optrA基因介導(dǎo)腸球菌對利奈唑胺耐藥的研究進(jìn)展
2024-12-17 00:00:00楊佩霓李慶蓉楊旭王清和平安呂梅張雙麗
中國抗生素雜志 2024年11期

摘要:腸球菌是導(dǎo)致尿路感染、心內(nèi)膜炎及醫(yī)院相關(guān)感染的常見病原體之一,利奈唑胺被認(rèn)為是治療由多重耐藥革蘭陽性菌引起嚴(yán)重感染的最后一種藥物,然而自利奈唑胺被引入臨床應(yīng)用以來,不斷有研究報道利奈唑胺耐藥腸球菌(linezolidin-resistant Enterococcus,LRE)的出現(xiàn),已成為臨床治療和公共衛(wèi)生面臨的重大挑戰(zhàn)。獲得可轉(zhuǎn)移性惡唑烷酮耐藥基因optrA是導(dǎo)致LRE的重要耐藥機(jī)制,其在腸球菌中的擴(kuò)散和流行,削弱了利奈唑胺治療臨床耐藥菌株的療效,應(yīng)在臨床應(yīng)用中引起極大重視。本文綜述了攜帶optrA基因的LRE菌株流行情況、參與optrA基因傳播的可移動性遺傳元件、optrA基因變體等,為臨床控制LRE的流行、合理選擇抗菌藥物等方面提供理論科學(xué)依據(jù)。
關(guān)鍵詞:腸球菌;利奈唑胺耐藥;利奈唑胺耐藥腸球菌;optrA基因
中圖分類號:R978.1 文獻(xiàn)標(biāo)志碼:A
Research progress on the optrA gene mediating linezolid resistance in enterococci
Abstract" " Enterococcus is one of the common pathogens causing urinary tract infections, endocarditis, and hospital-associated infections. Linezolid is considered to be the last line of treatment for serious infections caused by multidrug-resistant Gram-positive bacteria. However, since the introduction of linezolid into clinical application, the emergence of linezolidin-resistant Enterococcus (LRE) has been reported continuously, which has become a major challenge to clinical treatment and public health. The acquisition of the transferrable oxazolidinone resistance gene optrA is an important resistance mechanism leading to LRE. The spread and prevalence of optrA in enterococci weakened the efficacy of linezolid in the treatment of clinically resistant strains, and great attention should be paid to its clinical application. This article reviewed the prevalence of LRE strains carrying the optrA gene, the mobile genetic elements involved in the transmission of the optrA gene, and the variants of the optrA gene, so as to provide a theoretical and scientific basis for clinical control of the prevalence of LRE and rational selection of antibiotics.
Key words Enterococcus; Linezolid resistance; Linezolidin-resistant Enterococcus; optrA gene
