餌料對鹵蟲幼體生長、存活和營養(yǎng)組成的影響
2024-03-05 07:00:04吳水清吳建紹郭少鵬仇登高李雷斌羅輝玉
漁業(yè)研究 2024年1期
吳水清,吳建紹,郭少鵬,溫 憑,仇登高,李雷斌,羅輝玉
(福建省水產(chǎn)研究所,福建省海洋生物增養(yǎng)殖與高值化利用重點實驗室,福建 廈門 361013)
鹵蟲(Artemia)俗稱豐年蟲,又稱鹽水豐年蟲、豐年蝦等,是廣泛分布于自然鹽湖和人工鹽池等高鹽度水域中的小型甲殼類動物[1]。鹵蟲含有豐富的氨基酸、脂肪酸等營養(yǎng)物質(zhì)[2],同時因價格合適、容易獲取并能夠穩(wěn)定供應(yīng),而被廣泛作為海水魚類和蝦蟹類苗種培育的生物餌料[3-4]。但是因生活環(huán)境條件的差異,不同的品系、產(chǎn)地來源和采收時間的鹵蟲卵和鹵蟲幼體在大小、營養(yǎng)組成等方面存在較大差異[5-6],特別是高度不飽和脂肪酸的含量已成為水產(chǎn)苗種培育中選擇鹵蟲種類重要的影響因素[7-8]。在水產(chǎn)動物苗種培育中,通常是投喂鹵蟲無節(jié)幼體,或者是將鹵蟲無節(jié)幼體強化之后再使用,如作為日本川鰈(Paralichthysolivaceus)幼苗[9]、云龍石斑魚(Epinephelusmoara♀×E.lanceolatus♂)[10]和赤點石斑魚(E.akaara)魚苗[11]、凡納濱對蝦(Litopenaeusvannamei)[12]、匙吻鱘(Polyodonspathala)仔魚[13]和羅氏沼蝦(Macrobrachiumrosenbergii)幼體[14]等的餌料,然而在水產(chǎn)動物苗種的生長中,使用鹵蟲后期幼體進行投喂也具有較高的現(xiàn)實意義。
鹵蟲是典型的非選擇性濾食、雜食動物,如單細胞微藻、酵母和有機質(zhì)碎屑等都可以用于鹵蟲的培養(yǎng)[15]。利用湛江叉鞭金藻(Dicrateriazhanjiangensis)、亞心形扁藻(Platymonassubcordiformis)和小球藻(Chlorellapyenoidosa)分別投喂鹵蟲幼體,5 d的培養(yǎng)結(jié)果顯示,3種微藻均能促進鹵蟲幼體的生長[16];采用酵母粉為主要餌料開展的鹵蟲高密度養(yǎng)殖,經(jīng)過17 d的養(yǎng)殖,獲得了7.2 kg/m2鹵蟲產(chǎn)量的良好效果[17];利用麥糠微粒投喂鹵蟲 10 d,培養(yǎng)的鹵蟲幼體平均體長達到0.3 cm以上[18]。