魚類水產品中耐冷性摩根菌Morganella psychrotolerans污染情況與分離株組胺生成能力
2024-04-08 02:28:22陳勝軍吳燕燕李春生王悅齊
食品科學 2024年5期
李 錦,王 迪,陳勝軍,吳燕燕,李春生,王悅齊
(1.上海海洋大學食品學院,上海 201306;2.中國水產科學研究院南海水產研究所,農業農村部水產品加工重點實驗室,國家水產品加工技術研發中心,廣東 廣州 510300;3.三亞熱帶水產研究院,海南省深遠海漁業資源高效利用與加工重點實驗室,海南 三亞 572018)
組胺中毒是全球范圍內因水產品消費而引發的食品安全問題[1-2]。通常情況下,食用含有高于1000 mg/kg組胺水平的魚類后,中毒癥狀可能在數分鐘至3 h內出現[3],組胺引起的中毒反應,包括潮紅、頭痛、水腫、蕁麻疹、腹瀉、抽筋和嘔吐等典型的過敏樣食物中毒反應[4-6]。近年來,我國因攝入含有組胺的水產品而引發的食物中毒屢見不鮮[7],也曾報道過引發死亡的組胺中毒事件[8]。水產品中組胺的生成與積累主要是由于微生物的作用,伴隨著捕獲后魚體的死亡,魚體內微生物開始大量繁殖,其中產組胺微生物產生的組氨酸脫羧酶可將魚體中的組氨酸轉化為組胺。組胺一旦在食品中生成,普通的冷凍、加熱、微波、煙熏等加工方式無法將其破壞[9]。各國家對水產品中組胺含量都有明確的限量規定,其中歐盟允許鮮魚中存在的組胺含量最高不得高于200 mg/kg,其他水產品中最高不得超過400 mg/kg[10];美國食品藥品監督管理局(Food and Drug Administration,FDA)要求水產品中組胺濃度的安全標準為不高于50 mg/kg;我國GB 2733—2015《鮮、凍動物性水產品》中也對水產品中組胺含量做了明確的限量規定,其中青皮紅肉魚類不得高于400 mg/kg,其余水產品中組胺含量應小于200 mg/kg[11]。組胺生成菌廣泛存在于海水、魚的腸道、鰓與體表中[12]。在高于等于15 ℃的條件下培養48 h,組胺生成菌在金槍魚魚汁或含1%~2%L-組氨酸的胰蛋白胨大豆肉湯(L-histidine tryptone soy broth,TSBH)培養基中生成組胺量超過1000 mg/L,則可以將其定義為具有高組胺生成能力的組胺生成菌,低于500 mg/L定義為具有低組胺能力的組胺生成菌[13-14]。常見的組胺生成菌有Morganella morganii[15]、Photobacteriumdamselae[16]、P.phosphoreum[17]、Klebsiella oxytoca[18]、M.psychrtolerans[19]等,其中P.phosphoreum與M.psychrotolerans可在低溫下生長并生成組胺[20]。
