a-硫辛酸在微囊藻毒素-LR誘導草魚卵巢細胞損傷中的作用
2024-11-06 00:00:00王輝何麗阮記明梁惜梅李福貴隗黎麗
農業環境科學學報 2024年3期

關鍵詞:草魚卵巢細胞;a-硫辛酸;微囊藻毒素-LR;氧化應激;炎癥反應
微囊藻毒素(MicrocVstins,MCs)是淡水水體有毒藍藻產生的一類藻毒素,目前已有超過279種微囊藻毒素異構體,其中微囊藻毒素-LR(MicrocVstin-LR,MC-LR)是毒性最大、研究最廣泛的一種異構體。研究表明MC-LR具有顯著的肝毒性,它通過抑制絲氨酸一蘇氨酸磷酸酶1和2A而引起蛋白質磷酸化增加,導致細胞骨架結構的改變。此外,研究表明,氧化應激在MCs誘導毒性作用中也起著重要作用,它們可以誘導細胞內活性氧(Reactive oxygen species,ROS)的形成,氧化和破壞細胞大分子,導致組織和器官的損傷。目前,MCs的氧化毒性機制已引起廣泛的關注,一些學者嘗試使用天然或化學抗氧化劑來減少MCs誘導的組織或器官的損傷,例如,Gehringer等研究發現維生素E對MC-LR誘導的小鼠肝臟有保護作用,其他學者還發現姜黃素、褪黑素、N-乙酰半胱氨酸、水飛薊素、巖藻多糖和大蒜等可顯著降低MC-LR誘導的肝臟氧化損傷。
MC-LR除了誘導肝毒性之外,還可在生殖系統中蓄積并產生生殖毒性。研究發現,MC-LR可破壞哺乳動物睪丸、卵巢、前列腺和胎盤等器官的結構和功能,從而降低哺乳動物的生殖能力;同時,MC-LR的毒性還可以通過胎盤傳給后代,造成后代死亡、畸形、生長遲緩以及器官功能障礙等。在魚類中,MC-LR能影響斑馬魚的內分泌和卵子發生,并干擾卵母細胞減數分裂,從而對斑馬魚的生殖造成不利影響,同時其還能在魚類的卵巢和精巢中積累并傳遞給后代。卵巢由于富含不飽和脂質而很容易受到氧化損傷。Hou等研究發現卵巢的抗氧化防御系統可能是斑馬魚對抗MC-LR毒性作用的重要機制。因此,有學者對抗氧化劑降低MC-LR誘導的生殖毒性做了一些探索性的工作。例如,N-乙酰半胱氨酸可以降低MC-LR誘導的中國倉鼠卵巢細胞(CHO)和C57BL/6小鼠卵巢的氧化損傷,但這方面的研究還比較少。
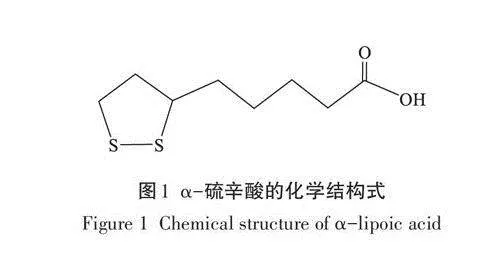
a-硫辛酸(Alpha lipoic acid, a-LA)是一種含有1個二硫鍵的天然有機硫化物(圖1),屬于B族維生素,兼具水溶性和脂溶性的性質,極易被多種組織器官吸收利用。此外,a-LA對內源性的抗氧化劑如維生素C、維生素E以及谷胱甘肽(Glutathione,GSH)等具有再生作用,從而可提高機體的抗氧化能力。同時,a-LA還可通過提高抗氧化酶的活性從而發揮抗氧化功能,如飼料中添加a-LA可顯著提高皺紋盤鮑體內的超氧化物歧化酶(Superoxide dis-mutase.SOD)活性以及GSH含量。a-LA作為一種強大的抗氧化劑,可有效提高機體的抗氧化能力,從而降低MC-LR的毒性。也有一些研究表明,a-LA能有效降低MC-LR對魚類肝臟、大腦和肌肉的毒性,并通過激活核因子相關因子2(Nuclear factorervthroid 2-related factor 2,Nrf2)再生GSH來保護MC-LR誘導的肝毒性。