目前,有關(guān)飼養(yǎng)鹵蟲的研究大多集中在鹵蟲幼體營養(yǎng)強化、不飽和脂肪酸含量富集等方面,對多日齡鹵蟲養(yǎng)殖過程中的生長及其營養(yǎng)成分的綜合評價較少。為進一步了解餌料對鹵蟲生長、存活和營養(yǎng)組成的影響,本研究采用3種餌料[酵母(Saccharomycescerevisiae)、小球藻藻粉和發(fā)酵蝦片]分別投喂孵化的鹵蟲無節(jié)幼體,統(tǒng)計分析其生長和存活情況,比較分析3種餌料飼養(yǎng)的鹵蟲幼體的營養(yǎng)成分,以期為鹵蟲的養(yǎng)殖、營養(yǎng)強化方式提供參考,也為鹵蟲在海水經(jīng)濟動物苗種培育中的應(yīng)用提供科學(xué)依據(jù)。
1 材料與方法
1.1 試驗材料
試驗于2022年3—6月在福建漳州東山縣逸昌水產(chǎn)養(yǎng)殖場苗種培育車間進行。鹵蟲卵和小球藻藻粉(粗蛋白40.6%、粗脂肪5.6%)由山東宏福水產(chǎn)養(yǎng)殖有限公司提供;酵母為湖南仁恒生物科技有限公司生產(chǎn)(蛋白質(zhì)40.0%、粗脂肪4.3%);發(fā)酵蝦片購于廈門市海林生物科技有限公司(粗蛋白45.2%、粗脂肪9.6%)。
1.2 鹵蟲孵化與養(yǎng)殖
1.2.1 鹵蟲孵化
鹵蟲卵放置于底部透光的500 L錐形桶中,放卵密度為1 g/L,孵化溫度為28 ℃,24 h充氣,溶解氧(DO)維持在4~5 mg/L,光照強度5 000~6 000 lx。經(jīng)過24 h的孵化,檢查發(fā)現(xiàn)大部分鹵蟲卵已經(jīng)孵化之后,停止充氣。除底部外,錐形孵化桶上部均遮光,靜置10 min,根據(jù)鹵蟲的趨光性,利用150目的篩絹網(wǎng)在錐形孵化桶底部排水口處收集剛孵化的鹵蟲無節(jié)幼體(Ⅰ期鹵蟲無節(jié)幼體)。
1.2.2 鹵蟲養(yǎng)殖
鹵蟲養(yǎng)殖設(shè)計3個餌料組:酵母、小球藻藻粉、發(fā)酵蝦片(蝦片∶酵母∶水按照10∶1∶5密封發(fā)酵48 h),每種餌料均設(shè)置3個平行組,鹵蟲無節(jié)幼體按照1個/mL的密度接種于1 000 L養(yǎng)殖桶中。每天投喂3次(6:00、14:00、22:00),日換水量為20%,3種餌料初始每次每日投喂量均為10 g,之后每天投喂量增加20%。酵母經(jīng)300目的篩絹網(wǎng)揉搓后再進行投喂;小球藻藻粉直接投喂;發(fā)酵蝦片混勻后投喂,養(yǎng)殖周期均為6 d。試驗用海水均為砂濾自然海水,試驗期間pH為7.5~8.5,溫度為(25±0.5)℃,鹽度為25,24 h充氣,光照強度3 000~5 000 lx。