M.psychrotolerans作為在2006年首次被分離并命名的菌株[20-23],其與M.morganii的16S rRNA基因序列分析相似性高達98.6%,常被誤判為中溫性組胺生成菌M.morganii。但M.psychrotolerans菌株7 個蛋白編碼管家基因(atpD、dnaN、gyrB、hdc、infB、rpoB和tuf)在兩株菌之間的序列相似性均小于90.9%[20]。M.psychrotolerans可在0 ℃生長,而M.morganii在低于7 ℃條件下則無法生長[20]。研究表明,M.psychrotolerans廣泛存在于日本、丹麥和美國等海水與水產品中,且其分離株均顯示出較高的組胺生成能力[22-24],且M.psychrotolerans在日本與丹麥均引起過食物中毒[22,25]反應的發生。目前M.psychrotolerans在我國水產品中的污染情況尚鮮見文獻報道。劉紅等[26]從秋刀魚中分離的M.morganii在4 ℃培養24 h后檢測到了22 mg/kg組胺。鑒于M.morganii無法在4 ℃條件下生長,有理由懷疑我國市售水產品中也存在M.psychrotolerans。因此,本研究對廣東省市售水產品中M.psychrotolerans情況開展調查研究,并對其分離株組胺生成能力開展研究,為有效控制我國水產品中組胺積累問題提供理論數據支撐。
1 材料與方法
1.1 材料與試劑
本研究中使用的水產品均隨機購自于廣東省內的水產品市場與超市,選購大小均一的同種商品,將其分類置于冰水中,并于4 h內轉運回實驗室,進行實驗。水產品具體來源詳見表1。

表1 水產品來源Table 1 Sampling locations of commercial fish in this study
胰蛋白胨、酵母提取物 英國Oxoid公司;磷酸二氫鉀、磷酸氫二鉀、硫酸黏菌素、盧哥氏碘液、去離子水北京Solarbio科技有限公司;L-組氨酸鹽酸鹽一水合物、溴甲酚紅紫、碳酸鈣 上海麥克林生化科技有限公司;瓊脂、DNA Marker L(50~500 bp)生工生物工程(上海)股份有限公司;氯化鈉、氯化鐵 國藥集團化學試劑有限公司;磷酸鹽緩沖液(phosphate buffered saline,PBS)蘭杰柯科技有限公司;生理鹽水、胰蛋白胨大豆肉湯(tryptone soy broth,TSB)、大豆酪蛋白瓊脂(tryptose soya agar,TSA)培養基、苯丙氨酸 廣東環凱微生物科技有限公司;組胺測定試劑盒 日本Kikkoman公司;Emerald Amp?Max PCR Master Mix日本Takara公司;酒精 美國Supelco公司;結晶紫染色液 北京康佳宏原生物科技有限公司;沙黃復染液上海源葉生物科技有限公司;煙酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NADH)氧化酶試劑盒、過氧化氫酶試劑盒 北京盒子生工科技有限公司;硝酸鹽還原試劑盒、葡萄糖發酵管 青島高科園海博生物技術有限公司。
M.psychrotoleransJCM16473T模式菌株購自廣東省微生物保藏中心。
1.2 儀器與設備