但目前還沒有報道a-LA在MC-LR誘導的生殖毒性方面的保護作用。草魚(Ctenopharyngodon idella)作為我國四大家魚之一,是我國重要的淡水養殖魚類,但目前關于MC-LR對草魚生殖的影響還未見報道,更沒有相關研究報道a-LA在MC-LR誘導的草魚卵巢(Grass carp ovary,GCO)細胞毒性中的作用。結合相關文獻報道,我們推測a-LA可通過抑制MC-LR誘導GCO細胞氧化應激從而保護細胞。因此,本研究以GCO細胞為研究對象,探討a-LA在MC-LR致GCO細胞毒性中的保護作用,并為MC-LR生殖毒性的防治提供新的理論依據。
1材料與方法
1.1試驗細胞
試驗所用GCO細胞是中國科學院水生生物研究所在20世紀70年代開展草魚m血病研究時建立的一株細胞系,由江西師范大學付建平老師保存惠贈。
1.2主要儀器與試劑
細胞培養所用的C02培養箱為Hengzi公司產品。冷凍離心機和NANODROP2000超微量分光光度計為Thermo公司產品。MULTISKANFC全波段掃描酶標儀和CFX96/384 Touch實時熒光定量PCR儀為Bio-Rad公司產品。MC-LR(純度gt;98%)與a-LA分別購自Expreess Technology與Sigma公司。胎牛血清、M199培養基、0.25%胰蛋白酶購自Biological Indus-tries公司。青鏈霉素混合液、二甲基亞砜(DMSO)、1×PBS、噻唑蘭(Methylthiazolyldiphenyl-tetrazolium bro-mide,MTT)檢測試劑盒為Solarbio公司的產品。提取RNA的TRIzol Reagent試劑盒購自Invitrogen公司。合成cDNA的PrimeScript RT reagent Kit With gDNAEraser試劑盒與用于qRT-PCR的SYBR Premix ExTaprMⅡ試劑盒均購自Takara公司。乳酸脫氫酶(Lac-tic dehydrogenase, LDH)、GSH、丙二醛(Malondialde-hvde,MDA)測定試劑盒購自南京建成生物工程研究所有限公司,二喹啉甲酸法(Bicinchoninic Acid Assay,BCA)蛋白濃度測定試劑盒購自Bevotime公司。
1.3 GCO細胞復蘇及培養
從-80℃冰箱中取出凍存的GCO細胞,置于26℃水浴鍋中水浴溶解,將溶解的GCO細胞懸液用基礎培養基重復清洗兩次,隨后加入完全培養基重懸細胞,并將細胞轉移到培養皿或培養瓶中,在條件為26℃、5% C02的培養箱中進行培養,并取對數生長期的細胞用于后續試驗。
1.4 MTT法測定細胞活力
取對數生長期的GCO細胞,以1.2×105 ind·mL-1的濃度接種到96孔細胞培養板中,設置不同MC-LR濃度(0、5、10、20、40、60umol·L-1)和a-LA濃度(0、62.5、125、250、500、1000、2000umol`L-1)的完全培養基處理GCO細胞。每個濃度設5個平行,處理24h后,采用MTT法分別分析MC-LR和a-LA對GCO細胞活力的影響。MTT法簡單操作步驟如下:在避光條件下每孔加入20uL MTT溶液,于培養箱內孵育4h,小心吸出上清液,避免吸到藍紫色結晶,隨后每孔加入150uL DMSO溶液,搖勻后,孵育10min,充分溶解結晶,最后采用酶標儀測定各孔在570nm處的吸光度值(OD),計算細胞活力。