1.3 測定指標與方法
1.3.1 生長性狀測量和存活率分析
隨機取200只鹵蟲無節(jié)幼體,解剖鏡下測量其體長,作為試驗初始體長。試驗組每隔2天隨機取200只鹵蟲幼體在解剖鏡下測量體長。試驗結(jié)束時,將養(yǎng)殖桶鹵蟲樣品混勻后,隨機取樣1 000 mL進行全部計數(shù),計算存活率。
相關(guān)指標的計算方法如下:
增長率(%)=(L2-L1)/L1×100
(1)
特定生長率(%/d)=(lnL2-lnL1)/t×100
(2)
存活率(%)=N2/N1×100
(3)
式(1)~(3)中:L2為終末體長,μm;L1為初始體長,μm;t為投喂天數(shù),d;N2為終末數(shù)量;N1為初始數(shù)量。
1.3.2 營養(yǎng)組成分析
本試驗先取部分剛孵化的鹵蟲無節(jié)幼體進行清洗、過濾之后,將其冰凍保存、待測,其營養(yǎng)成分分析結(jié)果作為本底對照及與鹵蟲養(yǎng)殖試驗結(jié)果的比較。至試驗結(jié)束時,利用80目篩絹網(wǎng)收集所有養(yǎng)殖后的鹵蟲幼體,清洗干凈、過濾之后,用于營養(yǎng)成分分析。水分測定參照GB 5009.3—2010直接干燥法;灰分測定參照GB 5009.4—2010馬弗爐高溫灼燒法;粗蛋白測定參照GB 5009.5—2010凱氏定氮法測定;粗脂肪參照GB/T 5009.6—2003索氏抽提法;氨基酸測定參照GB/T 5009.124—2003酸水解法;脂肪酸測定參照GB 5009.168—2016氣相色譜法。
1.4 數(shù)據(jù)分析
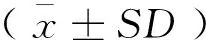
2 結(jié)果與分析
2.1 不同餌料組的鹵蟲幼體生長比較
采用酵母、小球藻藻粉和發(fā)酵蝦片3種餌料對鹵蟲無節(jié)幼體進行6 d的養(yǎng)殖,各組鹵蟲幼體生長情況如表1所示。酵母組鹵蟲幼體生長最慢,體長為(1 070.75±84.32)μm;發(fā)酵蝦片組鹵蟲幼體生長最快,體長為(1 234.54±102.32)μm;3種餌料對鹵蟲幼體生長速率的影響依次為酵母組<小球藻藻粉組<發(fā)酵蝦片組,且不同餌料對鹵蟲幼體生長影響顯著(P<0.05),發(fā)酵蝦片組顯著大于小球藻藻粉組和酵母組(P<0.05),小球藻藻粉組和酵母組差異不顯著(P>0.05)。3個餌料組鹵蟲幼體體長增長量、增長率和特定生長率的變化均與體長變化一致。

