小麥新種質普冰3228籽粒礦質元素QTL定位與候選基因分析
2024-11-07 00:00:00王健勝王二偉馬愛鋤侯桂玲程世平
江蘇農業科學 2024年18期
摘要:小麥籽粒礦質元素含量對人體健康具有重要影響。為解析小麥新種質普冰3228礦質元素遺傳機制并為小麥礦質元素遺傳改良提供新材料。以小麥新種質普冰3228與京4839雜交產生的重組自交系群體為材料,利用55K SNP芯片對小麥籽粒Ca、Mg、Fe、Zn這4種元素含量進行了QTL定位及候選基因分析。結果表明,普冰3228×京4839 RIL群體4種礦質元素均存在豐富的遺傳差異;共檢測到與礦質元素相關的QTL 15個,這些QTL主要分布于1A、1D、3D、5A、5B、6B和7D染色體上,QTL LOD值介于2.58~35.89之間,解釋變異率分布于0.16%~12.30%之間,其中包括與Zn元素含量相關的QTL 4個,與Ca元素含量相關的QTL 4個,與Mg元素含量相關的QTL 7個。進一步分析發現,與籽粒礦質元素含量相關的候選基因18個,這些候選基因主要與小麥礦質元素的吸收、轉運、累積等功能相關。該結果將為小麥籽粒礦質元素遺傳機制深入解析及遺傳改良提供有用的信息和豐富的基因資源。
關鍵詞:小麥;普冰3228;礦質元素;QTL定位;候選基因
中圖分類號:S512.103.2 文獻標志碼:A
文章編號:1002-1302(2024)18-0026-09
收稿日期:2023-10-16
基金項目:國家自然科學基金委員會-河南省人民政府聯合基金(編號:U1804102)。
作者簡介:王健勝(1978—),男,陜西禮泉人,博士,副教授,主要從事小麥遺傳育種。E-mail:wjsheng1998@163.com。
礦質元素在人體新陳代謝中發揮著重要作用,其直接影響人體健康。近年來由于礦質元素缺乏引起的相關疾病已在臨床醫學中被發現并引起了社會的普遍關注[1-2]。例如,據估計世界范圍內有近20億人患有因鐵元素缺乏引起的貧血,這個問題在發展中國家表現尤為突出。小麥是世界主要糧食作物之一,其籽粒礦質元素含量對保障人體礦質元素正常攝入具有重要影響,為此,國際農業研究磋商組織(CCIAR)等國際機構于2003年開始實施國際性重大農業項目Harvest Plus計劃,旨在通過培育富含礦物營養成分的小麥等主要糧食作物,解決礦質營養元素缺乏問題。可見,開展小麥礦質元素研究進而提高其礦質元素含量已成為目前小麥生產的緊迫科學問題。