根據吸光度值采用SPSS 22.0軟件計算得到MC-LR 24h的半數增殖抑制濃度(Half maximal inhibitory concentration,IC50)約為48umol·L-1,隨后選擇12umol·L-1(IC50/4)和24umol·L-1 (IC50/2) MC-LR作用于GCO細胞并進行形態觀察,發現24umol·L-1 MC-LR組細胞內出現自噬體、細胞質內出現較多的空泡、線粒體發生腫脹、內質網擴張等,因此,選擇1/2 IC50即24umol-L-1MC-LR為后續試驗的染毒濃度,相關信息參考本實驗室前期發表的文章。
1.5試驗分組及處理
取對數生長期的GCO細胞,以1.2x105ind·mL-1的濃度接種到100mm培養皿中,待細胞貼壁生長至80%左右后,根據1.4活力測定的MC-LR和a-LA對GCO細胞活力影響的結果,設置空白對照組(不添加MC-LR和a-LA)、a-LA組(125umol·L-1a-LA)、MC-LR組(24umol·L-1MC-LR)以及a-LA+MC-LR組(125umol·L-1 a-LA+24umol·L-1 MC-LR),每個處理設置3個重復。細胞培養24h后取樣分析。
1.6 LDH活性和抗氧化指標的測定
由1.5分組處理結束后,采用PBS溶液洗兩遍,離心收集細胞,加入200uL PBS重懸細胞,利用超聲儀超聲破碎細胞使細胞內成分釋放,離心10min(12000r·min-1),收集上清。LDH活性以及組織中MDA和GSH含量均按照試劑盒說明書嚴格操作測定,測定GSH及MDA含量時,采用BCA法測定蛋白含量。
1.7引物設計
根據NCBI基因庫已有的草魚基因全長序列,使用Primer Premier 5軟件設計實時熒光定量(qRT-PCR)的引物,其中內參基因GAPDH的選用參考本實驗室前期已發表的文章。引物由上海生物工程有限公司合成,試驗所用引物如表1所示。
1.8RNA提取及qRT-PCR
根據1.5中的分組方案將GCO細胞處理24 h后,PBS溶液清洗細胞兩次,收集細胞于無RNA酶的離心管。使用Trizol法提取細胞總RNA,再利用微量分光光度計測定RNA的純度和濃度。然后嚴格按照PrimeScript RT reagent Kit with gDNA Eraser試劑盒使用說明合成cDNA。將合成的cDNA稀釋5倍,再根據SYBR Premix Ex TapⅡ試劑盒使用說明配制10uL qRT-PCR反應體系,每個樣品設置3個重復孔,采用Bio-Rad CFX熒光定量PCR儀進行反應,以GAPDH為內參基因,采用2法計算各基因在GCO細胞中的相對表達量。
1.9統計學處理
本試驗的所有結果均以平均數±標準差(Mean±SD)表示。使用SPSS 22.0軟件對數據進行單因素方差分析(One-ANOVA),運用Tukey檢驗進行組間多重比較,其中Plt;0.05表示組間差異顯著,具有統計學意義。
2結果與分析
2.1a-LA對GCO細胞活力的影響
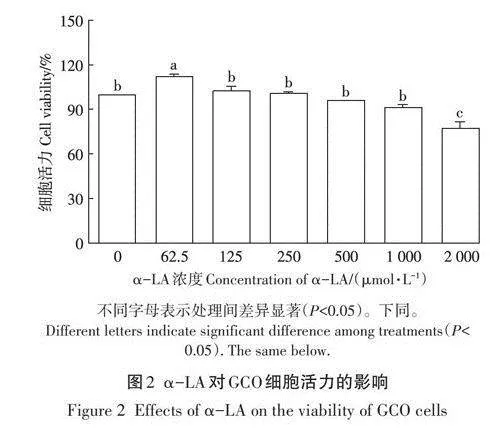
