TLR4/NF-κB p65高表達對皮膚鱗狀細胞癌增殖、侵襲和轉移的影響
2024-04-23 02:49:06劉莉
中華養生保健 2024年8期
劉 莉
(寧夏醫科大學總醫院腫瘤研究所,寧夏 銀川,750001)
皮膚鱗狀細胞癌簡稱皮膚鱗癌,屬于惡性腫瘤,在國內外均較為常見。該疾病來源于皮膚的鱗狀上皮組織,屬于非黑色素瘤皮膚癌[1-2]。近些年,由于環境污染與人口老齡化的原因,皮膚癌的發病率也隨之升高,有研究顯示,在我國皮膚惡性腫瘤中,皮膚鱗狀細胞癌的比例占到80.0%,且發病率仍在上升,對人類生命安全造成嚴重的威脅[3]。皮膚鱗癌的病因至今仍未明確,可能與紫外照射損傷、人乳頭瘤病毒、免疫抑制、砷角化病、放射性皮炎、黏膜白斑、瘢痕和白化病等有關[4-5]。皮膚鱗癌的發生僅次于基底細胞癌,占非黑色素瘤皮膚癌的20%。與基底細胞癌不同,皮膚鱗癌具有高度的侵襲能力,因此容易發生轉移和復發[6]。早期的皮膚鱗癌可以通過手術切除,但是對發生轉移的皮膚鱗癌患者的長期預后效果很差,1年的生存率為50%,5年的生存率僅有5%,而對于復發的患者,5年的生存率也不到10%[7]。核因子KappaB(NF-κB)存在于人體內,屬于重要的轉錄因子,在抑制腫瘤細胞增殖、促進細胞凋亡中發揮作用,NF-κBp65是常見的NF-κB類型[8]。近年來研究證明,在皮膚鱗狀細胞癌中,NF-KBp65蛋白呈現出低表達狀態,而且當病情加重時,其表達水平也隨之降低[9]。Toll樣受體4(Toll-like recepTor 4,TLR4)不僅可以識別胞外抗原,對胞內損傷相關因子也起到應答作用,進而促使細胞分泌炎性因子和干擾素[10]。本研究具體探討與分析了TLR4/NF-κB p65高表達對皮膚鱗狀細胞癌增殖、侵襲和轉移的影響,以明確TLR4/NF-κB p65的作用機制,現報道如下。
1 材料與方法
1.1 材料來源
研究時間為2020年2月—2023年2月,自武漢普諾賽生命科技有限公司購買人皮膚鱗狀細胞癌A431細胞株,細胞培養于含10.0%胎牛血清(生產企業:杭州四季青生物公司)、100 U/ml青霉素(生產企業:上海生物工程有限公司)、0.1 mg/ml鏈霉素(生產企業:上海生物工程有限公司)的DMEM低糖液體培養液(生產企業:美國InviTrogene公司)中,超表達載體-pc3.0-TLR4NF-κB p65與空載體pc3.0由本實驗室保存,LipofecTamine 2 000購自美國InviTrogene公司。Transwell小室、MTT試劑盒、AnnexinV-FITC試劑盒都購自碧云天公司,抗Caspase-9蛋白、抗Myc蛋白購自美國InviTrogene公司。
1.2 細胞轉染
A431細胞在37 ℃、5%CO2條件下培養,實驗時取對數生長期細胞。空白組不進行轉染,加入等量的DMEM培養基;對照組的轉染物質為pc3.0空載體,實驗組的轉染物質為pc3.0-TLR4NF-κB p65載體。
1.3 TLR4/NF-κB p65 RNA表達檢測
細胞轉染后24 h、48 h,收獲細胞,并提取總RNA(方法為:Trizol法),使用qRT-PCR方法檢測mTLR4/NF-κB p65的表達水平,配制20μL體系,行定量PCR檢測,每個反應3個復孔。PCR條件:95 ℃3 min,35個循環:95 ℃12 s,56 ℃ 50 s。