SQ510C立式壓力蒸汽滅菌器、IN612C低溫培養箱日本Yamato公司;SPX智能型生化培養箱 寧波江南儀器廠;Milli-Q純水系統 美國Millipore公司;NanoDrop2000超微量分光光度計 美國NanoDrop公司;電泳儀凝膠成像系統 北京百思佳特科技有限責任公司;Mastercycler nexus型聚合酶鏈式反應(polymerase chain reaction,PCR)擴增儀 德國艾本德股份公司;3K30臺式高速冷凍離心機 美國Sigma公司;拍打式無菌均質機 杰瑞安科技有限責任公司;金屬浴 上海一恒科學儀器有限公司;顯微鏡 德國Leica公司;渦旋混勻儀 國藥集團化學試劑有限公司。
1.3 方法
1.3.1 培養基的制備
1×Morganella enrichment(MOE)液體培養基[22]:胰蛋白胨10 g/L、酵母提取物5 g/L、磷酸二氫鉀7 g/L、磷酸氫二鉀7 g/L、L-組氨酸鹽酸鹽一水合物10 g/L、溴甲酚紅紫0.03 g/L,蒸餾水1 L,用1 mol/L的鹽酸/氫氧化鈉溶液調至pH 5.1,121 ℃滅菌15 min。
2×MOE液體培養基[22]:胰蛋白胨20 g/L、酵母提取物10 g/L、磷酸二氫鉀14 g/L、磷酸氫二鉀 14 g/L、L-組氨酸鹽酸鹽一水合物20 g/L、溴甲酚紅紫0.06 g/L,蒸餾水1 L,用鹽酸/氫氧化鈉溶液調至pH 5.1,121 ℃滅菌15 min。
按照Niven等[27]的方法制備Niven’s agar固體培養基。
TSBH液體培養基:在TSB液體培養基加入0.01 g/mLL-鹽酸組氨酸,用1 mol/L的鹽酸/氫氧化鈉溶液調至pH 6,121 ℃滅菌15 min。
1.3.2 魚類水產品中M.psychrotolerans的分布污染情況調查
本研究采用最大可能數法-聚合酶鏈式反應(most probable number-polymerase chain reaction,MPN-PCR)法對樣品中的M.psychrotolerans污染情況進行調查。
在實驗開始前,先在配制完成的MOE培養基中加入硫酸黏菌素(32 mg/L;1×MOE培養基中加入0.1 mL,2×MOE培養基中加入0.2 mL)。在超凈臺的無菌環境中將切好的25 g魚肉樣品加入225 mL配制好的無菌PBS中,均質拍打3 min后,分別取1 mL和0.1 mL分別加入到體積為9 mL和9.9 mL的1×MOE培養基中,取10 mL樣品液加入到體積為10 mL的2×MOE培養基中,渦旋。制備成不同梯度濃度的MOE培養基,每個梯度設置3 組平行。將MOE篩選培養基置于12 ℃培養7 d后,觀察培養基顏色變化并記錄[28]。
從顏色發生變化的培養基中吸取1 mL培養液,10000 r/min離心2 min,棄去上層液體,保留菌體沉淀,在沉淀中加入400 μL的Triton-100,渦旋后放入金屬浴中,99 ℃處理10 min,將此作為DNA擴增模板。