小麥礦質元素遺傳機制研究是遺傳改良并提升小麥籽粒礦質元素含量的重要基礎。眾所周知,小麥籽粒礦質元素含量是受多基因控制的數量性狀,而數量性狀基因座(QTL)分析為礦質元素遺傳機制解析提供了一條有效途徑。圍繞小麥籽粒礦質元素QTL定位,國內外學者開展了較多的相關研究。郝志利用雙單倍體群體對小麥籽粒Fe、Zn、Cu、Mn這4種元素含量進行了QTL定位,結果共檢測到10個QTL位點,其中Fe元素檢測到4個QTL位點,Zn元素檢測到3個QTL位點,Cu元素共檢測到3個QTL位,Mn元素只檢測到1個QTL位點,這些QTL分別位于1B、2A、5D、3D、5A、5B、1D、2B、4D和3A染色體上[3]。Wang等對小麥苗期和成株期Se元素含量進行了QTL定位分析,結果共檢測到與Se元素相關的QTL 16個,其中苗期發現7個QTL,成株期發現9個QTL,這些QTL主要L8Sxyhwa/B3C8XFcn2rdPQ==分布于1B、2B、4B、5A、5B、5D、6A和7D染色體上[4]。Wang等利用京冬8號與矮抗58產生的254個重組自交系群體,結合55K SNP芯片對小麥籽粒Zn元素和Fe元素含量進行了QTL分析,結果共檢測到與Zn元素含量相關的QTL 7個,發現與Fe元素含量相關的QTL 4個[5]。另外,也有學者利用全基因組關聯分析的方法檢測到了小麥礦質元素相關的部分QTL[6-10]。
綜上所述,雖然前人針對小麥籽粒礦質元素開展了較多的QTL定位研究,但有關小麥新種質普冰3228礦質元素的遺傳研究仍未見報道。基于此,本研究以小麥新種質普冰3228×京4839自交系群體(RIL)為材料,利用55K SNP芯片對小麥籽粒礦質元素進行了QTL定位及候選基因分析,以期為小麥礦質元素遺傳改良提供有益信息和有效材料。
1 材料與方法
1.1 研究材料