腸球菌是一類革蘭陽性兼性厭氧菌,其廣泛存在于自然環(huán)境、人和動物的胃腸道中,在過去幾十年里,腸球菌被認(rèn)為是導(dǎo)致醫(yī)院相關(guān)感染如血液感染、心內(nèi)膜炎、腦膜炎和尿路感染的主要病原體之一。根據(jù)中國細(xì)菌耐藥監(jiān)測網(wǎng)(CHINET)統(tǒng)計在2021年1月—12月腸球菌屬細(xì)菌中糞腸球菌和屎腸球菌分別占41.8%和48.6%,其他腸球菌占9.6%[1]。腸球菌被認(rèn)為是醫(yī)院感染控制的重大挑戰(zhàn),不僅對多種抗菌藥物具有內(nèi)在耐藥性,還能通過質(zhì)粒和轉(zhuǎn)座子的轉(zhuǎn)移獲得額外的耐藥性,使它們成為多重耐藥菌株甚至形成高危腸球菌復(fù)合體(high-risk enterococcal complexes,HiRECC)在醫(yī)院環(huán)境中進(jìn)一步傳播。
惡唑烷酮類抗菌藥物是一類新型人工合成的抗生素,代表藥物為利奈唑胺,其在2007年引入我國的。利奈唑胺具有作用機(jī)制獨(dú)特、抗菌譜廣等特點(diǎn),其作用于細(xì)菌核糖體50S亞基的23S rRNA上,與肽基轉(zhuǎn)移酶中心(peptidyl transferase center,PTC)的A位點(diǎn)結(jié)合,阻止形成tRNAfMet、核糖體和mRNA三元起始復(fù)合物,同時阻止了mRNA的翻譯,從而抑制細(xì)菌蛋白質(zhì)的合成起到殺菌作用[2]。利奈唑胺作為1種全合成的抗菌藥物,細(xì)菌很難對其產(chǎn)生耐藥性。目前大多數(shù)革蘭陽性細(xì)菌(gt;99%)仍然對利奈唑胺敏感,然而由于惡唑烷酮類藥物在人類醫(yī)學(xué)中的頻繁使用,腸球菌可通過多種機(jī)制產(chǎn)生對利奈唑胺的耐藥性或敏感性降低,導(dǎo)致耐利奈唑胺腸球菌(linezolidin-resistant Enterococcus, LRE)的不斷出現(xiàn)和散播,因此利奈唑胺的耐藥性也受到全球范圍內(nèi)Zyvox年度藥效和譜評估(annual appraisal of potency and spectrum,ZAAPS)、利奈唑胺經(jīng)驗(yàn)及耐藥性準(zhǔn)確測定(linezolid experience and accurate determination of resistance,LEADER)等項(xiàng)目的監(jiān)測[3-4]。
近年來在世界范圍內(nèi)腸球菌對利奈唑胺耐藥性的發(fā)生率顯著增加,optrA基因作為腸球菌對利奈唑胺耐藥的重要機(jī)制之一,其在腸球菌中的傳播流行顯著降低了利奈唑胺治療腸球菌的療效,對人類健康構(gòu)成巨大的潛在威脅[5-6]。本文結(jié)合國內(nèi)外研究現(xiàn)狀對攜帶optrA基因的LRE菌株流行情況、腸球菌對利奈唑胺的耐藥機(jī)制包括參與optrA基因傳播的可移動性遺傳元件和optrA基因變體等方面進(jìn)行如下綜述。
1 optrA基因
optrA(oxazolidinonephenicol transferable resistance)基因最初是在來自中國的動物和人類糞腸球菌和屎腸球菌分離株中被發(fā)現(xiàn)的[7],其屬于ABC-F家族的ATP結(jié)合盒蛋白,與其他使用主動外排的ABC轉(zhuǎn)運(yùn)蛋白不同,其通過核糖體保護(hù)機(jī)制賦予抗菌藥物耐藥性[8]。optrA基因介導(dǎo)對惡唑烷酮類(利奈唑胺和泰地唑胺)和苯酚類(氯霉素和氟苯尼考)抗菌藥物交叉耐藥[9-10],一般情況下,optrA基因賦予利奈唑胺相對較低的MIC(4~16 mg/L)[11-12]。氟苯尼考作為一類獸用抗菌藥物,廣泛用于治療食源性動物呼吸道和腸道相關(guān)感染,在食源性動物中使用該藥物可能會共同選擇惡唑烷酮類藥物的耐藥性,這是對人類臨床和獸醫(yī)抗菌治療的嚴(yán)峻挑戰(zhàn)[13]。optrA基因的宿主菌主要為腸球菌,尤其是在糞腸球菌和屎腸球菌廣泛存在,此外在葡萄球菌、莢膜梭菌和彎曲桿菌等基因組中也有檢出[14-16]。截至目前,攜帶optrA基因的腸球菌菌株已在多個國家的多種來源中被發(fā)現(xiàn),涵蓋6大洲的29個國家或地區(qū),不僅包括住院患者、健康人群和各種食源性動物如雞、豬、牛和鴨等[17-19],還在城市廢水、蔬菜和土壤等被檢測到[20-21]。
2 攜帶optrA基因的LRE菌株流行情況
