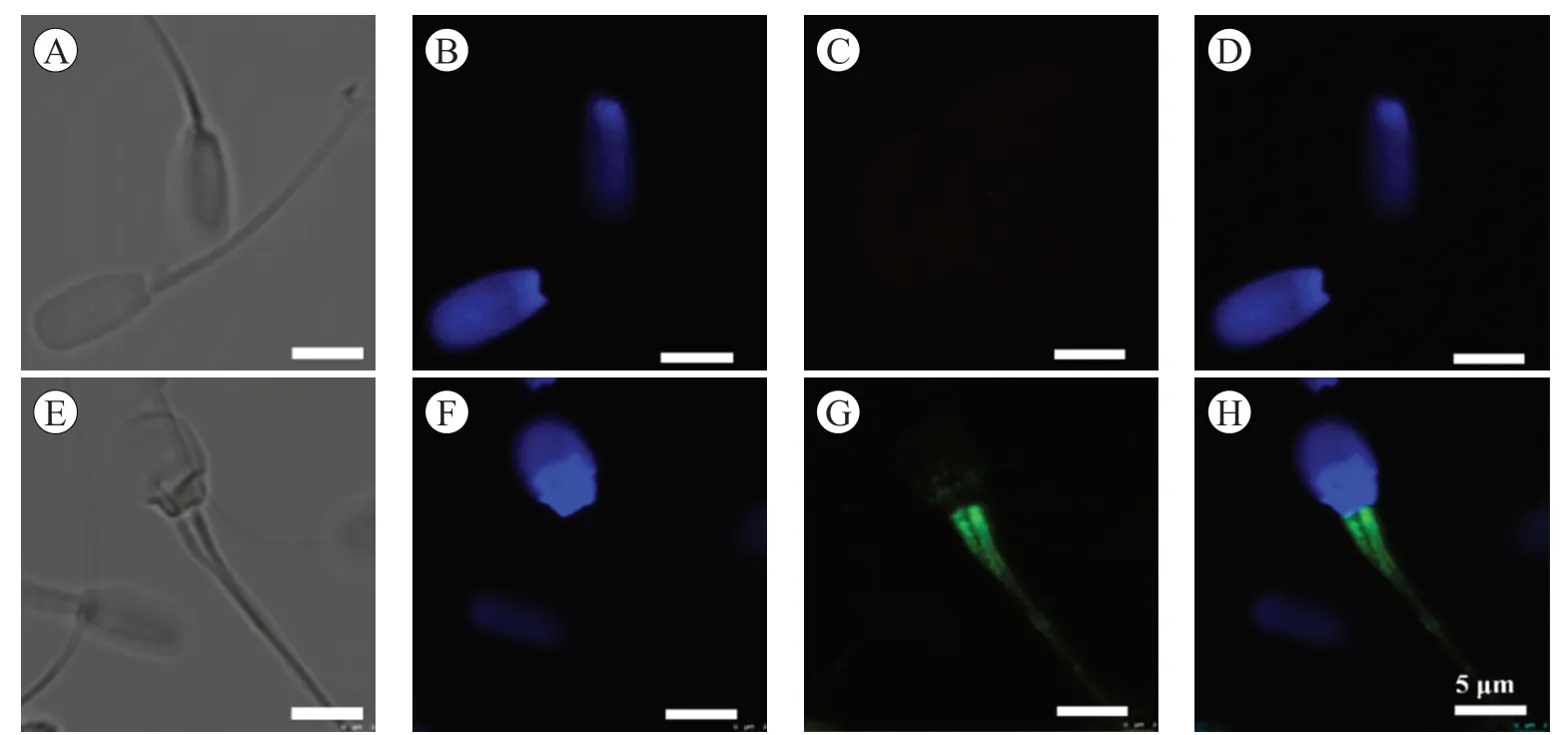
圖4 Hsp90 蛋白在豬精子中的表達
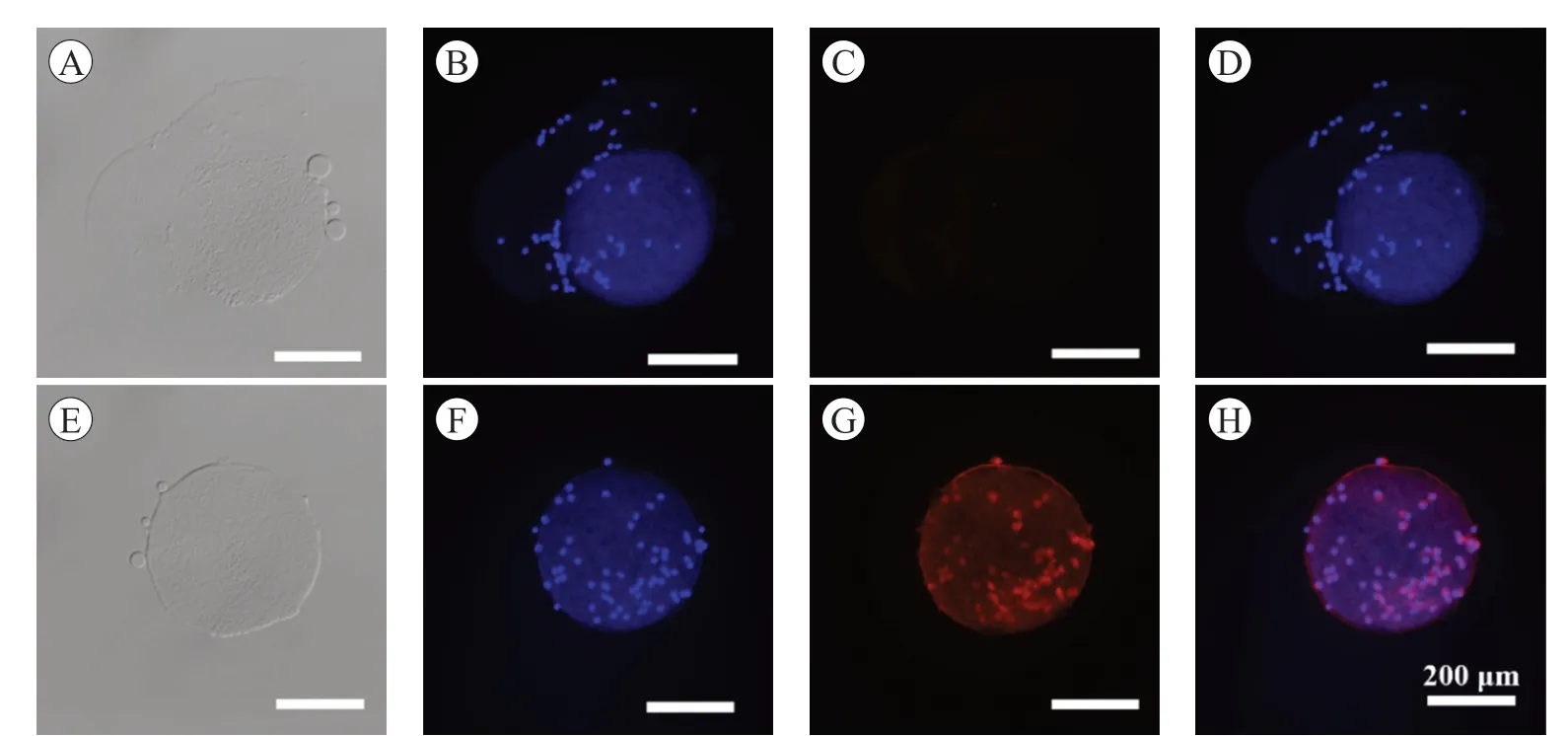
圖5 Hsp90 蛋白在豬卵丘-卵母細胞復合體中的表達
3 討論
Hsp90 蛋白濃度在豬精液冷凍過程中發生了顯著的變化,冷凍使豬精子的化學組成和結構都發生了顯著的改變,而豬精子中蛋白質的變化對豬精子冷凍效率具有極其重要的影響[13]. 冷凍保存可以有效提高種精液的利用率和延長精液的使用時間[14]. 但在冷凍解凍后,大部分精子的活力喪失,導致精子品質下降,繁殖力下降,從而影響優良種公豬的種用價值. 伴隨著豬精子冷凍程序的進行,Hsp90 的表達量逐漸降低,且其表達量下降開始于精子品質下降之前,Hsp90 表達量的大幅降低可能與冷凍過程中豬精子的活率下降相關[15]. 隨后,Huang 等利用Hsp90 的抑制劑格爾德霉素進一步驗證了Hsp90 與精子活力的關系,結果證明Hsp90 在調控豬精子活力方面發揮了重要作用[16],格爾德霉素增加一氧化氮的產生并促進公豬精子獲能[17]. 另外,Hsp90 在非獲能條件下長時間熱應激期間維持公豬精子活力和線粒體膜電位的作用,在獲能條件下,所研究的精子參數在孵育4 h 后均不受Hsp90 抑制劑的影響[18].