本研究材料是小麥新種質普冰3228與京4839雜交產生的重組自交系(RIL)群體,該群體共包括203個株系。普冰3228是普通小麥與冰草通過遠緣雜交獲得的遺傳穩定的新種質,其具有高產、抗病、抗旱等多種優異農藝性狀[11-13]。本研究所用材料均由中國農業科學院作物科學研究所李立會課題組提供。
1.2 小麥田間種植
2018—2019年和2019—2020年將普冰3228×京4839 RIL群體分株系在河南平頂山和陜西咸陽2個環境下種植,株系間采用隨機區組設計,每個株系種植4行,設3次重復。種植行長保持2 m,行距20 cm,株距6.7 cm,單籽粒點播。試驗地周圍均種植保護行,按照普通產田水平進行田間統一管理。待小麥成熟后,在每個株系中隨機收獲5~6株,分單株收獲并脫粒。
1.3 小麥籽粒礦質元素含量測定
收獲的小麥籽粒自然充分晾干,取一定量籽粒粉碎并過篩后,稱取0.50 g樣品放入50 mL微波消解管中,加入適量硝酸和過氧化氫,混勻后放入微波消解儀進行消解,消解液經充分過濾后用1%硝酸溶液定容至50 mL。接著利用日本島津原子吸收分光光度儀AA-6300,采用火焰-原子吸收法測定小麥種質籽粒的Ca、Mg、Fe、Zn這4種元素含量,每個小麥樣品重復測定3次,取平均值作為每個材料籽粒的元素含量。
1.4 基因型檢測
本研究采用十二烷基苯磺酸鈉(SDS)法提取基因組DNA[14],取親本和RIL群體幼苗葉片提取DNA,將提取后的DNA保存在TE中,并用瓊脂糖凝膠電泳法檢測其純度。然后將遺傳群體檢測質量合格的樣品DNA送往北京博奧晶典生物技術有限公司,由該公司基于Illumina SNP Genotyping技術測試平臺使用微珠芯片技術進行檢測,其多態性使用Genomestudio v 1.0軟件進行分析,最終獲得研究遺傳群體的有效基因型。
1.5 遺傳圖譜構建及QTL定位
基于上述獲得的定位群體的基因型,通過QTL IciMapping 4.0軟件構建群體的分子標記遺傳連鎖圖譜,接著結合籽粒礦質元素含量的表型性狀,采用完備區間作圖法(ICIM-ADD)進行QTL檢測,LOD閾值為2.0[15-16]。QTL按照通用規則命名,具體為:QTL+性狀英文縮寫+染色體。