1.4 MTT檢測細胞增殖活性
取對數期生長期的轉染后24 h、48 h的細胞,行胰酶消化,隨后取細胞,使用新鮮培養基重懸細胞,以1×103/孔接種細胞到96孔板中,將其放入37 ℃孵箱中培養。培養12 h后加入MTT溶液10μl/孔,再將其放入37 ℃孵箱,培養時長為1 h。用酶標儀測定450 nm測定OD值,計算細胞增殖活性。
1.5 Transwell小室檢測細胞侵襲和轉移指數
轉染24 h、48 h后,將細胞進行鋪板,位置:Transwell小室里的上層、下層加入DMEM+10.0%胎牛血清培養基600 μl,把Transwell小室放進24孔板里,培養24 h后取出,多聚甲醛固定并用結晶紫染色,選擇5個隨機視野進行細胞計數,計算細胞侵襲指數與細胞轉移指數。
1.6 Western blot法檢測Caspase-9、Myc蛋白表達水平
轉染24 h、48 h后,收集細胞并加入細胞裂解液,充分裂解后以12 000 r/min離心,定量蛋白濃度后進行SDS-PAGE凝膠電泳,電泳結束后進行轉膜,采用5.0%的脫脂奶粉進行封閉。加入抗Caspase-9蛋白、抗Myc蛋白,稀釋濃度分別為1:500與1:1 000,洗膜后加入二抗,室溫孵育1 h,洗膜后用化學發光劑顯色,使用凝膠成像儀,觀察目的蛋白條帶表達情況,以β-acTin作為內參蛋白,計算目的蛋白的相對表達水平。
上述實驗均進行3次,取平均值。
1.7 統計學分析
選擇SPSS 22.0軟件對本研究所有的計量數據進行統計學分析,描述計量數據為(),兩組間比較行t檢驗,多組間比較行方差分析。P<0.05代表差異有統計學意義。
2 結果
2.1 TLR4/NF-κB p65 RNA表達水平
轉染24 h、48 h后,實驗組的TLR4、NF-κB p65 RNA表達水平均較對照組與空白組高,差異有統計學意義(P<0.05),空白組與對照組比較,差異無統計學意義(P>0.05),見表1。
表1 三組轉染不同時間點后的TLR4/NF-κB p65 RNA表達水平比較 ()
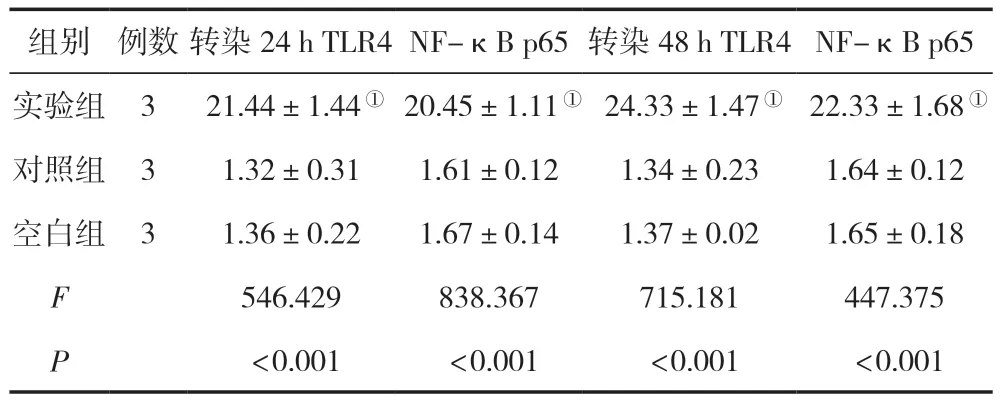
表1 三組轉染不同時間點后的TLR4/NF-κB p65 RNA表達水平比較 ()
注:與對照組相比,①P<0.05。
組別例數轉染24 h TLR4NF-κB p65 轉染48 h TLR4NF-κB p65實驗組3 21.44±1.44① 20.45±1.11① 24.33±1.47① 22.33±1.68①對照組31.32±0.311.61±0.121.34±0.231.64±0.12空白組31.36±0.221.67±0.141.37±0.021.65±0.18 F 546.429838.367715.181447.375 P<0.001 <0.001 <0.001 <0.001
2.2 細胞增殖指數
轉染24 h、48 h后,實驗組的細胞增殖指數顯著低于空白組與對照組,差異無統計學意義(P<0.05),對照組與空白組比較,差異無統計學意義(P>0.05),見表2。
表2 三組轉染不同時間點后的細胞增殖指數比較 (,%)

表2 三組轉染不同時間點后的細胞增殖指數比較 (,%)
注:與對照組相比,①P<0.05。