Hsp90 調控生精細胞的分裂,參與精子發生過程,在生殖細胞的快速定位、精子微管的形成、保護精子免受氧化應激、調節獲能等過程中發揮不可缺少的作用[1,19-21]. Volpe 等研究發現,Hsp90 定位于貓精子尾部的中段,馬的精子則是在頸部和尾部的全段,但在豬精子上的定位是在精子中段的原生質粒上,在豬精子的免疫熒光圖片上,熒光信號在中段的近端和遠端末端以小單點的形式持續存在[22]. 在本試驗中,可以清楚地看到Hsp90 蛋白在精子的頸部和尾部中段表達,本研究在前人的基礎上,更加確定了Hsp90 蛋白在精子中的位置,結果的差異可能與精子的成熟、品種和抗體種類有關. Hsp90 在頸部和尾部的定位與其在精子運動中的作用有關,但需要進一步的研究來證明這些分子與精子的運動和受精能力的功能相關性[23].研究發現,卵母細胞質量的優劣取決于細胞核和細胞質是否發育成熟,當兩者都發育成熟的時候,才能進行受精;卵母細胞的成熟依賴于卵丘細胞,它們之間進行營養物質和信號分子轉運[24]. 在熱中性或熱休克條件下體外成熟期間抑制Hsp90 會損害牛卵母細胞的發育能力[25]. 本研究發現Hsp90 在豬卵母細胞和卵丘細胞中表達,該蛋白可能對卵母細胞與卵丘細胞之間交流互作產生影響,可為相關生理機制的研究提供基礎數據.
本研究結果顯示Hsp90 在豬主要的繁殖組織和生殖細胞中高表達,可為我國豬精液冷凍技術和胚胎發育的研究提供理論基礎. Hsp90 蛋白的表達與豬精子品質的提高息息相關[26-27],該精液蛋白的直接影響精子的活力和獲能,對豬精液冷凍添加劑的研究具有指導價值. 另外,研究發現,將重組Hsp90 蛋白加入雞精液稀釋劑中,解凍后的精子活力顯著提高[28]. 后續研究的設計是建立表達豬源Hsp90 蛋白的細胞系,獲得重組蛋白Hsp90,將其加入豬精液稀釋劑中,為解凍融精子冷凍損傷的潛在分子機制和提高凍融豬精子生育能力提供新思路和實驗數據.
4 結論
綜上所述,在組織層面發現Hsp90 主要在豬的睪丸和卵巢中高表達;在細胞層面發現Hsp90 在睪丸的間質細胞、次級精母細胞和精子細胞中表達,在卵巢的卵母細胞、卵丘細胞和顆粒細胞中表達,Hsp90 蛋白定位在精子的頸部和尾巴中部,以及卵丘細胞的細胞質和卵母細胞中.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