對M.psychrotolerans的VasD基因進行PCR定向擴增,PCR擴增引物和擴增體系見表2、3,擴增條件為95 ℃預變性4 min,95 ℃變性30 s,62 ℃退火30 s,72 ℃延伸30 s,共30 個循環,接著72 ℃終延伸4 min,4 ℃保存擴增產物。PCR產物經2%瓊脂糖凝膠電泳,實驗后觀察是否存在VasD基因特異性條帶。

表2 PCR實驗引物[29]Table 2 Primer sequences used for PCR[29]

表3 PCR擴增體系Table 3 Composition of PCR amplification system
1.3.3 魚類水產品中M.psychrotolerans菌株的分離及鑒定
從1.3.2節中得到的陽性試管中取培養液,經適當稀釋后,涂布于Niven’s agar固體培養基,25 ℃培養72 h,挑取平板上具有顏色變化的菌落進行劃線分離,并將分離得到的單菌落繼續依照1.3.2節中的PCR方法進行VasD基因擴增驗證實驗。
將分離后的M.psychrotolerans菌株進一步進行生理生化實驗驗證,包括革蘭氏染色實驗(酒精擦拭玻片,在酒精燈上干燥后,用接種環挑取一環生理鹽水到載玻片上,挑取少量純培養物在水滴上涂抺至均勻分散,將涂片在酒精燈火焰上滅活、固定;滴加結晶紫染色液1~2 滴,染1 min,用去離子水水洗;待干,滴加革蘭氏碘液,1 min后用去離子水水洗;滴加體積分數95%乙醇溶液,直至無紫色脫落為止,用去離子水水洗;油鏡鏡檢、氧化酶實驗、過氧化氫酶實驗、苯丙氨酸脫氨酶實驗(從TSA培養基上挑取培養物,接種于苯丙氨酸培養基上,在25 ℃培養18~24 h后,加入氯化鐵溶液4~5 滴,轉動試管,觀察結果)、硝酸鹽還原實驗、葡萄糖發酵實驗、D-半乳糖產酸實驗驗證[19,28-30]。
瓊脂糖凝膠電泳結果存在VasD基因擴增條帶,且菌株生理生化實驗結果顯示為革蘭氏陰性、氧化酶陰性、過氧化氫酶陽性、苯丙氨酸酶陽性、硝酸鹽還原性、葡萄糖發酵性、D-半乳糖的非酸產生性。PCR與生理生化鑒定雙重實驗確保分離單菌落為M.psychrotolerans。
1.3.4M.psychrotolerans分離株組胺生成能力測定
將分離得到的M.psychrotolerans菌株進行組胺生成能力的測定。勾取TSA平板中分離株單菌落接種于TSB培養基,25 ℃培養20 h,吸取菌液1 mL,經PBS洗滌兩次(10000 r/min、2 min、4 ℃)后,取100 μL接入5 mL TSBH液體培養基中(初始接菌落總數約為1×105CFU/mL),20 ℃培養48 h后,測定菌落總數與組胺含量。組胺測定:取1 mL上述TSBH培養基菌液,100 ℃金屬浴15 min后,以10000 r/min離心2 min,取上清液依照試劑盒中的實驗步驟測定組胺。
1.3.5 分離株與模式菌株生長與組胺生成能力比較
將上述分離到的產組胺能力最強的菌株與模式菌株進行生長與組胺生成能力的比較分析。將分離株與模式菌株分別接入TSB,25 ℃培養20 h,對預培養菌液稀釋100 倍,取0.1 mL稀釋培養液加入到9.9 mL TSBH(pH 6.0)中(初始接菌落總數約4×103CFU/mL),分別置于4 ℃和20 ℃培養,并在不同時間點取樣分別對菌落總數和組胺含量進行測定。