a-LA作用24 h對GCO細胞活力的影響結果如圖2所示,與不添加a-LA組相比,隨著a-LA濃度的升高,細胞活力呈現先上升后下降的趨勢。62.5pmol·L-1a-LA可顯著提高細胞活力(Plt;0.05),而當a-LA濃度為125-1000umol·L-1時,細胞活力無顯著變化(Pgt;0.05),當a-LA的濃度上升至2000umol·L-1時,細胞活力顯著下降(Plt;0.05)。
2.2a-LA對MC-LR誘導GCO細胞活力的影響
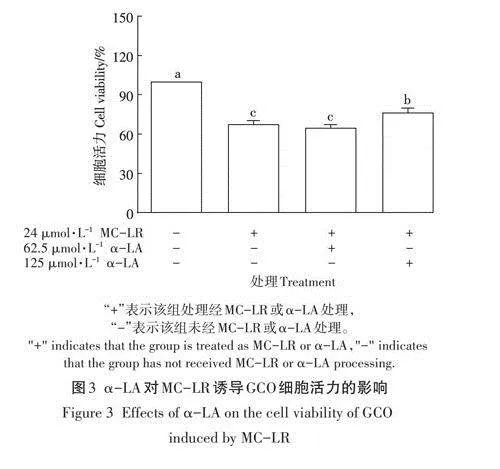
如圖3所示,與對照組相比,MC-LR單獨處理和MC-LR聯合不同濃度a-LA處理均能夠顯著降低細胞活力(Plt;0.05).而與MC-LR組相比,125umol·L-1a-LA+MC-LR組細胞活力顯著上升(Plt;0.05),說明該濃度的a-LA對MC-LR致GCO細胞毒性具有較好的保護效果。因此,后續選用125umol·L-1的a-LA進行試驗。
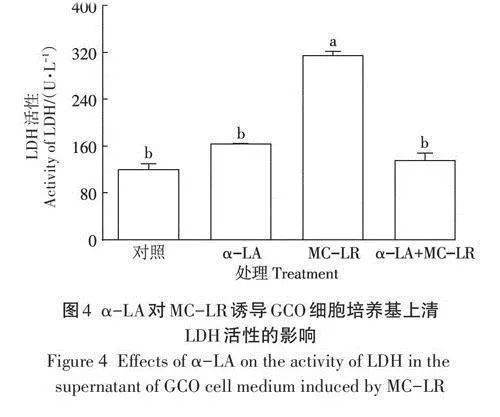
2.3 a-LA對MC-LR誘導GCO細胞培養基上清LDH活性的影響
GCO細胞培養基上清中LDH的活性結果如圖4所示,與對照組相比,a-LA組LDH活性無顯著變化(Pgt;0.05),MC-LR組LDH活性顯著上升(Plt;0.05)。而與MC-LR組相比,a-LA+MC-LR組LDH活性顯著下降(Plt;0.05)。
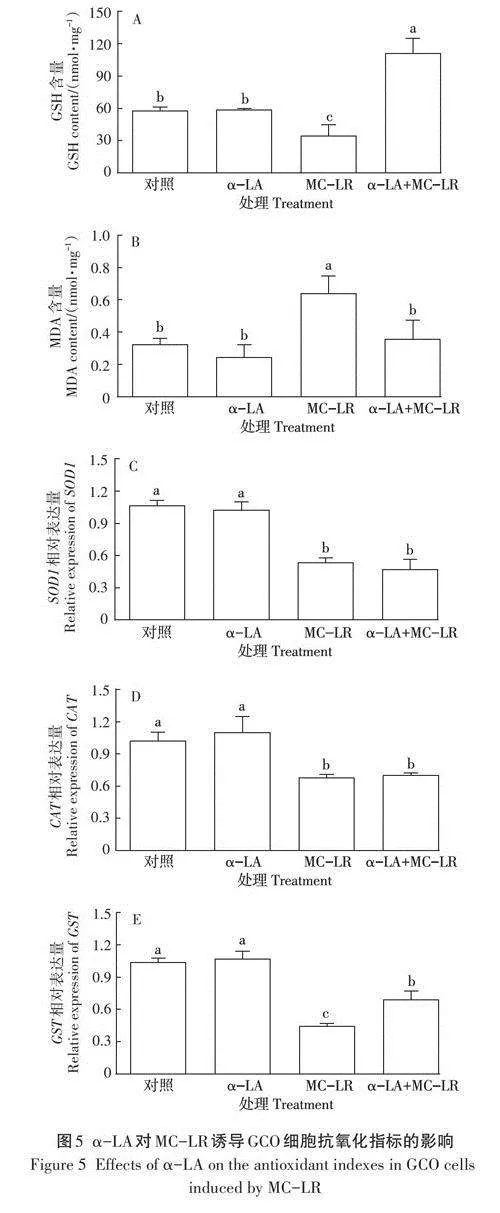
2.4 a-LA對MC-LR誘導GCO細胞抗氧化指標的影響
如圖SA所示,與對照組相比,GCO細胞內GSH相對含量在MC-LR組顯著下降(Plt;0.05),而在a-LA+MC-LR組中其含量顯著上升(Plt;0.05)。與MC-LR組相比,a-LA+MC -LR組GCO細胞內GSH相對含量顯著上升(Plt;0.05)。由圖SB可知,MC-LR組GCO細胞內MDA相對含量與各組相比均顯著上升(Plt;0.05),且在對照組、a-LA組、a-LA+MC-LR組間無顯著變化。
如圖5C、圖5D、圖5E所示,GCO細胞內SOD1、CAT、GST基因相對表達量在對照組與a-LA組間均無顯著變化(Pgt;0.05);相對于對照組與a-LA組,MC-LR組、a-LA+MC-LR組的SOD1、CAT、GST基因相對表達量均顯著下降(Plt;0.05);相對于MC-LR組,a-LA+MC-LR組的.SOD1、CAT基因相對表達量無顯著變化(Pgt;0.05),GST基因相對表達量顯著上升(Plt;0.05)。
2.5a-LA對MC-LR誘導GCO細胞炎癥相關基因表達的影響
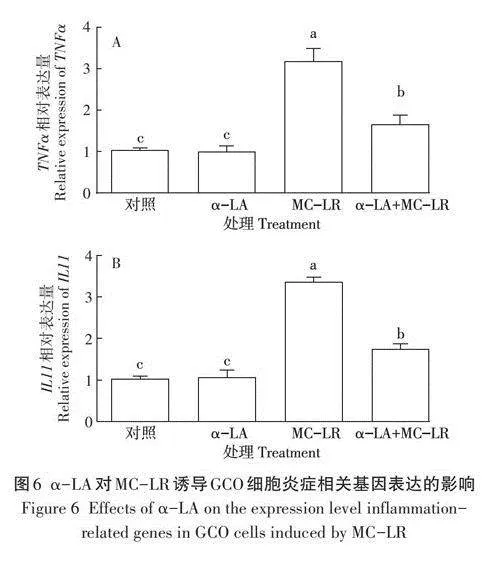
a-LA對MC-LR誘導GCO細胞炎癥相關基因表達的影響結果如圖6所示,與對照組相比,a-LA組中TNFa基因的相對表達量無顯著變化(Pgt;0.05),與MC-LR組相比,a-LA+MC -LR組中TNFa基因的相對表達量顯著下降(Plt;0.05,圖6A)。與對照組相比,a-LA組中基因的相對表達量無顯著變化(Pgt;0.05),與MC-LR組相比,a-LA+MC-LR組中基因的相對表達量顯著下降(Plt;0.05,圖6B)。