近年來,在世界范圍內(nèi)腸球菌對利奈唑胺耐藥性的發(fā)生率顯著增加[22-23],而optrA基因已經(jīng)逐漸成為腸球菌屬對利奈唑胺耐藥的主要機(jī)制,其在腸球菌中檢出率有逐漸上升趨勢。韓國一家三級護(hù)理醫(yī)院對利奈唑胺耐藥的糞腸球菌進(jìn)行耐藥機(jī)制調(diào)查,結(jié)果顯示所有利奈唑胺耐藥糞腸球菌菌株均攜帶有optrA基因[24]。法國一項(xiàng)監(jiān)測調(diào)查研究顯示,在2006—2016年LRE檢出率顯著增加,在2016年達(dá)到了0.8%,且主要是由于optrA基因的傳播導(dǎo)致的。美國SENTRY耐藥監(jiān)測項(xiàng)目統(tǒng)計報告顯示,2014—2016年,糞腸球菌對利奈唑胺耐藥最主要的機(jī)制是optrA基因?qū)е碌模?2.2%,26/36)。中國重慶一項(xiàng)耐藥監(jiān)測結(jié)果顯示,利奈唑胺耐藥腸球菌的分離率呈逐年上升,2011—2018年從0上升至6.8%,且98.8%LRE菌株攜帶optrA基因[25]。此外,在一些國家也報道了LRE菌株克隆性傳播暴發(fā)的情況,例如在上奧地利州出現(xiàn)由利奈唑胺耐藥屎腸球菌引起的醫(yī)院感染暴發(fā)[26];在韓國一家醫(yī)院出現(xiàn)耐利奈唑胺及耐萬古霉素腸球菌的暴發(fā)流行[27]。
耐藥基因擴(kuò)散傳播的重要機(jī)制之一是優(yōu)勢克隆菌株,optrA基因的傳播與克隆株也存在一定關(guān)系。攜帶optrA基因的LRE菌株的各種ST(sequence type)型如ST16、ST78、ST80、ST168、ST480、ST330和ST585等已在世界各地不同來源的樣本中被鑒定[28],在不同國家或地區(qū)可表現(xiàn)出不同的流行趨勢。中國南京一項(xiàng)對利奈唑胺耐藥的腸球菌分析中,ST16、ST480和ST476最常見,且出現(xiàn)了ST16克隆傳播的情況[29]。在西班牙一項(xiàng)研究中的LRE分離株屬于7種不同的STs,但ST16、ST480和ST585占主要優(yōu)勢[11]。在捷克共和國的一項(xiàng)臨床來源的利奈唑胺耐藥腸球菌的研究中,有超過50%的LRE菌株為ST80[5];在愛爾蘭的住院患者中ST480型的LRE菌株較為常見[28]。
3 腸球菌對利奈唑胺的耐藥機(jī)制
目前為止,腸球菌產(chǎn)生對利奈唑胺的耐藥性或敏感性降低的幾種機(jī)制包括:①23S rRNA結(jié)合位點(diǎn)的突變(如G2576U、G2447U和G2504A等等)和編碼50S核糖體蛋白的基因突變(如L3、L4、L22的rplC、rplD、rplV)。②獲得可轉(zhuǎn)移性惡唑烷酮耐藥基因,如編碼ABC-F家族的核糖體保護(hù)蛋白o(hù)ptrA基因和poxtA基因、修飾細(xì)菌23S rRNA甲基轉(zhuǎn)移酶的cfr基因家族[30-31]。cfr基因家族包含4種變體,即cfr(B)、cfr(C)、cfr(D)和cfr(E),其中cfr(C)和cfr(E)尚未在腸球菌中報道[32]。屎腸球菌對利奈唑胺的耐藥性通常與23S rRNA 基因的突變有關(guān),而獲得性利奈唑胺耐藥基因cfr、cfr(B)、optrA和poxtA主要發(fā)生在糞腸球菌中[30]。
4 參與optrA基因傳播的可移動性遺傳元件
細(xì)菌可通過水平基因轉(zhuǎn)移(horizontal gene transfer, HGT)獲得耐藥基因,進(jìn)而獲得相應(yīng)抗菌藥物抗性[33]。HGT通常由可移動遺傳元件(mobile genetic elements, MGE)如質(zhì)粒、轉(zhuǎn)座子、噬菌體、整合子和插入序列等介導(dǎo),其能促進(jìn)細(xì)菌耐藥基因在種內(nèi)甚至不同種屬間發(fā)生水平傳播。自首次發(fā)現(xiàn)人類和食源性動物的腸球菌中存在optrA基因以來,人們對其在腸球菌中傳播的遺傳基礎(chǔ)進(jìn)行了廣泛研究,之前的研究已表明optrA基因可以定位在菌株質(zhì)粒或染色體上,但主要位于可移動性的質(zhì)粒上,其在水平傳播過程中常常涉及不同的可移動遺傳元件(mobile genetic element, MGE)如質(zhì)粒、插入序列和轉(zhuǎn)座子[34]。
4.1 質(zhì)粒
質(zhì)粒是細(xì)菌染色體外的遺傳物質(zhì),目前在腸球菌中已知有6個家族的質(zhì)粒即theta復(fù)制質(zhì)粒(Rep_3, Inc18, RepA_N)和滾環(huán)復(fù)制質(zhì)粒(RCR: Rep_Trans, Rep_1和Rep_2),其中在腸球菌中與optrA基因相關(guān)的質(zhì)粒類型主要有與細(xì)菌素產(chǎn)生相關(guān)的非接合質(zhì)粒Rep_3、具有廣泛宿主范圍的Inc18、信息素反應(yīng)型質(zhì)粒RepA_N[35]。Inc18質(zhì)粒具有多種抗生素耐藥性,包括萬古霉素、氯霉素和大環(huán)內(nèi)酯-林可酰胺-鏈霉素(macrolides-lincoamide-streptomycin,MLS)組抗生素,該型質(zhì)粒常在腸球菌被檢測到[36]。信息素反應(yīng)型接合質(zhì)粒RepA_N為宿主范圍較窄的腸球菌質(zhì)粒,其主要分布于糞腸球菌中,在很大程度上與萬古霉素vanA耐藥基因的傳播有關(guān),但已有研究表明optrA基因介導(dǎo)的糞腸球菌利奈唑胺耐藥性可通過信息素反應(yīng)接合質(zhì)粒進(jìn)行傳播擴(kuò)散[37-38]。目前,許多報道表明攜帶optrA基因的可轉(zhuǎn)移性質(zhì)粒在體外可以通過接合或偶聯(lián)較為容易地在腸球菌菌株間轉(zhuǎn)移[18,38-40],被認(rèn)為是促進(jìn)腸球菌optrA基因進(jìn)行水平轉(zhuǎn)移或傳播的重要可移動性遺傳元件。