1.4 數據處理與分析
2 結果與分析
2.1 魚類水產品中M.psychrotolerans污染調查
表4顯示,在101~102MPN/100 g范圍內的樣本數為16 個(16%),在102~103MPN/100 g范圍內的樣本數為11 個(11%),在103~104MPN/100 g范圍內的樣本數為5 個(5%),超過104MPN/100 g的樣本數為2 個(2%)。研究結果表明在隨機檢測的廣東省在售的100 個水產品樣本中,陽性樣品率為34%,34 個陽性樣本中包括鯖魚13 個(38.24%)、金槍魚10 個(29.41%)、三文魚5 個(14.71%)、沙丁魚3 個(8.82%)、竹莢魚2 個(5.88%)、鰈魚1 個(2.94%);其中78 個青皮紅肉魚類樣品中呈現28 個陽性樣本(35.9%),22 個白肉魚類樣品中呈現6 個陽性樣本(27.3%)(圖1)。綜上,M.psychrotolerans在廣東在售水產品中廣泛存在,尤其是青皮紅肉魚類水產品,需重點關注。
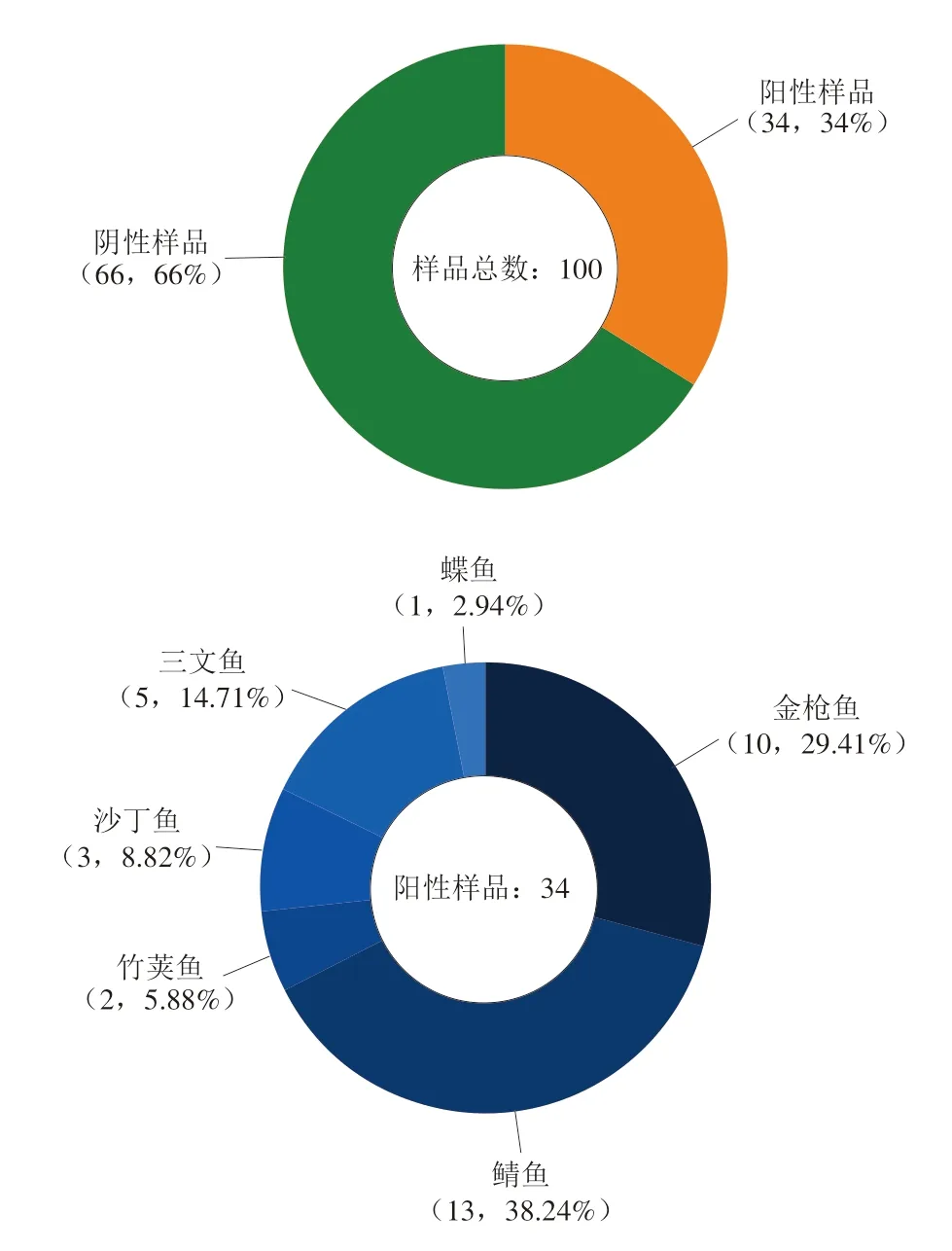
圖1 陽性樣品和陰性樣品占總樣品的比例和不同種類魚在陽性樣品中的占比Fig.1 Proportions of positive and negative samples among total samples and proportions of different kinds of fish in positive samples
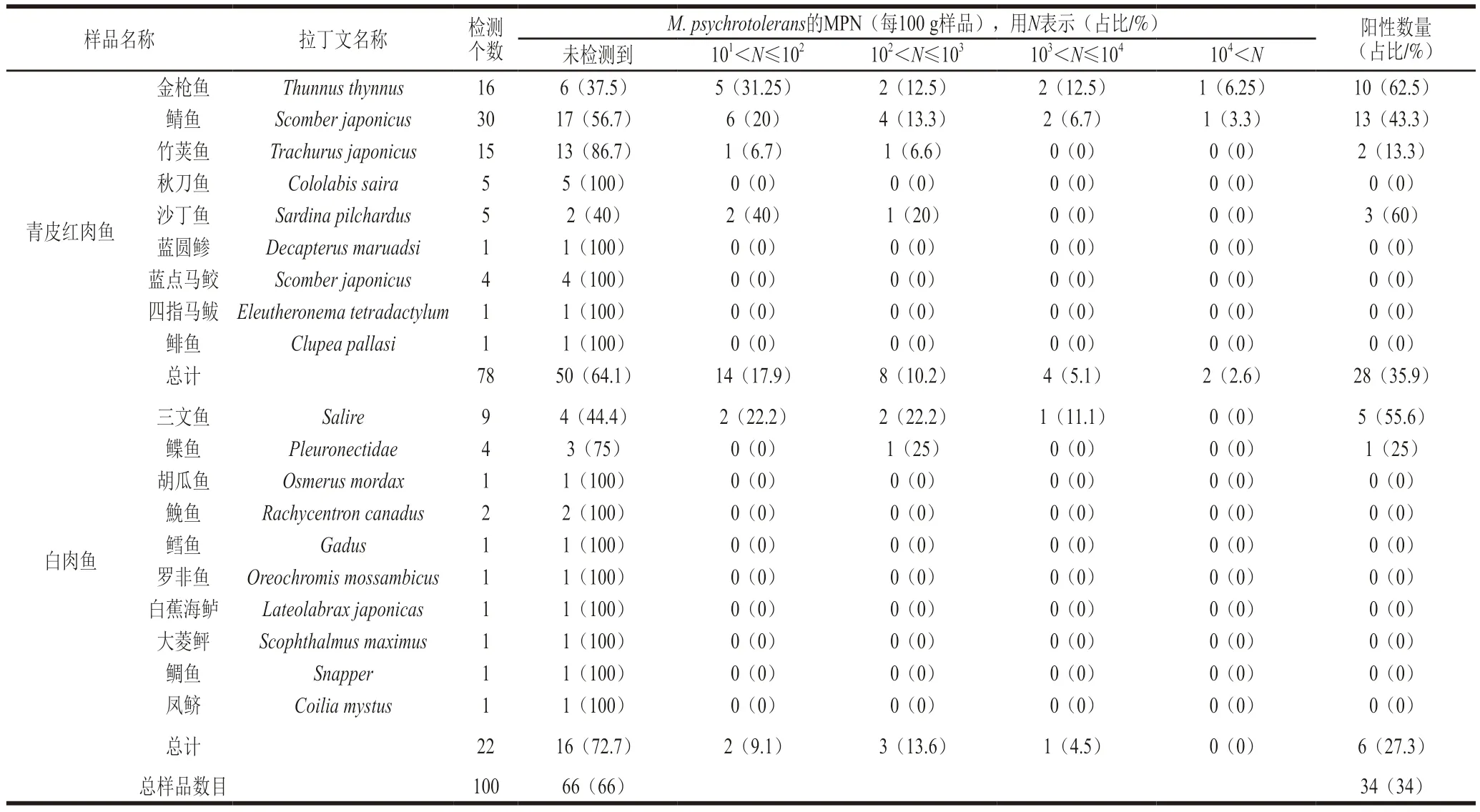
表4 魚類水產品中MPN-PCR陽性樣品數Table 4 Number of MPN-PCR positive samples in different kinds of fish
2.2 水產品中M.psychrotolerans的篩選與分離鑒定
如圖2所示,其中16 株分離株顯示出VasD特征基因條帶,同時16 株分離株生理生化實驗研究發現,分離株符合M.psychrotolerans的生理生化特征,表現為革蘭氏陰性、氧化酶陰性、過氧化氫酶陽性、苯丙氨酸酶陽性、硝酸鹽還原性、葡萄糖發酵性、D-半乳糖的非酸產生性。其中,16 株M.psychrotolerans分別來自金槍魚(1~4 分離株)、鯖魚(5~14分離株)與三文魚(15~16 分離株)。
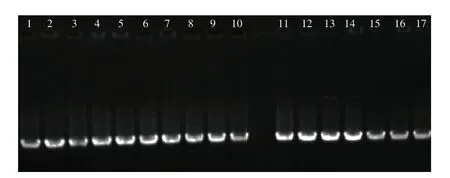
圖2 分離株VasD特征基因PCR擴增瓊脂糖凝膠電泳Fig.2 Agarose gel electrophoresis of PCR amplified products of the characteristic gene VasD from isolated strains