1.6 候選基因分析
以與籽粒礦質元素含量緊密連鎖SNP標記序列為探針,以QTL定位置信區間作為候選基因的預測區間,在NCBI(https://www.ncbi.nlm.nih.gov)和ENA(https://www.ebi.ac.uk/ena)數據庫中進行Blast比對,獲得籽粒礦質元素含量相關候選基因,并對部分候選基因進行功能注釋。
2 結果與分析
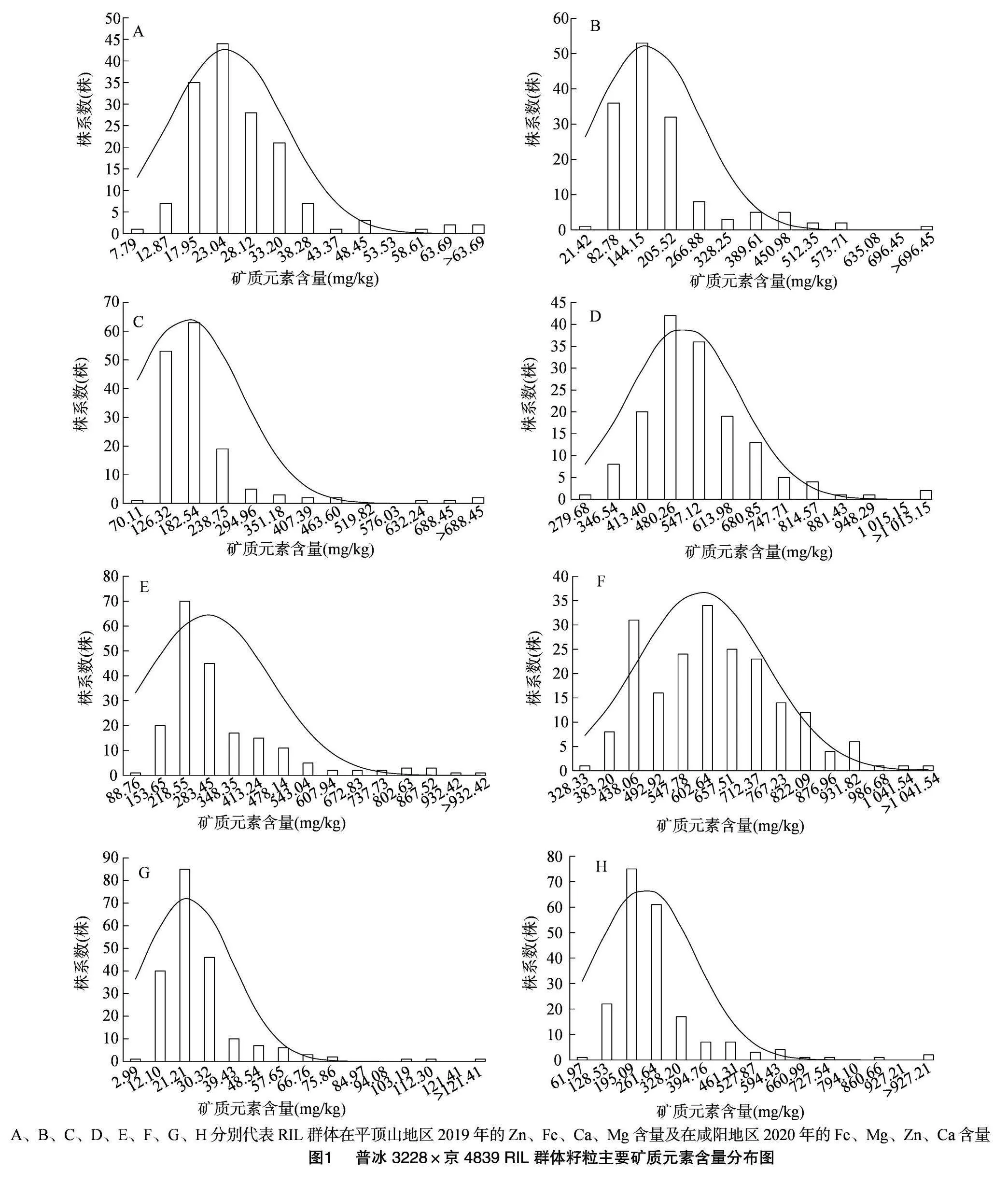
2.1 普冰3228×京4839 RIL群體籽粒主要礦質元素含量表型分析
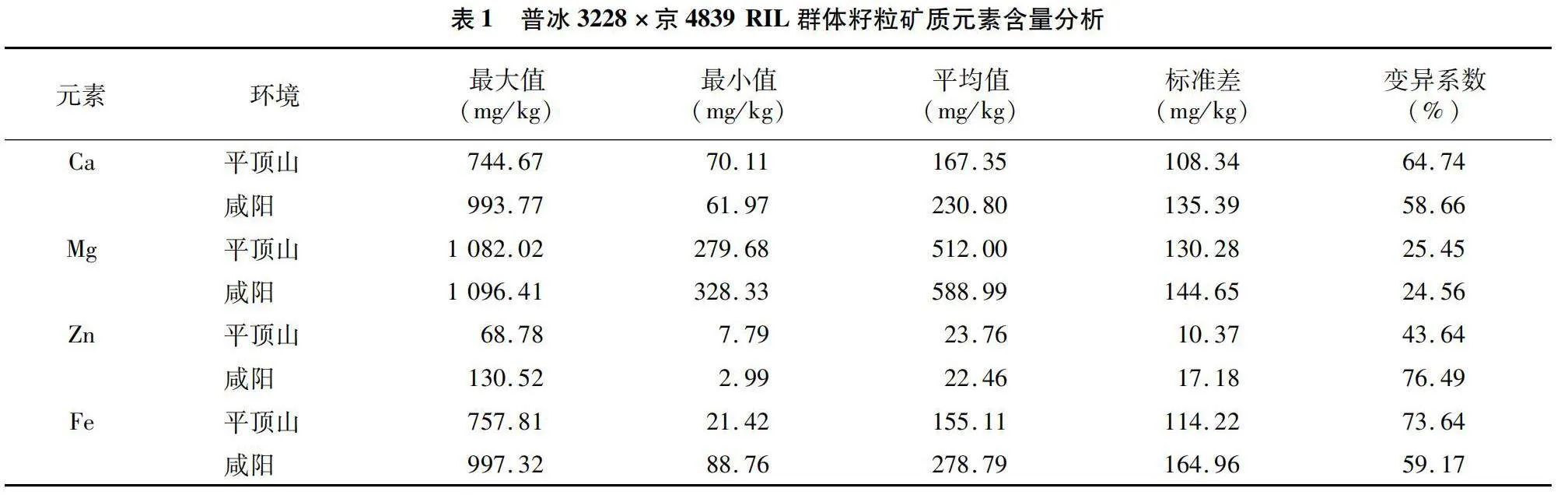
由圖1、表1可知,普冰3228×京4839 RIL群體中Ca、Mg、Fe、Zn這4種元素含量在不同環境下表現不盡相同。在籽粒Ca元素含量方面,RIL群體在平頂山地區分布在70.11~744.67 mg/kg之間,平均為167.35 mg/kg,而在咸陽地區,RIL群體分布在61.97~993.77 mg/kg之間,平均為230.80 mg/kg,可以看出,該群體籽粒Ca元素含量在咸陽環境下相對較高,但從RIL群體籽粒Ca元素含量變異程度來看,平頂山環境下變異更豐富,其變異系數為64.74%,稍高于咸陽環境下的58.66%。在籽粒Mg元素含量方面,平頂山環境下分布在279.68~1 082.02 mg/kg之間,平均為512.00 mg/kg,而在咸陽地區分布在328.33~1 096.41 mg/kg之間,平均為588.99 mg/kg,可以看出,不同環境下群體Mg元素含量較為接近。Zn元素含量在2個環境下差異較大,在平頂山地區分布在7.79~68.78 mg/kg之間,而在咸陽地區分布在2.99~130.52 mg/kg之間,但2個環境下Zn元素含量平均值較為接近,分別為23.76、22.46 mg/kg,從Zn元素含量的變異程度來看,咸陽地區較平頂山地區具有更豐富的遺傳變異。