4.2 轉(zhuǎn)座子
位于腸球菌染色體上的optrA基因通常由轉(zhuǎn)座子(transposon, Tn)介導(dǎo)其在不同屬細(xì)菌之間傳播,其作用機(jī)制包括從宿主DNA中切除并產(chǎn)生環(huán)狀形式,然后將轉(zhuǎn)座子整合到新的靶序列中。目前與optrA基因水平傳播相關(guān)的轉(zhuǎn)座子包括Tn554家族轉(zhuǎn)座子Tn6674和Tn558。Tn554轉(zhuǎn)座子(GenBank登錄號:MK737778)攜帶轉(zhuǎn)座酶基因tnpA、tnpB、tnpC及大觀霉素耐藥基因spc和erm(A)[41]。Tn6674屬于非接合多抗性轉(zhuǎn)座子,最初在中國的豬源糞腸球菌分離株染色體DNA中被鑒定,其大小為12,932 bp,作為Tn554的衍生物,除攜帶Tn554轉(zhuǎn)座子本身具備的基因外,還攜帶耐藥基因optrA與氟苯尼考耐藥基因fexA。Tn6674被插入到染色體rac基因中,通過PCR實(shí)驗(yàn)可檢測到Tn6674的環(huán)狀形式,表明該轉(zhuǎn)座子具有功能活性,該類轉(zhuǎn)座子在希臘的人源糞腸球菌、韓國的雞肉以及瑞士的地表水中分離的糞腸球菌中被檢測出[40,42-43]。Tn558轉(zhuǎn)座子由轉(zhuǎn)座酶基因tnpA、tnpB、tnpC以及fexA構(gòu)成,該轉(zhuǎn)座子與腸球菌optrA基因環(huán)境的相關(guān)性在中國的人源以及豬源腸球菌均有報道[17,44]。Tn6261轉(zhuǎn)座子(GenBank登錄號:KU354267)來自中國的豬源糞腸球菌,該轉(zhuǎn)座子全長8,886 bp,除含有optrA基因外,還含有類似于Tn558的tnpA和tnpB基因、erm(A)基因和編碼依賴SAM的甲基轉(zhuǎn)移酶基因,該轉(zhuǎn)座子也被整合到糞腸球菌染色體rac基因中。
4.3 插入序列
插入序列(insert sequence, IS)也被稱為最簡單的轉(zhuǎn)座子,長度一般在3 kb以內(nèi),只編碼自身轉(zhuǎn)座所必需的轉(zhuǎn)座酶(transposase, tnp)。IS元件介導(dǎo)的重組在腸球菌攜帶的optrA基因傳播中起著重要作用。當(dāng)optrA基因編碼在質(zhì)粒時,通常位于IS1216E (IS6家族)或ISL3家族這類插入序列元件的上游或下游[35]。
IS1216E是IS1216的1種亞型,屬于IS6家族,其廣泛存在于多重耐藥的革蘭陽性菌中,是位于optrA基因側(cè)翼最常見的插入序列。optrA基因通常被兩個方向相同或相反、大小一致的IS1216E拷貝括起來,同時還攜帶兩個或更多基因包括耐藥基因erm(A)-like、cat和fexA;轉(zhuǎn)座酶基因tnp;DNA聚合酶基因impB;轉(zhuǎn)錄調(diào)節(jié)基因araC;酪氨酸激酶基因fer以及假定蛋白的hp基因等。在之前的研究已報道了optrA基因、IS1216E及其他基因共同組成的各種較為復(fù)雜的排列結(jié)構(gòu),如來自中國豬源糞腸球菌pE508分離的IS1216E-optrA-erm(A)-like-IS1216E序列(GenBank登錄號:EMK425645)、中國人源糞腸球菌pXM2013_42321分離的IS1216E-fexA-hp-optrA-IS1216E序列(GenBank登錄號:MH225423)、中國豬肉或蔬菜的糞腸球菌質(zhì)粒檢測到的IS1216E-hp-fexA-hp-optrA-hp-hp-hp-erm(A)-IS1216E[45]等。當(dāng)這些位于optrA基因側(cè)翼的IS1216E元件重組時,能產(chǎn)生一個可整合到接合質(zhì)粒、整合結(jié)合元件(integrative and conjuative element, ICEs)或不同染色體位點(diǎn)上的微型圓環(huán)(也被稱為易轉(zhuǎn)位單位TU),從而導(dǎo)致optrA基因在相同或不同菌種之間傳播。除了IS1216E外,插入序列ISEfa15被發(fā)現(xiàn)其存在于來自意大利臨床來源的攜帶有optrA基因的屎腸球菌分離株中[46],組成了ISEfa15-optrA-△ISEfa15序列片段,該片段在其他文獻(xiàn)中也被稱為Tn6628[47]。參與optrA基因傳播的可移動性遺傳元件如表1所示。