2.3 M.psychrotolerans分離株的組胺生成能力
對分離到的16 株M.psychrotolerans進行生長與組胺生成能力的測定。如圖3所示,在TSBH培養基中經20 ℃(室溫)培養48 h后,菌落總數均上升至9(lg(CFU/mL))左右,但未顯示出顯著差異(P>0.05),對其組胺生成量進行研究發現,分離菌株經20 ℃培養48 h后的組胺生成量在1903~4281 mg/L之間,分離株1呈現最強組胺生成能力,分離株13呈現出最弱組胺生成力,但分離株組胺生成量均超過1000 mg/L,符合Staruszkiewicz等[13]對于高組胺生成菌的定義。不同菌株之間組胺生成量呈現顯著差異。分離株M.psychrotolerans1的組胺生成量為4281 mg/L,略高于模式菌株的組胺生成量(4147 mg/L)。
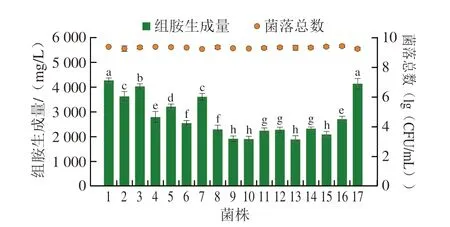
圖3 M.psychrotolerans菌株生長及組胺生成能力Fig.3 Growth and histamine-producing capacity of M.psychrotolerans
2.4 M.psychrotolerans分離株1與模式菌株生長和組胺生成能力
將分離株1與M.psychrotoleransJCM16473T模式菌株在4 ℃(冷藏)和20 ℃(室溫)條件下的生長曲線和組胺生成能力進行比較研究,結果如圖4所示。在4 ℃條件下培養時,分離株1 的初始接菌量約為3.9(lg(CFU/mL)),模式菌株的初始接菌量約為3.2(lg(CFU/mL))。在培養2 d后,兩株菌的菌落總數上升至4.5(lg(CFU/mL));培養4 d后,菌落總數上升至7(lg(CFU/mL))左右(圖4a),且檢測到了組胺的生成,分離株1的組胺生成量為208.4 mg/L,模式菌株的組胺生成量為197.2 mg/L(P>0.05)(圖4b);在培養8 d后,菌落總數雖未呈現快速上升,但菌株組胺積累量呈現顯著上升,分離株1與模式菌株組胺生成量分別為2531.57 mg/L與2177.67 mg/L;培養10 d后,分離株1與模式菌株組胺生成量分別上升至4224.17 mg/L與3960 mg/L,分離株1的組胺生成量顯著高于模式菌株(P<0.05)。在20 ℃(室溫)條件下,分離株1與模式菌株生長曲線呈現相似趨勢(圖4c),培養12 h后檢測到組胺生成,培養36 h后,兩株菌的組胺生成量均大于1000 mg/L,且模式菌株的組胺生成量略高于分離株1,但并未呈現顯著差異(P>0.05);當培養至48 h,分離株1組胺生成量顯著高于模式菌株分離株1(P<0.05);在培養結束時,模式菌株組胺生成量上升至3937.67 mg/L,顯著高于分離株1(P<0.05)(圖4d)。
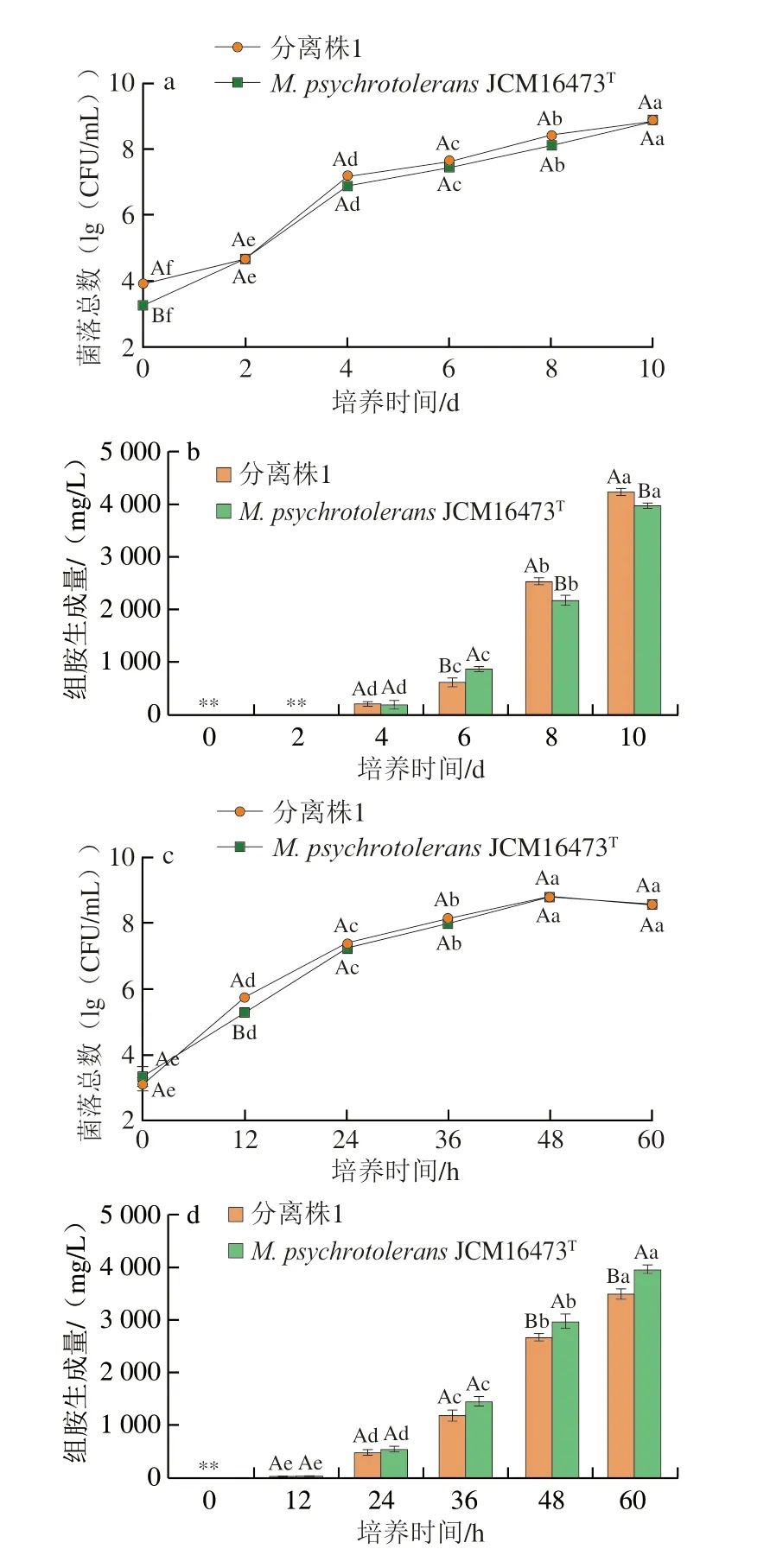