組別例數24 h48 h實驗組3 45.87±3.14① 56.69±4.21①對照組3 78.20±13.2989.20±4.39空白組378.44±2.8789.60±3.57 F 18.624 24.932 P<0.001<0.001
2.3 細胞侵襲指數與細胞轉移指數
轉染24 h、48 h后,在細胞侵襲指數與細胞轉移指數方面,實驗組均較空白組、對照組低,差異無統計學意義(P<0.05),對照組與空白組比較,差異無統計學意義(P>0.05),見表3。
表3 三組轉染不同時間點的細胞侵襲指數與細胞轉移指數比較 (,%)
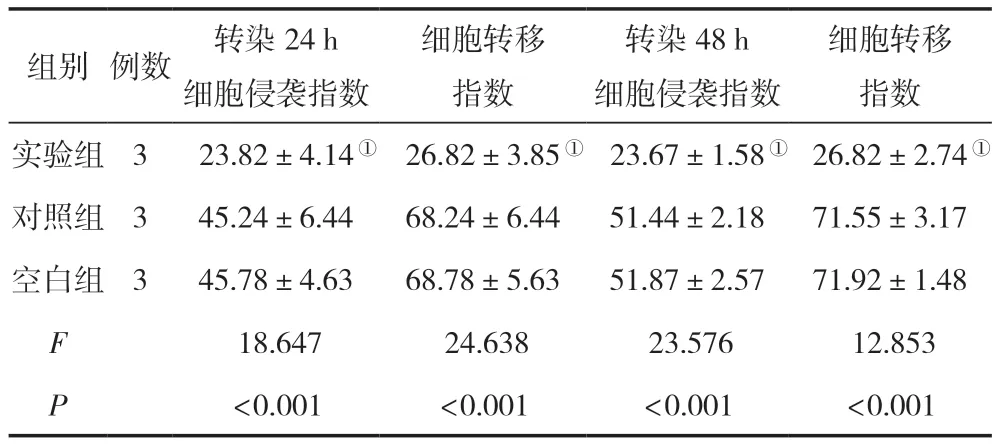
表3 三組轉染不同時間點的細胞侵襲指數與細胞轉移指數比較 (,%)
注:與對照組相比,①P<0.05。
細胞轉移指數實驗組3 23.82±4.14① 26.82±3.85① 23.67±1.58① 26.82±2.74①對照組345.24±6.4468.24±6.4451.44±2.1871.55±3.17空白組345.78±4.6368.78±5.6351.87±2.5771.92±1.48 F 18.647 24.638 23.576 12.853 P<0.001<0.001<0.001<0.001組別例數轉染24 h細胞侵襲指數細胞轉移指數轉染48 h細胞侵襲指數
2.4 Caspase-9/Myc蛋白表達水平
轉染24 h、48 h后,實驗組的Caspase-9/Myc蛋白相對表達水平與空白組、對照組相比顯著增高,差異有統計學意義(P<0.05),空白組與對照組比較,差異無統計學意義(P>0.05),見表4。
表4 三組轉染不同時間點后的Caspase-9/Myc蛋白表達水平比較 ()
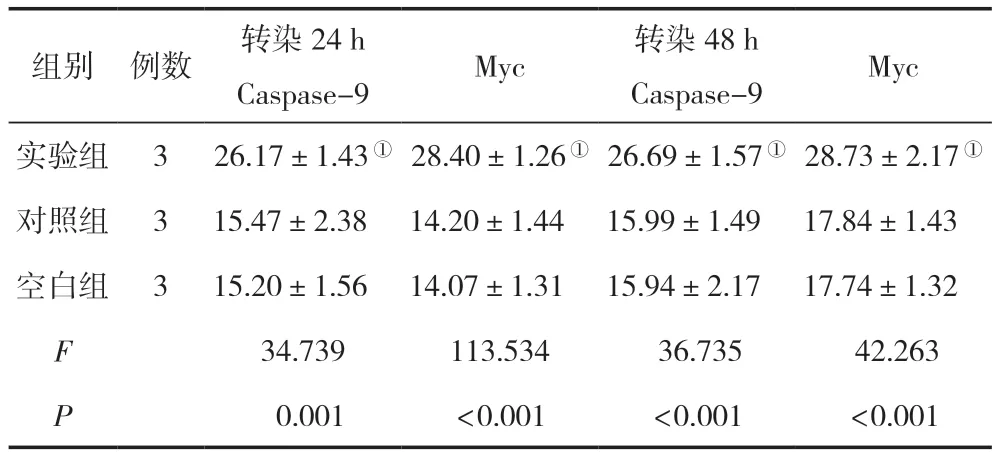
表4 三組轉染不同時間點后的Caspase-9/Myc蛋白表達水平比較 ()
注:與對照組相比,①P<0.05。
組別例數轉染24 h Caspase-9Myc 轉染48 h Caspase-9Myc實驗組326.17±1.43①28.40±1.26①26.69±1.57①28.73±2.17①對照組3 15.47±2.38 14.20±1.44 15.99±1.49 17.84±1.43空白組3 15.20±1.56 14.07±1.31 15.94±2.17 17.74±1.32 F 34.739113.534 36.735 42.263 P 0.001<0.001<0.001<0.001
3 討論
當前皮膚鱗狀細胞癌對人們健康的威脅日益嚴重,雖然當前醫療技術顯著提高,但患者的5年病死率一直居高不下[11]。至今為止,該疾病的發病機制仍不明確,涉及的病因包括炎性反應因子、外傷、人乳頭瘤狀病毒感染、長期暴露于不良環境、免疫抑制狀態等[12]。當前皮膚鱗狀細胞癌的發生具有逐漸上升的趨勢,尤其是在老年人中的發病率逐年增高,嚴重威脅著人類的健康[12]。本研究結果顯示,轉染24 h、48 h后,實驗組的TLR4、NF-κB p65 RNA表達水平較對照組、空白組高(P<0.05),空白組與對照組比較無明顯差異(P>0.05);實驗組的細胞增殖指數顯著低于空白組與對照組(P<0.05),對照組與空白組比較無明顯差異(P>0.05),表明TLR4/NF-κB p65高表達能抑制皮膚鱗狀細胞癌細胞的增殖。