表1 3種不同餌料對鹵蟲幼體生長的影響Tab.1 The effects of three different diets on growth of Artemia nauplii
2.2 不同餌料組對鹵蟲幼體存活率的影響
鹵蟲幼體存活率情況如圖 1 所示。小球藻藻粉組的鹵蟲幼體存活率最高,為(86.87±3.01)%;其次為發(fā)酵蝦片組,為(85.73±0.50)%;酵母組最低,為(79.17±3.82)%。酵母組的鹵蟲幼體存活率與小球藻藻粉組、發(fā)酵蝦片組均差異顯著(P<0.05),小球藻藻粉組和發(fā)酵蝦片組存活率差異不顯著(P>0.05)。
2.3 不同餌料組鹵蟲幼體常規(guī)營養(yǎng)成分
鹵蟲無節(jié)幼體和經(jīng)3種餌料養(yǎng)殖的鹵蟲幼體常規(guī)營養(yǎng)成分見表2。鹵蟲無節(jié)幼體的粗蛋白質(zhì)含量最低,為(52.68±0.303)%,顯著低于酵母組、小球藻藻粉組和發(fā)酵蝦片組(P<0.05);小球藻藻粉組和發(fā)酵蝦片組鹵蟲的蛋白質(zhì)含量顯著高于酵母組(P<0.05)。鹵蟲無節(jié)幼體的粗脂肪含量最高,為(11.93±0.033)%,顯著高于酵母組、小球藻藻粉組和發(fā)酵蝦片組(P<0.05);3種餌料投喂的鹵蟲幼體的粗脂肪含量大小依次為發(fā)酵蝦片組>小球藻藻粉組>酵母組。水分含量在各組中的大小依次為酵母組>發(fā)酵蝦片組>小球藻藻粉組>鹵蟲無節(jié)幼體。鹵蟲無節(jié)幼體灰分含量最高,為(21.56±0.009)%,顯著高于酵母組、小球藻藻粉和發(fā)酵蝦片組(P<0.05),3種餌料投喂的鹵蟲灰分含量差異不顯著(P>0.05)。
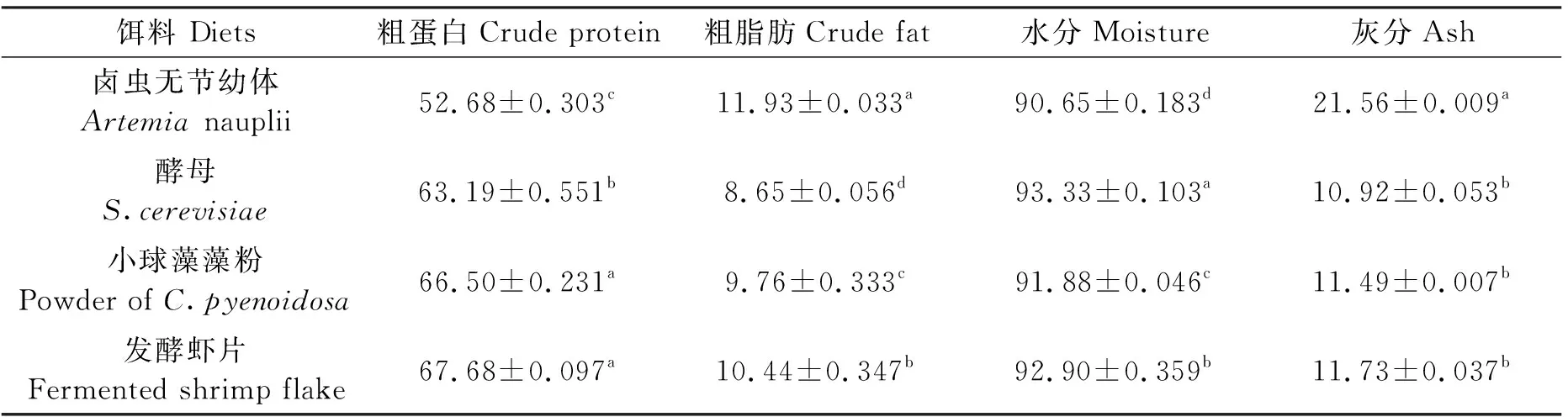
表2 鹵蟲無節(jié)幼體和經(jīng)3種餌料養(yǎng)殖的鹵蟲幼體常規(guī)營養(yǎng)成分(干物質(zhì))Tab.2 General nutrition composition of new hatched Artemia nauplii and Artemia nauplii fed with three different diets (dry weight) %
2.4 不同餌料組鹵蟲幼體氨基酸組成及含量