5 optrA基因變體
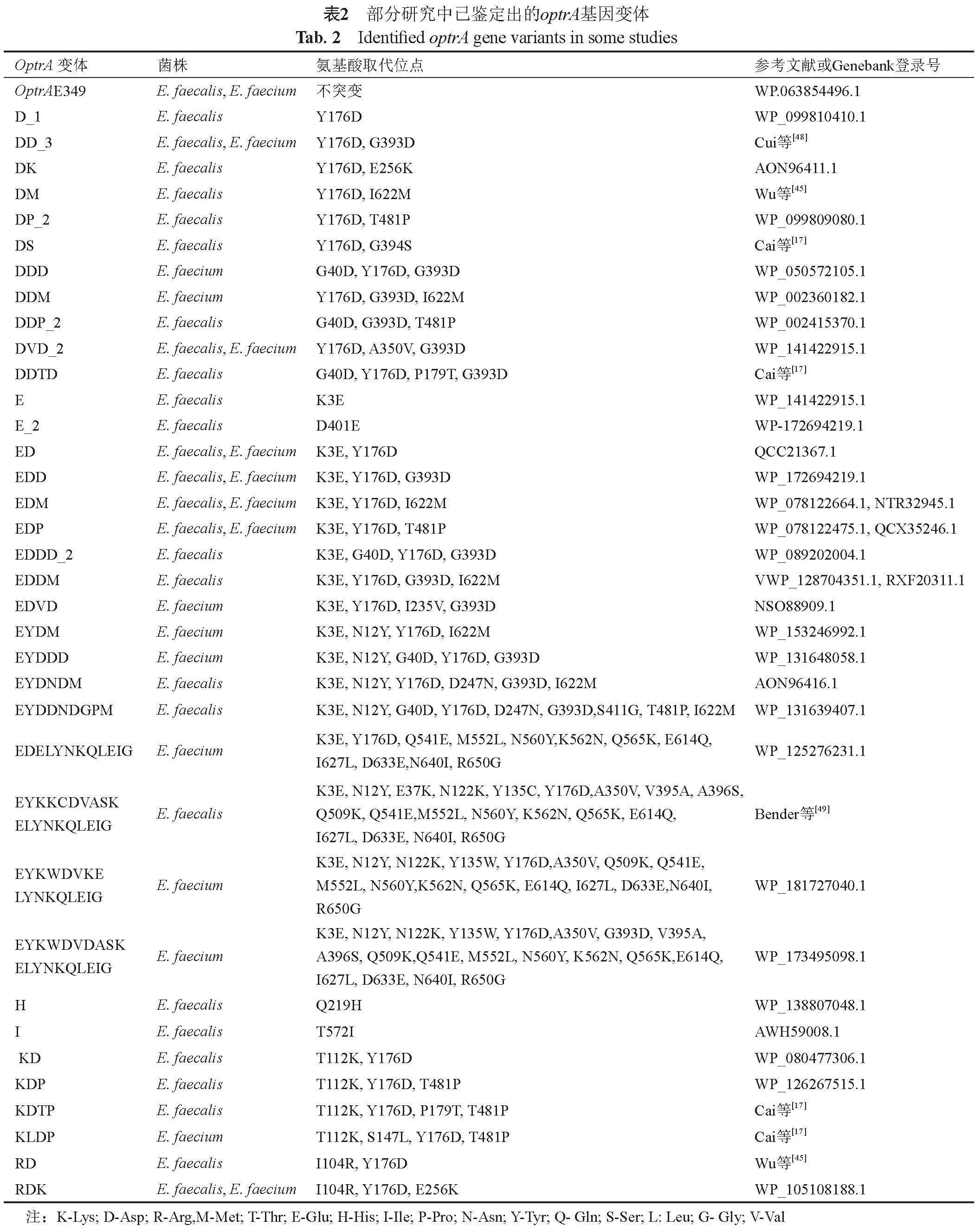
以optrA基因首次在腸球菌E349中被鑒定出的基因序列為野生型(wild-type, WT)optrA,之后不久便出現(xiàn)了與WT optrA基因不同的基因變體,目前至少鑒定出69種optrA基因變體,這些optrA基因變體表現(xiàn)為氨基酸的取代,與WT optrA基因序列相差1~20個氨基酸,其中較為常見的optrA變體就包括在176(Y176D)、393(G393D)、3(K3E)和40(G40D)位置的氨基酸替換。表2總結(jié)了部分研究中已鑒定出的optrA基因變體。
一些optrA基因變體在不同程度上可影響腸球菌對利奈唑胺的耐藥性,在之前的一些研究中測定了攜帶不同optrA變體菌株對利奈唑胺的MIC值。例如在Schwarz等[34]的研究中發(fā)現(xiàn),WT optrA基因及相關(guān)變體(D、EDP、KD、KLDP、RD、RDK和RDKP)常見于利奈唑胺耐藥分離株(MIC≥8 μg/mL),而其他變體(DDTD、EYDM、EYDDK、EYDNDM和KDTP)常見于利奈唑胺敏感分離株(MIC≤2 μg/mL)。此外在Li等[50]的研究中,成功構(gòu)建optrA基因過表達(dá)糞腸球菌,結(jié)果表明KD(T112K,Y176D)和RDK(I104R,Y176D,E256K)optrA變體能夠提高利奈唑胺MIC值。雖然部分optrA基因變體可影響腸球菌對利奈唑胺的耐藥性,但optrA基因變體與利奈唑胺相互作用的詳細(xì)機(jī)制還需要深入的研究闡明。
6 optrA基因與耐藥基因的共定位
6.1 與惡唑烷酮耐藥基因共定位
optrA基因可與poxtA、cfr等惡唑烷酮耐藥基因共存或3個基因同時共存于不同來源的腸球菌上[11,36,51-53],最常見的共定位包括optrA和cfr或optrA和cfr(D)。研究發(fā)現(xiàn)當(dāng)兩個或多個惡唑烷酮耐藥基因的同時存在可能是導(dǎo)致菌株對惡唑烷酮的最低抑菌濃度(MIC)較高的原因[54]。
6.2 與其他耐藥基因共定位
位于腸球菌的optrA基因也被發(fā)現(xiàn)與其他耐藥基因共存如fexA、erm(A)、erm(B)、tet(L)和tet(M)等,其中fexA-optrA或fexA-optrA-erm(A)共定位結(jié)構(gòu)較為常見。fexA和erm(A)基因分別介導(dǎo)菌株對酰胺醇類抗菌藥物(氯霉素、氟苯尼考)、大環(huán)內(nèi)酯類抗菌藥物耐藥。許多研究發(fā)現(xiàn)位于質(zhì)粒上optrA基因的上游均存在fexA基因[28,55-56],可能還同時存在ermA基因,出現(xiàn)fexA、optrA和ermA共同定位的結(jié)果[44],研究發(fā)現(xiàn)這種共定位現(xiàn)象表明耐利奈唑胺菌株的出現(xiàn)可能是由于使用非惡唑烷酮類抗菌藥物如大環(huán)內(nèi)酯類、氟苯尼考等選擇壓力下導(dǎo)致的[57]。根據(jù)之前的研究,在肉食動物類食品、醫(yī)院環(huán)境、城市廢水中均可分離到optrA陽性腸球菌[20,29,58],且位于質(zhì)粒上的optrA基因與人源性E349菌株野生型optrA序列具有高度相似性,表明了該基因在人類、動物、和環(huán)境之間具有高效傳播能力。由于食源性動物可導(dǎo)致含optrA基因的腸球菌傳播至人類,因此在動物生產(chǎn)和獸藥使用過程中應(yīng)謹(jǐn)慎使用酚類、酰胺醇類和大環(huán)內(nèi)酯類等抗菌藥物,并持續(xù)監(jiān)測畜禽養(yǎng)殖業(yè)動物腸球菌中的optrA基因的流行和傳播。
7 展望
利奈唑胺耐藥腸球菌的出現(xiàn)及optrA基因在腸球菌的廣泛傳播,為臨床抗感染治療帶來了極大的挑戰(zhàn),對公眾健康構(gòu)成潛在威脅,因此深入了解腸球菌中optrA基因的傳播機(jī)制及遺傳環(huán)境對臨床抗菌藥物的使用和控制耐藥菌的傳播有著重要意義。在臨床應(yīng)用抗菌藥物時應(yīng)根據(jù)實(shí)際情況合理用藥,謹(jǐn)慎使用利奈唑胺并采取相應(yīng)的感染控制措施以應(yīng)對危險因素,如長期住院、既往接觸抗生素和預(yù)防利奈唑胺耐藥性時的潛在疾病,減少利奈唑胺耐藥腸球菌的出現(xiàn)和進(jìn)一步傳播。此外,也應(yīng)意識到細(xì)菌對抗菌藥物產(chǎn)生耐藥的原因并不是孤立存在的,不僅需要對感染的患者、醫(yī)院、環(huán)境等進(jìn)行檢測和管控,同時也要對畜牧業(yè)、農(nóng)業(yè)等加強(qiáng)防治,才能阻斷耐藥菌的播散。