3討論
卵巢是雌性動物的重要生殖器官,可以產生卵子和類固醇性激素。急性暴露于MC-LR已被證明會導致斑馬魚卵巢的病理損傷,并引發氧化應激。本實驗室前期研究也發現MC-LR可誘導GCO細胞產生氧化應激。本研究旨在確定a-LA是否對MC-LR誘導的GCO細胞氧化損傷具有緩解作用。
a-LA作為一種天然的抗氧化劑,含有1個二硫鍵,其一旦進入細胞,二硫鍵就被還原為二氫硫辛酸(DHIA),并與蛋白質氨基酸殘基的氨基結合形成酰胺鍵,酰胺鍵可經脂酰胺酶作用裂解釋放具有生理功能的a-LA。研究表明a-LA在保護機體免受氧化損傷方面具有巨大的潛力,因此其在臨床研究以及生產實踐中得到廣泛應用。有研究表明,a-LA可以顯著提高缺氧狀態下大鼠皮層神經元細胞的活力,并且a-LA可顯著提高鎘脅迫下大鼠肝細胞活力。本研究發現a-LA可顯著提高MC-LR作用后的細胞活力(Plt;0.05)。此外,LDH是無氧酵解和糖異生的重要酶系之一,在體內能量代謝過程中發揮重要作用,幾乎存在于所有組織中,細胞損傷均可導致LDH濃度增加。進行離體細胞培養時,如果培養的細胞受損,LDH也會由細胞漏m至培養液中。本試驗結果表明,a-LA+MC-LR組LDH活性較MC-LR組顯著下降(Plt;0.05),這與其他人的研究結果類似.如Zhang等研究發現,a-LA能降低缺氧誘導的臍靜脈內皮細胞中LDH的釋放。因此,本研究結果表明a-LA能夠緩解MC-LR導致的GCO細胞損傷。
MDA被認為是評價機體氧化應激最重要的標志物之一,是來源于機體內不飽和脂肪酸被氧自由基攻擊而形成的過氧化產物,而GSH是機體內重要的抗氧化物,可被GST催化并與機體內有害的親電基團結合,從而清除體內的脂質過氧化物。有研究顯示,MCs能消耗GSH造成的細胞氧化應激和脂質過氧化(Lipid peroxidation,LPO),產生過氧化產物如MDA等,從而損傷細胞器,繼而引起內質網應激。也有研究發現小鼠經腹腔注射MC-LR后,其肝臟GSH水平發生時間依賴性變化,且谷胱甘肽過氧化物酶(Glutathione peroxidase,GPx)和谷胱甘肽還原酶(Glu-tathione reductase.GR)等的活性發生改變。Amado等的研究表明,a-LA可提高鯉魚腦和肝臟中GST的活性從而抵抗MC-LR誘導的機體損傷。Gu等的研究表明,a-LA可提高HepG2和Be17402細胞內GSH的生成以及SOD的活性,同時降低ROS的生成和MDA的含量,從而減弱MC-LR對細胞的損傷。還有研究發現在飼料中添加a-LA可提高鯽魚肝臟GPx和CAT活性,并提高機體的總抗氧化能力,降低MDA含量,從而減輕MC-LR對魚體的氧化損傷。在本研究中,相對于對照組,MC-LR可以顯著升高GCO細胞內MDA含量,并顯著降低GSH含量,這說明MC-LR可以誘導GCO細胞發生氧化應激。與MC-LR組相比,a-LA+MC-LR組GCO細胞內GSH含量顯著上升,且MDA含量顯著下降,說明a-LA可能通過升高機體內GSH含量而增強機體抗氧化能力,以此降低MC-LR造成GCO細胞氧化應激而升高MDA含量。但在本研究中,a-LA+MC-LR組的CAT和SOD1基因表達水平相對于MC-LR組并沒有顯著差異,僅上調了CST基因的相對表達水平。因此推測本試驗中a-LA主要通過增加GCO細胞內GSH的含量,并促進GST的表達,從而促進GSH清除MC-LR誘導的自由基,最終降低細胞內MDA的生成,以此來緩解MC-LR引起的GCO細胞氧化損傷,這與Pflugmacher等的研究結果類似。