鹵蟲無節(jié)幼體和經(jīng)3種餌料養(yǎng)殖的鹵蟲氨基酸組成及含量見表3。在鹵蟲無節(jié)幼體和3種餌料組鹵蟲中均檢測到18種氨基酸。鹵蟲無節(jié)幼體的各氨基酸含量均顯著低于3個餌料組(P<0.05)。酵母組鹵蟲幼體的絲氨酸含量顯著低于小球藻藻粉組和發(fā)酵蝦片組(P<0.05)。小球藻藻粉組鹵蟲幼體的色氨酸含量顯著高于酵母組和發(fā)酵蝦片組(P<0.05);酪氨酸含量顯著高于發(fā)酵蝦片組(P<0.05),但與酵母組差異不顯著(P>0.05)。不同餌料組鹵蟲幼體的丙氨酸、精氨酸、天門冬氨酸、谷氨酸、甘氨酸、組氨酸、異亮氨酸、亮氨酸、賴氨酸、苯丙氨酸、脯氨酸、蘇氨酸、纈氨酸含量大小依次為發(fā)酵蝦片組>小球藻藻粉組>酵母組(P<0.05)。發(fā)酵蝦片組的鹵蟲幼體的氨基酸總量和必需氨基酸總量均最高,大小均依次為發(fā)酵蝦片組>小球藻藻粉組>酵母組(P<0.05)。
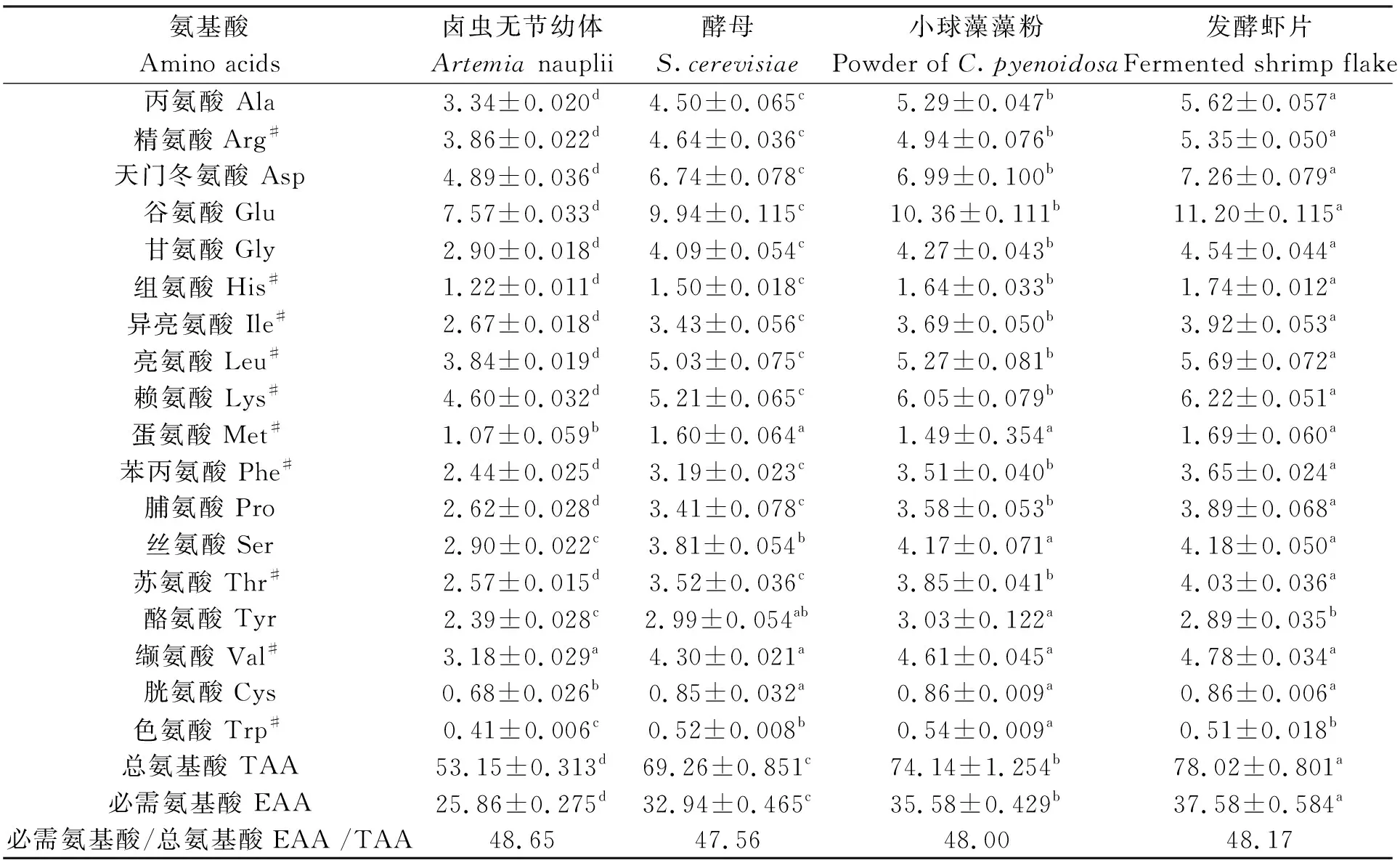
表3 鹵蟲無節(jié)幼體和經(jīng)3種餌料養(yǎng)殖的鹵蟲幼體氨基酸組成及含量(干物質(zhì))Tab.3 Amino acids composition and their contents of new hatched Artemia nauplii and Artemia nauplii fed with three different diets (dry weight) %
2.5 不同餌料組鹵蟲幼體脂肪酸組成及含量
鹵蟲無節(jié)幼體和經(jīng)3種餌料養(yǎng)殖的鹵蟲幼體的脂肪酸組成及含量見表4。在鹵蟲無節(jié)幼體中檢測到9種脂肪酸,其他組均缺少豆蔻酸。總脂肪酸、不飽和脂肪酸和單不飽和脂肪酸含量大小依次為鹵蟲無節(jié)幼體>發(fā)酵蝦片組>酵母組>小球藻藻粉組(P<0.05)。
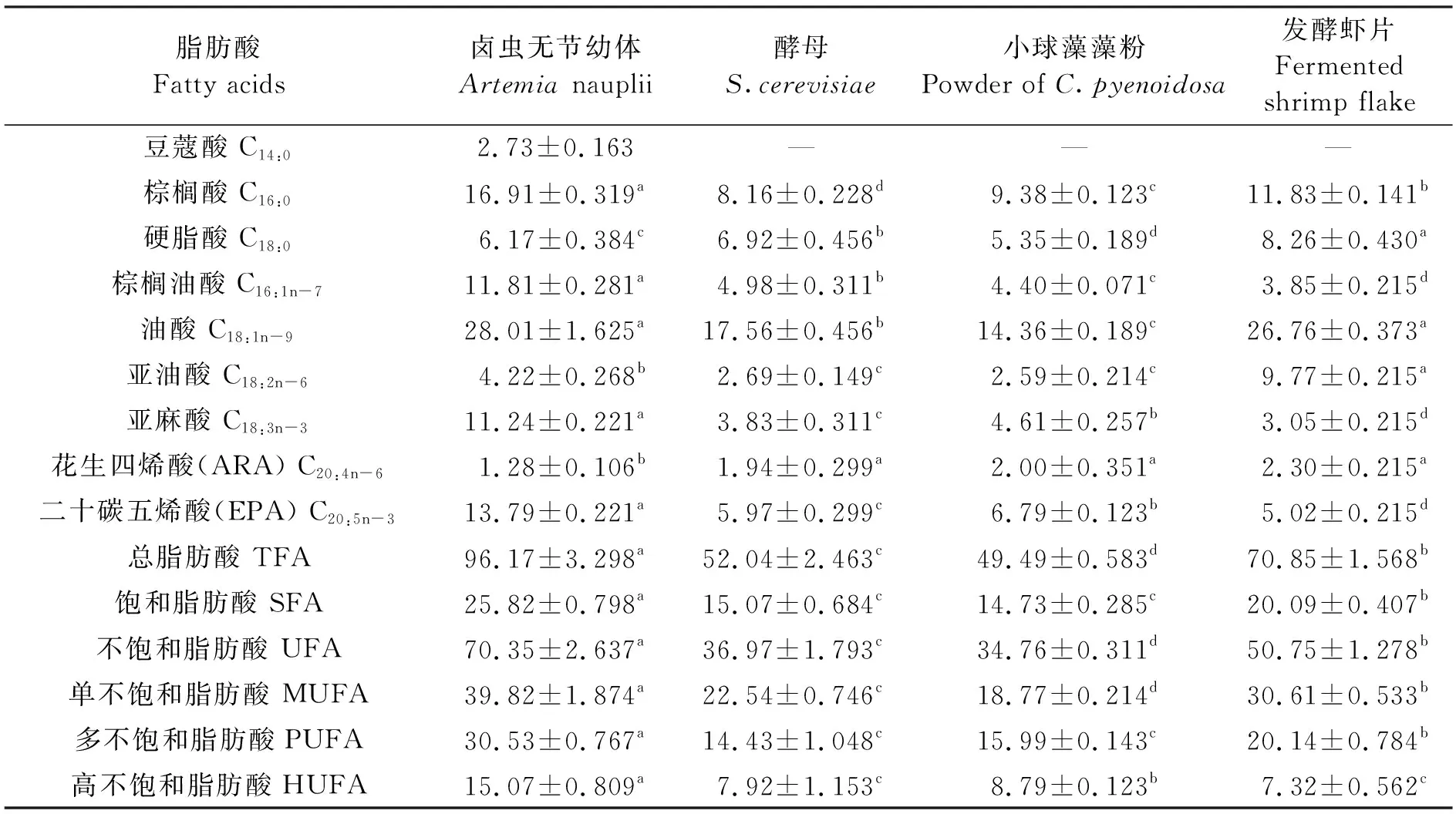
表4 鹵蟲無節(jié)幼體和經(jīng)3種餌料養(yǎng)殖的鹵蟲幼體脂肪酸組成及含量(干物質(zhì))Tab.4 Fatty acid profile of new hatched Artemia nauplii and Artemia nauplii fed with three different diets (dry weight) mg/g