參 考 文 獻(xiàn)
胡付品, 郭燕, 朱德妹, 等. 2021年CHINET中國細(xì)菌耐藥監(jiān)測[J]. 中國感染與化療雜志, 2022, 22(5): 521-530.
胡宇華, 趙欣彤, 李天磊, 等. 惡唑烷酮類抗菌藥物研究進(jìn)展[J]. 藥學(xué)學(xué)報, 2022, 57(11): 3276-3291.
Mendes R E, Deshpande L, Streit J M, et al. ZAAPS programme results for 2016: An activity and spectrum analysis of linezolid using clinical isolates from medical centres in 42 countries[J]. J Antimicrob Chemother, 2018, 73(7): 1880-1887.
Pfaller M A, Mendes R E, Streit J M, et al. Five-year summary of in vitro activity and resistance mechanisms of linezolid against clinically important Gram-positive cocci in the United States from the LEADER surveillance program(2011 to 2015)[J]. Antimicrob Agents Chemother, 2017, 61(7): e00609-17.
Mali?ová L, Jakub? V, Pomorská K, et al. Spread of linezolid-resistant Enterococcus spp. in human clinical isolates in the Czech Republic[J]. Antibiotics(Basel), 2021, 10(2). doi: 10. 3390/antibiotics10020219.
Bender J K, Baufeld E, Becker K, et al. CHROMAgar? LIN-R as an efficient screening tool to assess the prevalence of linezolid-resistant enterococci in German hospital patients-a multicentre study approach, 2021-2022[J]. J Antimicrob Chemother, 2023, 78(9): 2185-2191.
Wang Y, Lv Y, Cai J, et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin[J]. J Antimicrob Chemother, 2015, 70(8): 2182-2190.
Wang Y, Li X, Wang Y, et al. Intracellular accumulation of linezolid and florfenicol in optrA-producing Enterococcus faecalis and Staphylococcus aureus[J]. Molecules, 2018, 23(12): 3195.
Sharkey L K, Edwards T A, O'Neill A J. ABC-F proteins mediate antibiotic resistance through ribosomal protection[J]. Mbio, 2016, 7(2): e01975.
Sharkey L K, O'Neill A J. Antibiotic resistance ABC-F proteins: Bringing target protection into the limelight[J]. Acs Infect Dis, 2018, 4(3): 239-246.
Ruiz-Ripa L, Fe?ler A T, Hanke D, et al. Mechanisms of linezolid resistance among Enterococci of clinical origin in Spain-detection of optrA- and cfr(D)-carrying E. faecalis[J]. Microorganisms, 2020, 8(8): 1155.