ILII是一種與炎癥及免疫疾病相關的多功能細胞因子,也具有抗炎作用。有研究表明,炎癥是刺激ILII表達的因素之一,當機體受到感染、損傷和炎癥反應時,ILII基因表達會增加。TNFa也是一種炎性細胞因子,可與受體TNFR2結合介導細胞激活NF-KB信號通路,進而抑制細胞凋亡或促進細胞炎癥的發生,所以當機體發生炎癥時,通常會促進TNFa的表達。有研究表明MC-LR能通過誘導血管內皮細胞氧化應激,致使TNFa的表達升高,從而誘導炎癥反應。在本研究中MC-LR同樣可導致ILII和TNFa基因表達水平的升高,表明MC-LR誘導了GCO細胞反應的發生。有趣的是,有研究發現a-LA同樣具有緩解機體內炎癥損傷的作用。如Costa等研究發現,a-LA可降低IL6和TNFa水平,抑制藥物誘導小鼠炎癥的發生,從而提高小鼠的存活率。此外,a-LA還可作為抗炎劑抑制小鼠IL-1和TNFa蛋白的表達【46]。本試驗結果與上述研究結果相似,表明a-LA能通過下調GCO細胞中TNFa和ILII基因的表達來抑制炎癥的發生,從而減弱MC-LR對GCO細胞的損傷。
到目前為止,人們盡管嘗試了維生素E、姜黃素、褪黑素、N-乙酰半胱氨酸、水飛薊素、巖藻多糖和大蒜等天然或化學抗氧化劑來降低MC-LR誘導的肝臟、腎臟、心臟等組織或器官毒性,但仍然處于實驗室研究階段。與其他抗氧化劑相比,a-LA最獨特的特點是它可以同時與脂質和水溶性化合物反應。研究表明a-LA在體內外實驗中均展示了其可緩解MC-LR誘導的肝毒性,也有文獻表明a-LA可作為抗MC-LR誘導的肝臟氧化損傷的潛在藥物。本研究所有結果也表明a-LA可緩解MC-LR誘導的生殖細胞毒性,然而,本研究缺乏體內生殖毒性實驗的數據,對a-LA誘導的生殖毒性的具體調控機制也未開展研究,在今后的研究中,將重點對該方面進行深入探討。
4結論
a-硫辛酸作為抗氧化劑具有很大的潛力,可以提高草魚卵巢細胞經微囊藻毒素-LR誘導后的抗氧化能力,并通過緩解氧化應激反應的發生,抑制炎癥反應,從而減弱微囊藻毒素-LR對草魚卵巢細胞的損傷。因此,a-硫辛酸對預防微囊藻毒素-LR誘導的生殖毒性有一定的作用。
猜你喜歡
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中西醫結合心血管病電子雜志(2016年21期)2017-03-03 04:50:02
中外醫學研究(2016年36期)2017-03-01 05:33:24
中國醫學創新(2017年1期)2017-02-23 14:41:26
中國實用醫藥(2016年15期)2016-05-24 03:57:32
中國實用醫藥(2016年9期)2016-05-17 11:38:42
中國實用醫藥(2016年8期)2016-03-30 00:14:55
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