小球藻藻粉組鹵蟲幼體的亞麻酸和EPA含量分別為(4.61±0.257)、(6.79±0.123)mg/g,均顯著高于酵母組和發(fā)酵蝦片組(P<0.05);酵母組鹵蟲幼體的棕櫚油酸含量為(4.98±0.311)mg/g,顯著高于小球藻藻粉組和發(fā)酵蝦片組;發(fā)酵蝦片組鹵蟲幼體的油酸和亞油酸含量分別為(26.76±0.373)、(9.77±0.215)mg/g,均顯著高于酵母組和小球藻藻粉組(P<0.05);高不飽和脂肪酸含量大小依次為鹵蟲無節(jié)幼體>小球藻藻粉組>酵母組>發(fā)酵蝦片組。
3 討論
3.1 餌料對鹵蟲幼體生長和存活的影響
鹵蟲因生活環(huán)境的不同而具有獨特的特性,不同產(chǎn)地(品系)的鹵蟲卵徑、無節(jié)幼體體長和營養(yǎng)組成存在明顯差異[19],一般高海拔地區(qū)鹵蟲卵徑大、顏色較深,孵化的無節(jié)幼體個體也較大[20-21];較高的溫度環(huán)境的鹵蟲卵徑較小、無節(jié)幼體也較小[22]。本研究所用的鹵蟲卵的產(chǎn)地為山東,孵化的鹵蟲無節(jié)幼體平均體長為(404.15±18.81)μm,明顯小于西藏4個鹽湖產(chǎn)的鹵蟲無節(jié)幼體(519.6~601.1 μm)[8],但與山東濰坊產(chǎn)地的鹵蟲無節(jié)幼體(415.8±23.6 μm)[23]和渤海灣鹽田[(350.10±21.44)μm][12]相當(dāng),而同一產(chǎn)地的鹵蟲卵和無節(jié)幼體大小的差異可能與孤雌和兩性品系混雜鹵蟲卵有關(guān)[23]。
酵母、小球藻藻粉和發(fā)酵蝦片是鹵蟲養(yǎng)殖中常用的餌料,這3種餌料均能滿足鹵蟲無節(jié)幼體生長的需求。在用鹽藻、酵母、麩皮3種餌料培養(yǎng)鹵蟲的試驗中發(fā)現(xiàn),酵母組鹵蟲的生長和存活率均最低[24];利用不同餌料養(yǎng)殖南京豐年蟲(ChirocephalusnankinensisShen)的結(jié)果也表明,酵母養(yǎng)殖的南京豐年蟲存活率較低[25]。本研究中鹵蟲無節(jié)幼體平均成活率為79.17%~86.87%,其中發(fā)酵蝦片組和小球藻藻粉組的鹵蟲幼體存活率顯著高于酵母組(P<0.05),原因可能是酵母菌群促進了培養(yǎng)水體中有機物質(zhì)含量的增加,導(dǎo)致水質(zhì)敗壞,從而降低了鹵蟲的存活率[26],這在輪蟲等濾食性生物培養(yǎng)中也有相似的結(jié)果[27-28]。鹵蟲生長還與其獲取的餌料大小、營養(yǎng)等有關(guān),有研究認為鹵蟲生長與餌料中蛋白質(zhì)的含量呈正相關(guān)[29]。本養(yǎng)殖試驗中各組鹵蟲幼體平均體長為1 070.75~1 234.54 μm,而發(fā)酵蝦片組鹵蟲幼體的生長顯著優(yōu)于酵母組和小球藻藻粉組(P<0.05),小球藻藻粉組鹵蟲幼體的生長與酵母組差異不顯著(P>0.05),這與餌料中蛋白質(zhì)含量差異基本一致(發(fā)酵蝦片粗蛋白45.2%、酵母粗蛋白40.0%、小球藻藻粉粗蛋白40.6%)。