RIL群體Fe元素含量在平頂山地區最小值為21.42 mg/kg,最大值為757.81 mg/kg,平均值為155.11 mg/kg,而在咸陽地區,其最小值為88.76 mg/kg,最大值達到了997.32 mg/kg,平均值為278.79 mg/kg,Fe元素含量變異程度以平頂山地區較為豐富,其變異系數達到了73.64%。從4種元素的綜合表現來看,以Mg元素的含量為最高,其次是Fe元素和Ca元素,Zn元素的含量為最低。
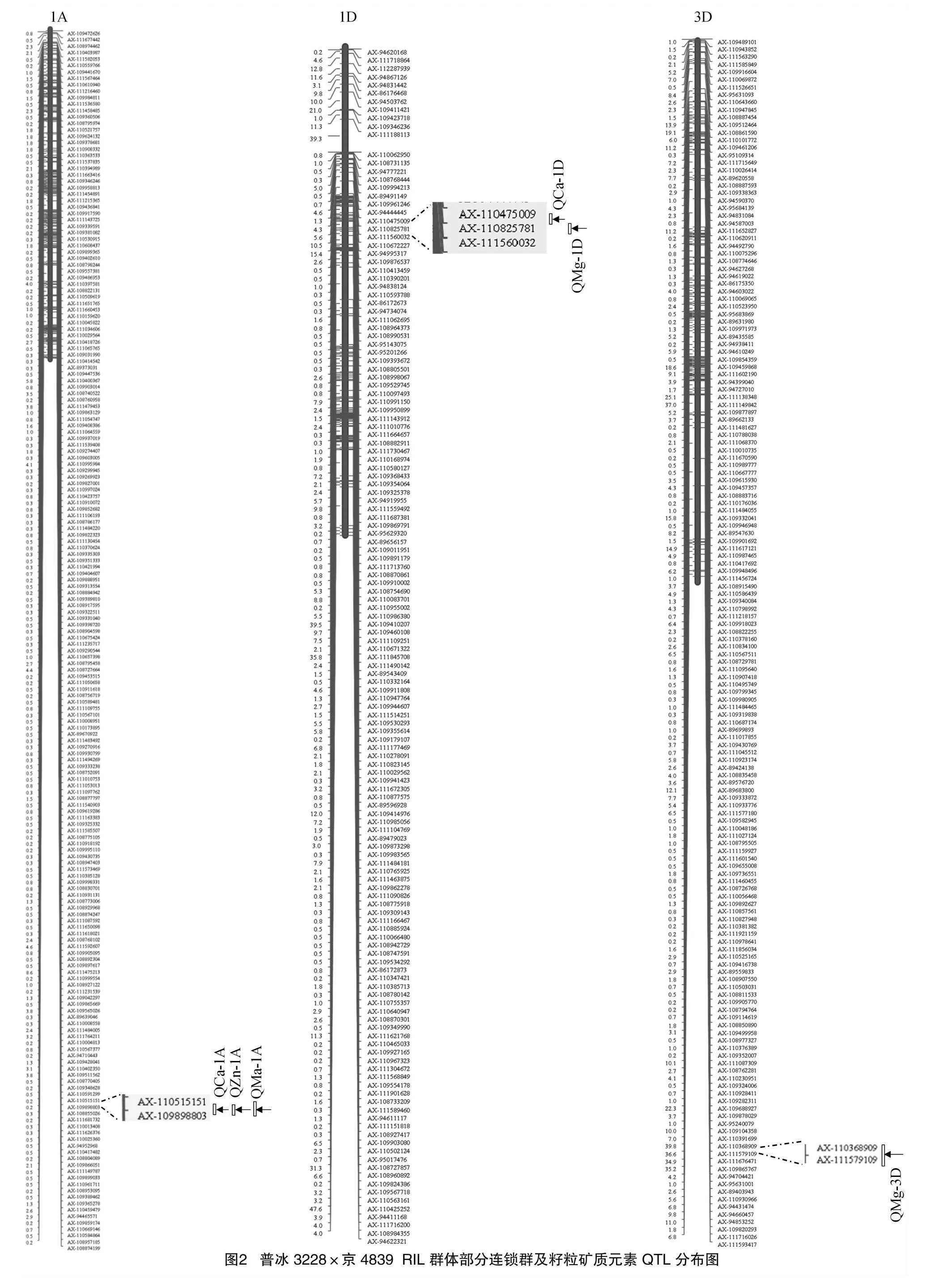
2.2 普冰3228×京4839 RIL群體主要礦質元素QTL定位
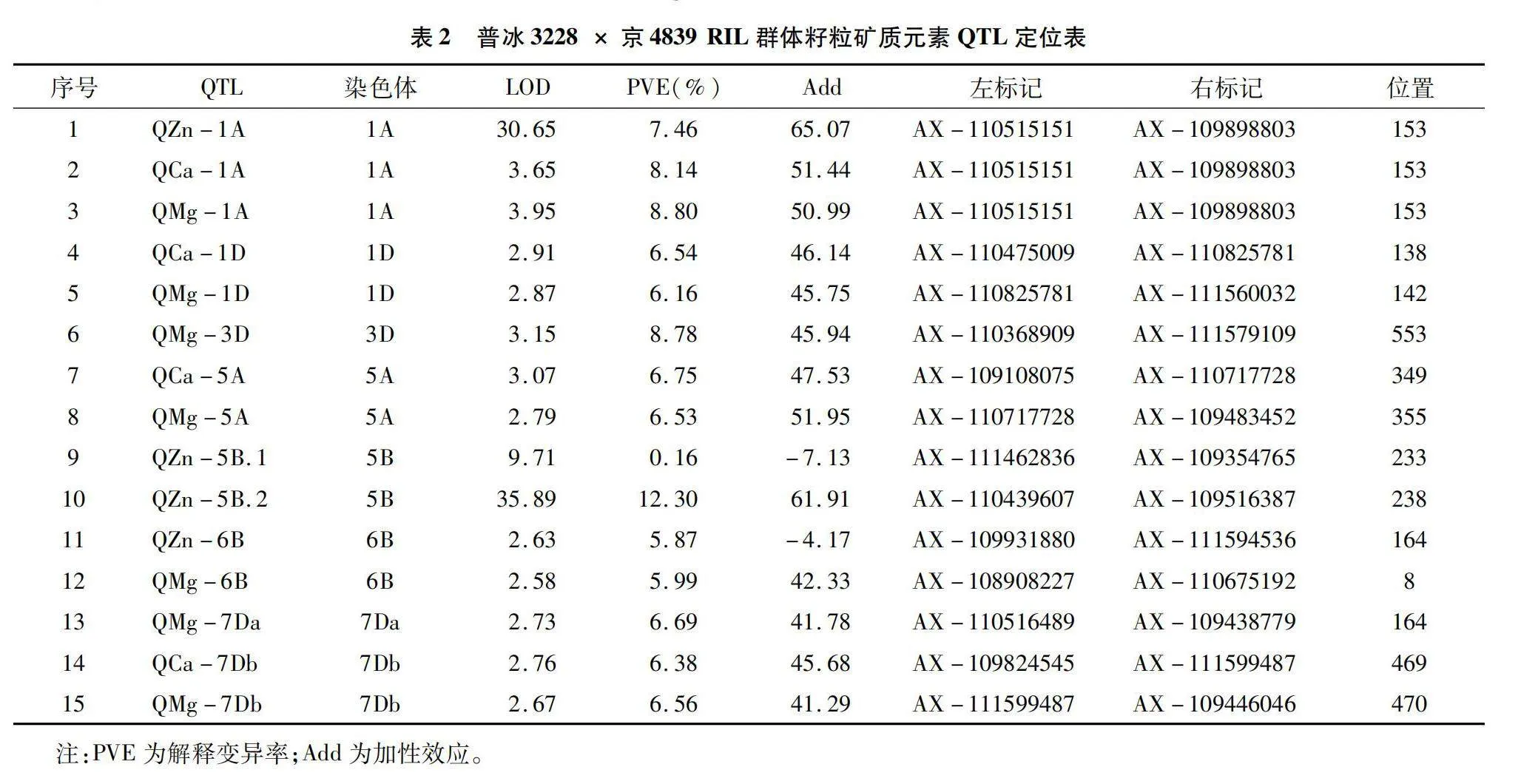
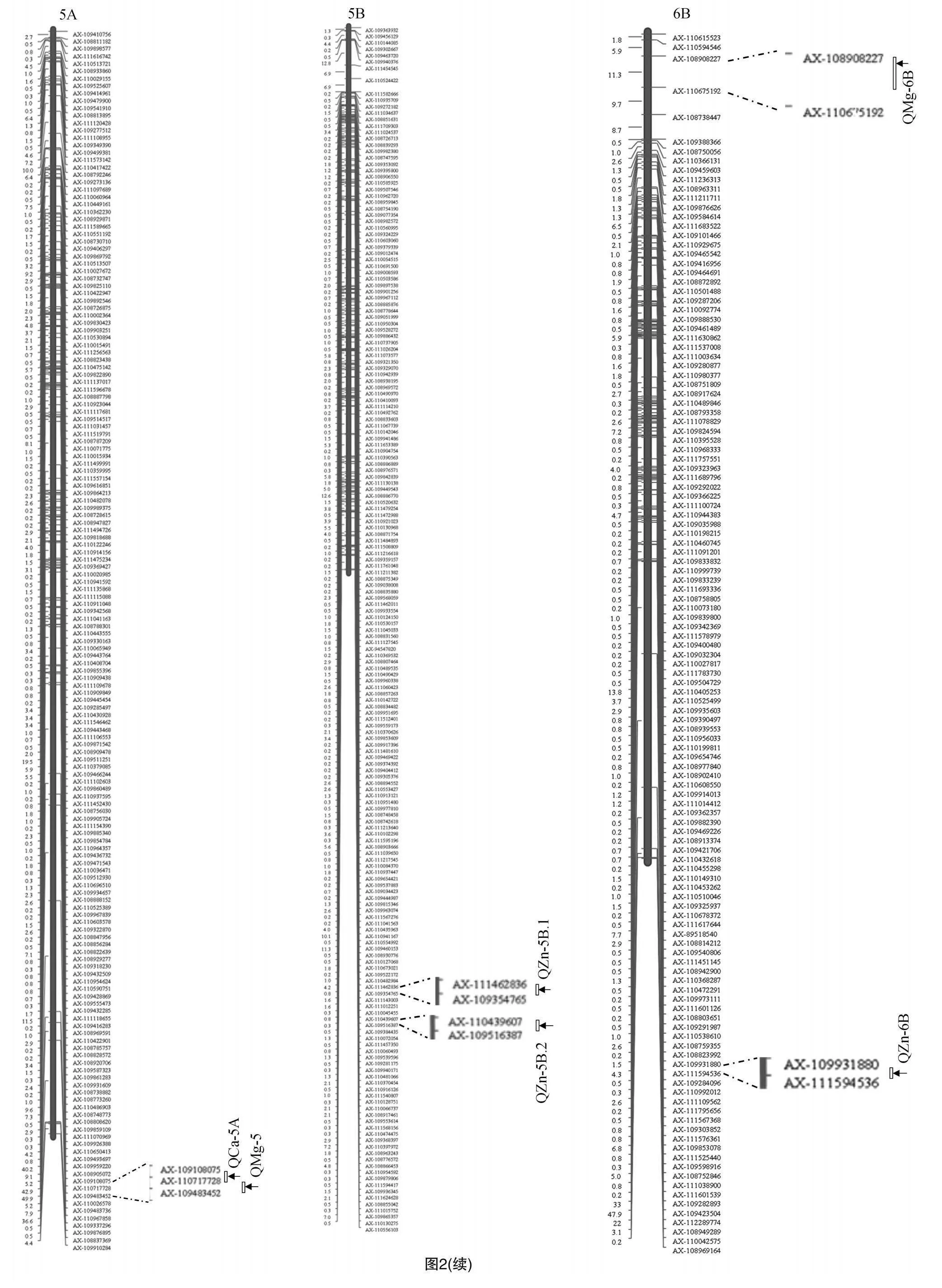
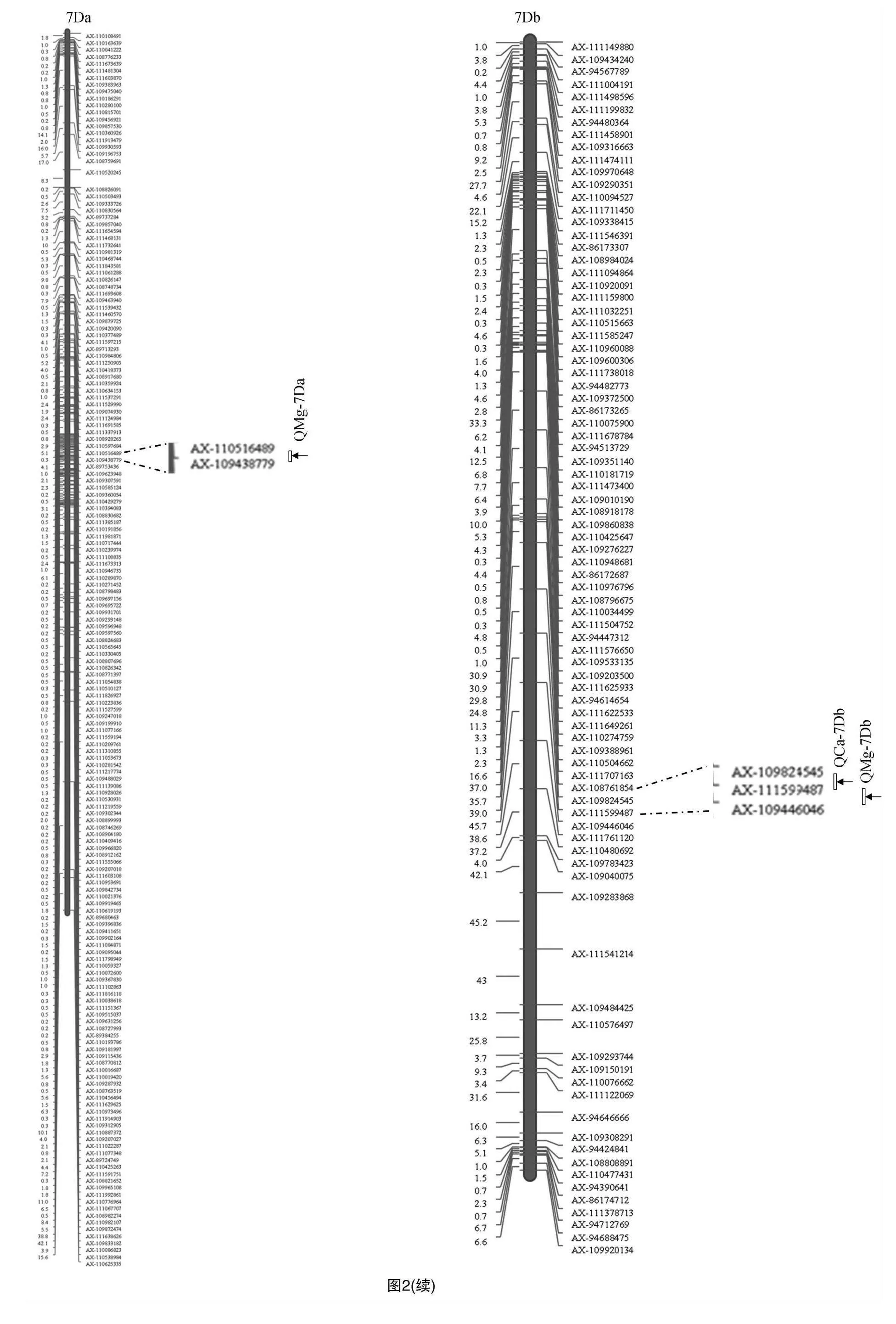
利用55K SNP芯片,本研究構建了普冰3228×京4839 RIL群體高密度遺傳連鎖圖譜,利用該圖譜結合群體不同環境下元素表型性狀對Ca、Mg、Fe、Zn這4種元素進行了QTL分析,共檢測到相關QTL 15個(表2),這些QTL主要分布于1A、1D、3D、5A、5B、6B和7D染色體上(圖2)。檢測到的QTL中包括與Zn元素相關的QTL4個,與Ca元素相關的QTL 4個,與Mg元素相關的QTL 7個,而未檢測到與Fe元素相關的QTL。檢測到的QTL LOD值介于2.58~35.89之間,解釋變異率分布在0.16%~12.30%之間,其中定位于5B染色體上AX-110439607~AX-109516387區間的控制小麥籽粒Zn含量的QZn-5B.2表現最為突出,其LOD值達到了35.89,解釋變異率為12.30%。從QTL的加性效應來看,除定位于5B、6B控制Zn含量的2個QTL為負值外,其余QTL的加性效應均為正值,這表明絕大多數QTL都來自普冰3228。同時可以看出,檢測到的QTL在小麥不同基因組間分布比較均勻,在A基因組上發現5個QTL,在B基因組上發現4個QTL,而在D基因組上發現的QTL最多,為6個。
2.3 普冰3228×京4839 RIL群體主要礦質元素候選基因分析