圖4 不同溫度條件下分離株1與模式菌株在TSBH中的生長和組胺生成情況Fig.4 Growth and histamine-producing capacity of isolate 1 and type strain in TSBH at different temperatures
3 討論
水產品是優質蛋白的重要來源之一,其高蛋白質、低熱量、高不飽和脂肪酸的特點深受消費者喜愛,是居民膳食結構的重要組成部分。我國是水產品生產和消費大國,居民對水產品的消費比重不斷增加,但水產品易發生品質劣化且存在食品安全風險,其中組胺作為影響水產品質量安全的重要因素之一。隨著冷鏈物流的發展,冷鏈已可以解決由中溫性組胺生成菌引起的組胺積累的問題,但由耐冷性組胺生成菌引起的組胺積累問題并未減少,即使在冷鏈物流發達的美國與日本亦是如此。M.psychrotolerans作為2006年才被分離出的耐冷性組胺生成菌,研究發現其具有較強的組胺生成能力,并且能夠在0 ℃條件下生長并生成組胺,且廣泛存在于日本在售水產品中,其在北海道水產中的污染率達42.6%,在青皮紅肉魚和白肉魚中的污染率分別為31.6%和62.5%[25]。與本實驗研究結果進行總結比較發現,M.psychrotolerans在北海道在售水產品與廣東省在售魚類水產品中污染率均達到30%以上。此外,Bjornsdottir-Butler等[31]從美國阿拉巴馬州當地超市的真空包裝的鬼頭刀魚中也分離出了M.psychrotolerans;Frith等[32]在研究環境和水質變量對墨西哥灣北部組胺生成菌濃度和種類的影響中發現,M.psychrotolerans占所有分離菌株的1%。以上研究結果均表明M.psychrotolerans廣泛存在海水與水產品中。因此,需更多關注由M.psychrotolerans引起的水產品中組胺積累問題。
為了更好地調查M.psychrotolerans在廣東市售水產品中的污染分布情況,本研究利用MPN-PCR法在12 ℃條件下進行實驗,該溫度可以有效抑制中溫性組胺生成菌的生長[25],同時在培養基中添加硫酸黏菌素,可以有效抑制其他微生物的生長,但由于M.psychrotolerans具有硫酸黏桿菌抗生素抗性[20],M.psychrotolerans可在此培養基中很好地生長。此外,M.psychrotolerans因與M.morganii具有高度相似性,本研究利用PCR法對M.psychrotolerans的特征基因VasD進行擴增,有效地區分了M.morganii與M.psychrotolerans。本研究采用MPN-PCR法可高效實現對水產品中M.psychrotolerans污染調查。本研究目前分離16 株M.psychrotolerans,其中14 株均來自青皮紅肉魚類水產品,占全部分離株的87.5%,分析其主要原因是由于青皮紅肉魚中組氨酸含量高于白肉魚,約為幾千到幾萬 μg/g[33],水產品中組胺含量與水產品中組氨酸含量呈現正相關。