3.2 鹵蟲基本營養(yǎng)成分比較分析
蛋白質(zhì)和脂肪含量是評價水產(chǎn)動物餌料營養(yǎng)價值的重要指標[30]。本研究中3種餌料投喂的鹵蟲幼體的粗蛋白含量為63.19%~67.68%、粗脂肪含量為8.65%~10.44%,均有較高的營養(yǎng)價值,可以滿足水產(chǎn)動物的生長需求。通過跟蹤不同餌料下鹵蟲總脂肪含量變化的結(jié)果,發(fā)現(xiàn)隨著鹵蟲幼體體長的增長,總脂肪含量呈現(xiàn)降低趨勢[31]。本研究中粗脂肪含量的變化情況也類似,初孵鹵蟲無節(jié)幼體的粗脂肪含量為(11.93±0.033)%,顯著高于3種餌料投喂的6日齡鹵蟲幼體(P<0.05)。以小球藻藻粉和發(fā)酵蝦片投喂的鹵蟲幼體粗蛋白和粗脂肪含量要明顯高于以酵母喂養(yǎng)的(P<0.05),這與張登瀝等[24]的研究結(jié)果相似,相對單獨的酵母,單細胞藻類和發(fā)酵產(chǎn)品更適合作為鹵蟲高密度培養(yǎng)的餌料。
3.3 不同餌料對鹵蟲幼體氨基酸含量的影響
水產(chǎn)動物對蛋白質(zhì)的需要量實際是對必需氨基酸和非必需氨基酸比例的需要,動物餌料中必需氨基酸總量應(yīng)不低于粗蛋白的1/3[32],魚蝦類限制性氨基酸通常是精氨酸、蛋氨酸、賴氨酸和苯丙氨酸[33],特別是對精氨酸的需求達到3.3%~6.8%[34-35]。本研究中鹵蟲無節(jié)幼體和3個餌料組鹵蟲幼體的氨基酸含量占干物質(zhì)的53.15%~78.02%,必需氨基酸與氨基酸總量比值為47.56%~48.65%,均超過40%,精氨酸占比為3.34%~5.62%,能夠滿足水產(chǎn)動物對蛋白質(zhì)的需求。通過對比鹵蟲無節(jié)幼體和3個餌料組鹵蟲幼體的氨基酸含量,發(fā)現(xiàn)6日齡鹵蟲幼體在氨基酸和必需氨基酸總量上均有顯著提高,特別是發(fā)酵蝦片組和小球藻藻粉組顯著高于酵母組(P<0.05)。但用小球藻和三角褐指藻(Phaeodactylumtricornutum)強化培養(yǎng)5~7 d的鹵蟲無節(jié)幼體的氨基酸總量和必需氨基酸總量卻都小于初孵的鹵蟲無節(jié)幼體[16],這可能是新鮮的小球藻因有完整的細胞壁而不易被消化,而本研究采用小球藻藻粉,其大部分細胞壁已被破碎,有利于鹵蟲的營養(yǎng)吸收[36]。