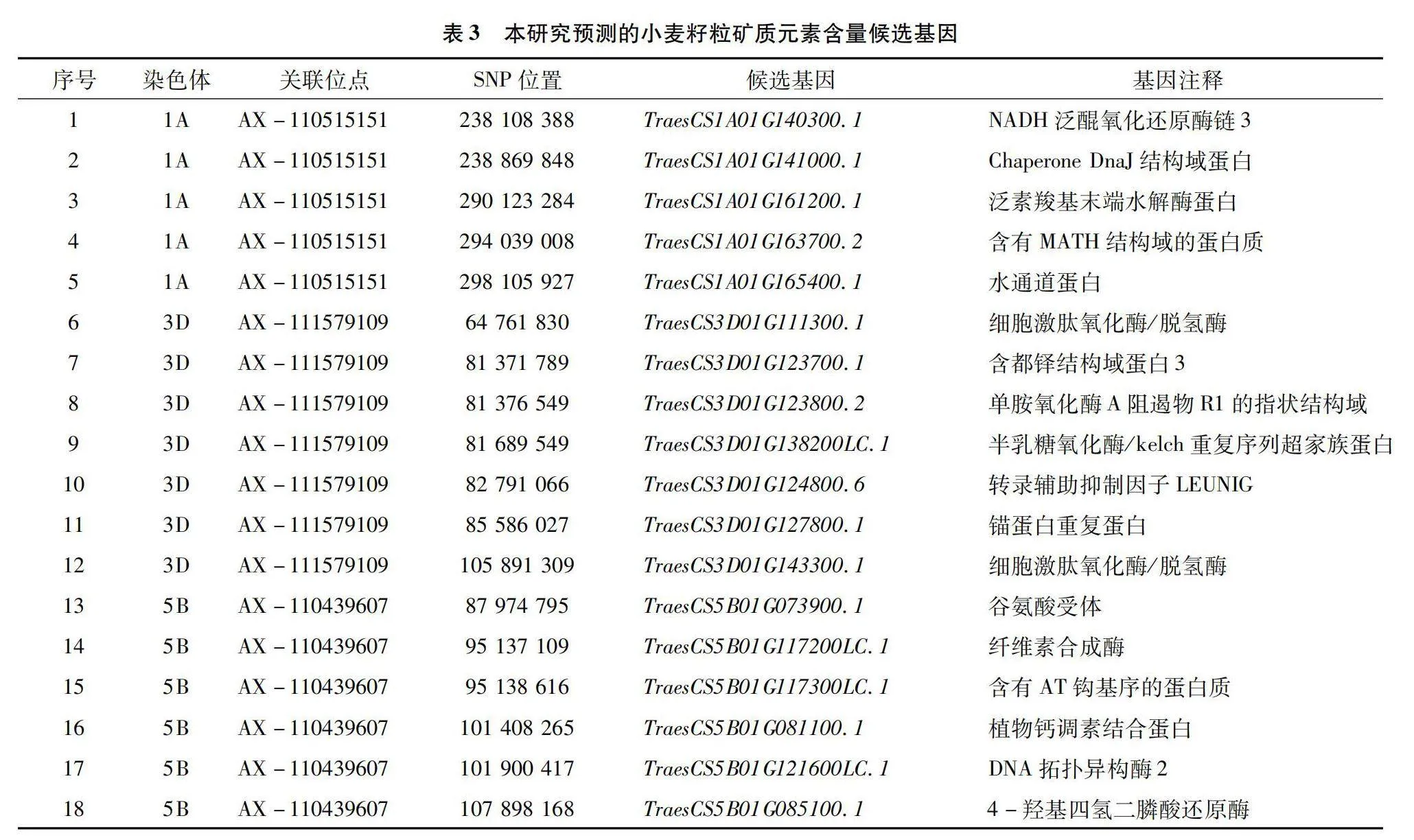
基于上述QTL定位結果,本研究對籽粒礦質元素進行了候選基因分析,具體結果見表3。由表3可知,共獲得籽粒礦質元素相關的候選基因18個,這些基因主要位于1A、3D和5B染色體上,其中在1A染色體上與AX-110515151關聯的候選基因有5個,分別是TraesCS1A01G140300.1、TraesCS1A01G141000.1、TraesCS1A01G161200.1、TraesCS1A01G163700.2和TraesCS1A01G165400.1。與3D染色體上AX-111579109關聯的候選基因有7個,包括TraesCS3D01G111300.1、TraesCS3D01G123700.1、TraesCS3D01G123800.2、TraesCS301G138200LC.1、TraesCS3D01G124800.6、TraesCS3D01G127800.1和TraesCS3D01G143300.1。與5B染色體上AX-110439607關聯的候選基因有6個,包括TraesCS5B01G073900.1、TraesCS5B01G117200LC.1、TraesCS5B01G117300LC.1、TraesCS5B01G081100.1、TraesCS5B01G121600LC.1和TraesCS5B01G085100.1。從候選基因的注釋可以看出,這些候選基因主要與小麥礦質元素的吸收、轉運、累積等功能相關。
3 討論與結論
隨著人們生活水平的提升和對自身健康的日益關注,提高籽粒微量礦質元素含量已成為主要糧食作物遺傳改良的重要目標之一,在其他作物上如玉米、水稻等已經開展了較多籽粒礦質元素研究[17-19],而有關小麥礦質元素的相關研究較少。在小麥籽粒礦質元素遺傳改良中,開展小麥新種質礦質元素測定與遺傳分析是培育高含量礦質元素小麥新品種的基礎性工作。普冰3228作為普通小麥與冰草遠緣雜交獲得的新種質,此前有關該種質礦質元素的研究尚未見報道。本研究以普冰3228和京4839為親本構建了重組自交系群體并對其Ca、Mg、Fe、Zn這4種礦質元素含量進行了QTL定位分析,結果檢測到與Zn相關的QTL 4個,其分別位于1A、5B和6B染色體上。