從機制上分析,Toll樣受體在固有免疫及獲得免疫過程中發揮著重要作用,是哺乳類動物細胞表面的跨膜受體[13]。TLR4是Toll樣受體的主要成員之一,可以識別各種與損傷有關的分子模式(damage-associaTedmolecularpaTTern,DAMPs)、病原有關分子模型(pathogen-associated molecular paTTern,PAMP),可激活下游NF-κB p65的表達,引起多種炎性反應介質的釋放,從而介導調節機體的免疫反應與炎性反應。TLR4/NF-κB p65高表達可調節腫瘤浸潤免疫作用,可抑制腫瘤細胞增殖[14-15]。
皮膚鱗狀細胞癌的發生、發展是多步驟、多因素共同作用的結果,在非黑色素瘤皮膚癌中,該疾病的發病率處于第二位,病死率一直比較高。當前研究顯示,TLR4介導炎性反應信號通路的持續激活,還可調控腫瘤細胞增殖活性,TLR4信號通路的活化可抑制多種惡性腫瘤的異常增殖[16]。TLR4在皮膚鱗狀細胞癌中的高表達可促進炎性反應,抑制腫瘤惡化[17]。本研究結果顯示,轉染24 h、48 h后,實驗組的細胞侵襲指數與細胞轉移指數都明顯低于空白組與對照組(P<0.05),對照組與空白組對比差異無統計學意義(P>0.05),表明TLR4/NF-κB p65高表達能抑制皮膚鱗狀細胞癌細胞侵襲與轉移。從機制上分析,TLR4是Toll樣受體家族亞型中最重要的成員,不僅廣泛表達于免疫原性細胞,也表達于多種軀體惡性腫瘤與腦腫瘤細胞中,與多種腫瘤的生物學行為密切相關。TLR4在發揮抗腫瘤作用時,可通過T細胞的方式,TLR4在誘導抗腫瘤免疫應答時,可通過Myd88依賴性途徑間接誘導,從而實現腫瘤抵抗作用[18]。TLR4還可通過活化多種信號途經激活Fas介導的途徑,可抑制腫瘤細胞的增殖,也可促進腫瘤細胞的凋亡[19]。
皮膚鱗狀細胞癌的發生、發展與多種信號通路失調有關,在皮膚鱗狀細胞癌中有多個蛋白表達水平發生異常。特別是在腫瘤的發生與蛋白酶級聯反應等相關程序的啟動與激活密切相關,因而參與細胞凋亡過程。細胞凋亡等相關級聯反應主要是由Caspase蛋白家族介導執行,其中Caspase-9、Myc作為凋亡啟動因子,非常重要,對其表達水平進行檢測后,其結果可作為細胞凋亡發生情況的間接反映。本研究結果顯示,轉染24 h、48 h后,實驗組的Caspase-9/Myc蛋白相對表達水平與空白組、對照組相比顯著增高(P<0.05),空白組與對照組相比差異無統計學意義(P>0.05),表明TLR4/NF-κB p65高表達能促進皮膚鱗狀細胞癌細胞的Caspase-9/Myc蛋白表達水平,從而發揮促進細胞凋亡的作用。上調TLR4的表達可將c-Myc和K-Ras相關通路激活,將促進Caspase-3、Caspase-9的表達,從而促進腫瘤細胞的凋亡[20]。不過本研究也有一定的不足,實驗分組比較少,尤其是沒有進行劑量分析,也沒有進行相應的臨床與動物模型分析,將在之后的研究中進行深入分析。
綜上所述,TLR4/NF-κB p65高表達能抑制皮膚鱗狀細胞癌細胞的增殖、侵襲,也可促進Caspase-9/Myc蛋白的表達,從而發揮促進腫瘤細胞凋亡的作用。