3.4 不同餌料對鹵蟲幼體脂肪酸含量的影響
鹵蟲卵和鹵蟲無節(jié)幼體中脂肪酸含量受產(chǎn)地影響很大,通常與所處的水環(huán)境因子和餌料的脂肪酸組成密切相關(guān)[37]。西藏4個鹽湖鹵蟲無節(jié)幼體含有較高的 二十二碳六烯酸(DHA)[8];在對不同鹵蟲幼體脂肪酸的測定中,結(jié)果也發(fā)現(xiàn)西藏雙湖鹵蟲卵和鹵蟲幼體中花生四烯酸(ARA)、二十碳五烯酸(EPA)和DHA顯著高于其他產(chǎn)地鹵蟲幼體[12]。對6個產(chǎn)地(3個為沿海鹽場、3個為內(nèi)陸鹽湖)鹵蟲無節(jié)幼體脂肪酸的組成分析發(fā)現(xiàn),其含有豐富的EPA,但均未檢測到DHA[7]。本研究也有類似的結(jié)果,試驗中鹵蟲卵來源于山東沿海鹽場,鹵蟲無節(jié)幼體也未檢測到DHA。分析西藏地區(qū)鹵蟲含有較高的DHA的原因,可能與強烈的紫外線照射、極低的水溫及水體中浮游植物和微生物含有較高的EPA和DHA含量有關(guān)[8,12]。
脂肪酸對魚類和蝦蟹類具有重要的作用,但是它們往往不能夠自行合成多不飽和脂肪酸,特別是高不飽和脂肪酸(HUFA)[38],餌料中高HUFA 含量和高 DHA/EPA比值有利于魚類和蝦蟹類的存活和生長發(fā)育[39],因此在鹵蟲生產(chǎn)中,通常添加不飽和脂肪酸的餌料或添加劑來營養(yǎng)強化鹵蟲[40],例如通過魚油和裂殖壺藻(Schizochytrium)進行的短期營養(yǎng)強化,可顯著提高鹵蟲HUFA含量。采用不同微藻強化鹵蟲,發(fā)現(xiàn)小球藻基本不能提高擬成蟲期鹵蟲的DHA含量,但可以促進鹵蟲幼體生長和EPA的積累[41];此外,小球藻等餌料不能提高鹵蟲幼體的蛋白質(zhì)含量[16]。小球藻作為鹵蟲的生物餌料,含有豐富的蛋白質(zhì)和不飽和脂肪酸含量,特別是含有較高的EPA、不含或者少量含有DHA[42-44]。蝦片是對蝦苗種微粒飼料,具有豐富的氨基酸、脂肪酸組成[45-46],其發(fā)酵產(chǎn)物在生物餌料培養(yǎng)中起到重要的肥水作用。本試驗中,發(fā)酵蝦片組總脂肪酸含量和多不飽和脂肪酸含量顯著高于小球藻藻粉組和酵母組,但是小球藻藻粉投喂的鹵蟲幼體的HUFA、EPA含量明顯高于酵母組和發(fā)酵蝦片組,表明發(fā)酵蝦片和小球藻藻粉相對于酵母具有鹵蟲營養(yǎng)強化的優(yōu)勢,可用于鹵蟲的養(yǎng)殖生產(chǎn)。