Zhang Y, Dong G, Li J, et al. A high incidence and coexistence of multiresistance genes cfr and optrA among linezolid-resistant enterococci isolated from a teaching hospital in Wenzhou, China[J]. Eur J Clin Microbiol, 2018, 37(8): 1441-1448.
Elghaieb H, Freitas A R, Abbassi M S, et al. Dispersal of linezolid-resistant enterococci carrying poxtA or optrA in retail meat and food-producing animals from Tunisia[J]. J Antimicrob Chemother, 2019, 74(10): 2865-2869.
Moawad A A, Hotzel H, Awad O, et al. Evolution of antibiotic resistance of coagulase-negative staphylococci isolated from healthy turkeys in Egypt: First report of linezolid resistance[J]. Microorganisms, 2019, 7(10): 476.
Zhou Y, Li J, Schwarz S, et al. Mobile oxazolidinone/phenicol resistance gene optrA in chicken Clostridium perfringens[J]. J Antimicrob Chemother, 2020, 75(10): 3067-3069.
Liu D, Yang D, Liu X, et al. Detection of the enterococcal oxazolidinone/phenicol resistance gene optrA in Campylobacter coli[J]. Vet Microbiol, 2020, 246: 108731.
Cai J, Schwarz S, Chi D, et al. Faecal carriage of optrA-positive enterococci in asymptomatic healthy humans in Hangzhou, China[J]. Clin Microbiol Infect, 2018, 25(5): 630. e1-630. e6.
Fukuda A, Nakajima C, Suzuki Y, et al. Transferable linezolid resistance genes(optrA and poxtA) in enterococci derived from livestock compost at Japanese farms[J]. J Glob Antimicrob Res, 2024, 36: 336-344.
項(xiàng)秋梅, 呂子全, 沈應(yīng)博, 等. 健康人群腸道中optrA陽性腸球菌的流行率及風(fēng)險因素[J]. 中華疾病控制雜志, 2022, 26(11): 1309-1314.
Freitas A R, Elghaieb H, León-Sampedro R, et al. Detection of optrA in the African continent(Tunisia) within a mosaic Enterococcus faecalis plasmid from urban wastewaters[J]. J Antimicrob Chemother, 2017, 72(12): 3245-3251.
Zhao Q, Wang Y, Wang S, et al. Prevalence and abundance of florfenicol and linezolid resistance genes in soils adjacent to swine feedlots[J]. Sci Rep, 2016, 6: 32192.
Bi R, Qin T, Fan W, et al. The emerging problem of linezolid-resistant enterococci[J]. J Glob Antimicrob Res, 2017, 13: 11-19.
Càmara J, Camoez M, Tubau F, et al. Detection of the novel optrA gene among linezolid-resistant enterococci in Barcelona, Spain[J]. Microb Drug Resist, 2018, 25(1): 87-93.
Hu Y, Won D, Nguyen L P, et al. Prevalence and genetic analysis of resistance mechanisms of linezolid-nonsusceptible Enterococci in a tertiary care hospital Examined via whole-genome sequencing[J]. Antibiotics(Basel), 2022, 11(11): 1624.
鄒家齊. 利奈唑胺耐藥腸球菌的耐藥傳播機(jī)制研究及感染危險因素調(diào)查[D]. 重慶: 重慶醫(yī)科大學(xué), 2020.
Kerschner H, Cabal A, Hartl R, et al. Hospital outbreak caused by linezolid resistant Enterococcus faecium in upper Austria[J]. Antimicrob Resist Infect Control, 2019, 8: 150.
Jung J, Park K, Shin S H, et al. The pitfall of cohort isolation in an outbreak of linezolid-resistant, vancomycin-resistant enterococci[J]. Clin Microbiol Infec, 2019, 25(12): 1568-1569.
Egan S A, Shore A C, O'Connell B, et al. Linezolid resistance in Enterococcus faecium and Enterococcus faecalis from hospitalized patients in Ireland: High prevalence of the MDR genes optrA and poxtA in isolates with diverse genetic backgrounds[J]. J Antimicrob Chemother, 2020, 75(7): 1704-1711.
Park K, Jeong Y S, Chang J, et al. Emergence of optrA-mediated linezolid-nonsusceptible Enterococcus faecalis in a tertiary care hospital[J]. Ann Lab Med, 2020, 40(4): 321-325.
Sadowy E. Linezolid resistance genes and genetic elements enhancing their dissemination in enterococci and streptococci[J]. Plasmid, 2018, 99: 89-98.
Roy S, Aung M S, Paul S K, et al. Drug resistance determinants in clinical isolates of Enterococcus faecalis in Bangladesh: Identification of oxazolidinone resistance gene optrA in ST59 and ST902 lineages[J]. Microorganisms, 2020, 8(8): 1240.
Shan X, Li X S, Wang N, et al. Studies on the role of IS1216E in the formation and dissemination of poxtA-carrying plasmids in an Enterococcus faecium clade A1 isolate[J]. J Antimicrob Chemother, 2020, 75(11): 3126-3130.
Hegstad K, Mikalsen T, Coque T M, et al. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant Enterococcus faecalis and Enterococcus faecium[J]. Clin Microbiol Infect, 2010, 16(6): 541-554.
Schwarz S, Zhang W, Du X D, et al. Mobile oxazolidinone resistance genes in Gram-positive and Gram-negative bacteria[J]. Clin Microbiol Rev, 2021, 34(3): e0018820.