張鵬霞在6B染色體上也發現了控制籽粒Zn含量的QTL[20];郝志在5B染色體上也檢測到控制籽粒Zn含量的QTL[3];孫憲印將小麥Zn含量QTL定位于3D、4A和6D染色體上[21];而Xu等在4B和5A染色體上檢測到了Zn含量QTL[22]。本研究在1A染色體上發現的Zn含量QTL在以往的研究中未見報道,可能為新發現的QTL。本研究將籽粒Ca元素含量QTL定位于1A、1D、5A和7D染色體上。張鵬霞也在7D染色體上發現了控制小麥籽粒Ca含量的QTL[20]。Shi等將小麥籽粒Ca含量QTL定位于2A、3A、4A、4B和5B等染色體上[9]。前人開展小麥籽粒Mg元素含量QTL分析研究較少,本研究和張鵬霞的研究[20]均在1A染色體上發現Mg含量QTL,在5A染色體上,本研究和孫憲印的研究[21]也均檢測到了Mg含量QTL。
性狀候選基因分析是解析其分子遺傳機制和進行分子遺傳改良的重要工作,雖然前人針對小麥礦質元素含量進行了部分QTL分析,但開展候選基因分析的研究報道甚少。本研究在對小麥籽粒礦質元素QTL定位的基礎上,對其也開展了候選基因分析,結果共獲得與籽粒元素相關的候選基因18個,這些候選基因主要編碼與元素吸收、轉運、累積等功能相關的蛋白或功能酶等。例如,TraesCS1A01G165400.1可能編碼一種水通道蛋白,已有研究證實該蛋白可影響植物體對部分礦質元素的吸收[23-24]。TraesCS5B01G081100.1可能編碼一種植物鈣調素結合蛋白,該蛋白可以影響植物對Ca元素的吸收[25-27]。TraesCS3D01G124800.6可能編碼一種轉錄輔助抑制因子,其可能通過影響與元素吸收轉運等代謝過程相關基因的轉錄間接影響植物對某些元素的吸收[28]。當然,本研究只是對小麥籽粒礦質元素含量相關候選基因的初步分析,這些基因還有待于下一步進行更為深入的研究。
參考文獻:
[1]譚綠貴,傅先蘭,張 鑫,等. 微量元素·人體健康·環境變化[J]. 微量元素與健康研究,2005,22(4):49-51.