14 株分離株又均來自金槍魚與鯖魚,主要可能由于金槍魚與鯖魚在青皮紅肉魚中其魚肉中組氨酸含量也較高,含量約為24000 μg/g和13500 μg/g,是FDA重點關注的具有組胺中毒風險的水產品;此外,本研究在三文魚中也分離到2 株M.psychrotolerans,分析其原因可能是由于M.psychrotolerans不僅為組胺生成菌,同時也是三文魚致病菌[34]。金槍魚、鯖魚、三文魚作為全球流通的水產品,廣泛存在于全球各國與各個城市的水產品市場中,研究結果提示需要重點關注這些水產品,同時對分離出M.psychrotolerans的水產品開展溯源調查研究,明確M.psychrotolerans主要污染源頭。
本研究中16 株M.psychrotolerans分離株均顯示出較強的組胺生成能力,且分離株組胺生成能力不盡相同(圖3、4),這與Kato等[22]的研究結果一致,其分離到的菌株也與模式菌株呈現出不同的組胺生成能力。這可能是由于組胺生成與多個酶相關,包括組氨酸脫羧酶、轉運組胺和組氨酸的相關蛋白的酶和編碼組織蛋白酶RNA合成酶[35-37],經過對比NCBI現有的M.psychrotolerans全基因組測序結果發現,編碼不同M.psychrotolerans菌株的組胺生成相關基因存在不同,這可能引起與組胺生成相關酶活力呈現不同;同時組氨酸轉運與組胺轉出還與菌株細胞膜蛋白相關,這些原因均可能引起分離株組胺生成能力存在不同。在4 ℃培養2 d后的分離株1和模式菌株均未檢測到組胺的生成(圖4b),這主要是因為細菌在新環境中的生長早期階段,通常會優先利用營養物質進行生長和繁殖,而不是產生大量的代謝產物,并且在較低的溫度下,微生物的生長和代謝活動都有所減緩,組胺的積累主要發生在微生物生長的對數生長后期與平穩期[38];在4 ℃培養4 d后模式菌株和分離株1的組胺生成量分別達到了197.2 mg/L和208.4 mg/L,均超過了FDA規定的組胺質量濃度安全標準(50 μg/mL),在北海道金槍魚和沙丁魚等水產品中分離到的M.psychrotolerans在4 ℃培養8 d組胺生成量達到了2800 mg/L[22],接種了丹麥分離的M.psychrotolerans的金槍魚罐頭在4 ℃培養6 d,組胺生成量超500 mg/kg[24]。結合本研究與文獻中報道中的研究結果表明,M.psychrotolerans廣泛存在于全球水產品中且低溫下具有高組胺生成能力的組胺生成菌。
4 結論
本研究針對M.psychrotolerans首次開展了其在水產品中污染情況的調查,研究發現M.psychrotolerans在廣東省市售水產品中廣泛存在且其分離株均具有較強組胺生成能力。本研究結果表明需重點關注水產品中的M.psychrotolerans控制由其可能帶來的水產品中組胺積累的問題,開發新型抑菌劑與抑菌技術有效控制M.psychrotolerans,同時開展耐冷性產胺菌生物胺生成與積累機制研究。本研究為水產品中M.psychrotolerans污染情況提供理論數據,對切實保障水產品品質與質量安全具有重要科學意義。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