[2]Anglani C. Wheat minerals:a review[J]. Plant Foods for Human Nutrition,1998,52:177-186.
[3]郝 志. 小麥籽粒主要有益礦質元素的含量及其QTL定位[D]. 泰安:山東農業大學,2008.
[4]Wang P,Wang H N,Liu Q,et al. QTL mapping of selenium content using a RIL population in wheat[J]. PLoS One,2017,12(9):e0184351.
[5]Wang Y,Xu X T,Hao Y F,et al. QTL mapping for grain zinc and iron concentrations in bread wheat[J]. Frontiers in Nutrition,2021,8:680391.
[6]AlomariD Z,Eggert K,von Wirén N,et al. Whole-genome association mapping and genomic prediction for iron concentration in wheat grains[J]. International Journal of Molecular Sciences,2018,20(1):76.
[7]AroraS,Cheema J,Poland J,et al. Genome-wide association mapping of grain micronutrients concentration in Aegilops tauschii[J]. Frontiers in Plant Science,2019,10:54.
[8]GorafiY S A,Ishii T,Kim J S,et al. Genetic variation and association mapping of grain iron and zinc contents in synthetic hexaploid wheat germplasm[J]. Plant Genetic Resources:Characterization and Utilization,2018,16(1):9-17.
[9]Shi X,Zhou Z F,Li W X,et al. Genome-wide association study reveals the genetic architecture for calcium accumulation in grains of hexaploid wheat (Triticum aestivum L.)[J]. BMC Plant Biology,2022,22(1):229.
[10]Wang J S,Shi X,Zhou Z F,et al. Genetic dissection of grain iron concentration in hexaploid wheat (Triticum aestivum L.) using a genome-wide association analysis method[J]. PeerJ,2022,10:e13625.
[11]王健勝,王 輝,劉偉華,等. 小麥-冰草多粒新種質及其多粒性遺傳分析[J]. 中國農業科學,2009,42(6):1889-1895.
[12]王健勝,劉偉華,王 輝,等. 小麥-冰草衍生系3228主要產量性狀的遺傳分析[J]. 植物遺傳資源學報,2010,11(2):147-151,156.
[13]Wang J S,Liu W H,Wang H,et al. QTL mapping of yield-related traits in the wheat germplasm 3228[J]. Euphytica,2011,177(2):277-292.
[14]Dellaporta S L,Wood J,Hicks J B. A plant DNA minipreparation:version Ⅱ[J]. Plant Molecular Biology Reporter,1983,1(4):19-21.
[15]Wang J K. Inclusive composite interval mapping of quantitative trait genes[J]. Acta Agronomica Sinica,2009,35(2):239-245.
[16]Meng L,Li H H,Zhang L Y,et al. QTL IciMapping:integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations[J]. The Crop Journal,2015,3(3):269-283.
[17]Hindu V,Palacios-Rojas N,Babu R,et al. Identification and validation of genomic regions influencing kernel zinc and iron in maize[J]. Theoretical and Applied Genetics,2018,131(7):1443-1457.
[18]Zhang J,Chen K,Pang YKOZyJch5WTAt+Ajtb8dxvw== L,et al. QTL mapping and candidate gene analysis of ferrous iron and zinc toxicity tolerance at seedling stage in rice by genome-wide association study[J]. BMC Genomics,2017,18(1):828.
[19]楊洪強,顧晶晶,田文仲,等. 不同灌水處理對小麥產量和籽粒品質的影響[J]. 江蘇農業科學,2022,50(19):74-78.
[20]張鵬霞. 不同氮鉀處理下小麥籽粒礦質元素相關性狀的QTL分析[D]. 泰安:山東農業大學,2021.
[21]孫憲印. 不同環境下小麥籽粒形態性狀及微營養物質含量的QTL分析[D]. 泰安:山東農業大學,2009.
[22]Xu Y F,An D G,Liu D C,et al. Molecular mapping of QTLs for grain zinc,iron and protein concentration of wheat across two environments[J]. Field Crops Research,2012,138:57-62.
[23]Li L Y,Pan S R,Melzer R,et al. Apoplastic barriers,aquaporin gene expression and root and cell hydraulic conductivity in phosphate-limited sheepgrass plants[J]. Physiologia Plantarum,2020,168(1):118-132.
[24]Maurel C,Verdoucq L,Luu D T,et al. Plant aquaporins:membrane channels with multiple integrated functions[J]. Annual Review of Plant Biology,2008,59:595-624.
[25]Wang W,Takezawa D,Narasimhulu S B,et al. A novel kinesin-like protein with a calmodulin-binding domain[J]. Plant Molecular Biology,1996,31(1):87-100.
[26]Xing J Y,Gruda N,Xiong J,et al. Influence of organic substrates on nutrient accumulation and proteome changes in tomato-roots[J]. Scientia Horticulturae,2019,252:192-200.
[27]PreussM L,Delmer D P,Liu B. The cotton kinesin-like calmodulin-binding protein associates with cortical microtubules in cotton fibers[J]. Plant Physiology,2003,132(1):154-160.
[28]Martin-Arevalillo R,Nanao M H,Larrieu A,et al. Structure of the Arabidopsis TOPLESS corepressor provides insight into the evolution of transcriptional repression[J]. Proceedings of the National Academy of Sciences of the United States of America,2017,114(30):8107-8112.