Freitas A R, Tedim A P, Novais C, et al. Comparative genomics of global optrA-carrying Enterococcus faecalis uncovers a common chromosomal hotspot for optrA acquisition within a diversity of core and accessory genomes[J]. Microb Genom, 2020, 6(6): e000350.
Kohler V, Vaishampayan A, Grohmann E. Broad-host-range Inc18 plasmids: Occurrence, spread and transfer mechanisms[J]. Plasmid, 2018, 99: 11-21.
Zou J, Tang Z, Yan J, et al. Dissemination of linezolid resistance through sex pheromone plasmid transfer in Enterococcus faecalis[J]. Front Microbiol, 2020, 11: 1185.
Shang Y, Li D, Shan X, et al. Analysis of two pheromone-responsive conjugative multiresistance plasmids carrying the novel mobile optrA locus from Enterococcus faecalis[J]. Infect Drug Resist, 2019, 12: 2355-2362.
Sassi M, Guérin F, Zouari A, et al. Emergence of optrA-mediated linezolid resistance in enterococci from France, 2006-16[J]. J Antimicrob Chemother, 2019, 74(6): 1469-1472.
Yoon S, Son S H, Kim Y B, et al. Molecular characteristics of optrA-carrying Enterococcus faecalis from chicken meat in South Korea[J]. Poult Sci, 2020, 99(12): 6990-6996.
Li D, Li X Y, Schwarz S, et al. Tn6674 is a novel Enterococcal optrA-carrying multiresistance transposon of the Tn554 Family[J]. Antimicrob Agents Chemother, 2019, 63(9): e00809-19.
Tsilipounidaki K, Gerontopoulos A, Papagiannitsis C, et al. First detection of an optrA-positive, linezolid-resistant ST16 Enterococcus faecalis from human in Greece[J]. New Microbes New Infect, 2019, 29: 100515.
Nüesch-Inderbinen M, Raschle S, Stevens, M J A, et al. Linezolid-resistant Enterococcus faecalis ST16 harbouring optrA on a Tn6674-like element isolated from surface water[J]. J Glob Antimicrob Res, 2021, 25: 89-92.
Kang Z Z, Lei C W, Kong L H, et al. Detection of transferable oxazolidinone resistance determinants in Enterococcus faecalis and Enterococcus faecium of swine origin in Sichuan Province, China[J]. J Glob Antimicrob Re, 2019, 19: 333-337.
Wu Y, Fan R, Wang Y, et al. Analysis of combined resistance to oxazolidinones and phenicols among bacteria from dogs fed with raw meat/vegetables and the respective food items[J]. Sci Rep, 2019, 9(1): 15500.
Morroni G, Brenciani A, Antonelli A, et al. Characterization of a multiresistance plasmid carrying the optrA and cfr resistance genes from an Enterococcus faecium clinical isolate[J]. Front Microbiol, 2018, 9: 2189.
D'Andrea M M, Antonelli A, Brenciani A, et al. Characterization of Tn6349, a novel mosaic transposon carrying poxtA, cfr and other resistance determinants, inserted in the chromosome of an ST5-MRSA-II strain of clinical origin[J]. J Antimicrob Chemother, 2019, 74(10): 2870-2875.
Cui L, Wang Y, Lv Y, et al. Nationwide surveillance of novel oxazolidinone resistance gene optrA in Enterococcus isolates in China from 2004 to 2014[J]. Antimicrob Agents Chemother, 2016, 60(12): 7490-7493.
Bender J K, Fleige C, Lange D, et al. Rapid emergence of highly variable and transferable oxazolidinone and phenicol resistance gene optrA in German Enterococcus spp. clinical isolates[J]. Int J Antimicrob Agents, 2018, 52(6): 819-827.
Li P, Yang Y, Ding L, et al. Molecular investigations of linezolid resistance in enterococci optrA variants from a hospital in Shanghai[J]. Infect Drug Resist, 2020, 13: 2711-2716.
Chen L, Han D, Tang Z, et al. Co-existence of the oxazolidinone resistance genes cfr and optrA on two transferable multi-resistance plasmids in one Enterococcus faecalis isolate from swine[J]. Int J Antimicrob Agents, 2020, 56(1): 105993.
Ruiz-Ripa L, Fe?ler A T, Hanke D, et al. Detection of poxtA- and optrA-carrying E. faecium isolates in air samples of a Spanish swine farm[J]. J Glob Antimicrob Res, 2019, 22: 28-31.
Hao W, Shan X, Li D, et al. Analysis of a poxtA- and optrA-co-carrying conjugative multiresistance plasmid from Enterococcus faecalis[J]. J Antimicrob Chemother, 2019, 74(7): 1771-1775.
Brenciani A, Morroni G, Schwarz S, et al. Oxazolidinones: Mechanisms of resistance and mobile genetic elements involved[J]. J Antimicrob Chemother, 2022, 77(10): 2596-2621.
Gawryszewska I, ?abicka D, Hryniewicz W, et al. Linezolid-resistant enterococci in Polish hospitals: Species, clonality and determinants of linezolid resistance[J]. Eur J Clin Microbiol, 2017, 36(7): 1279-1286.
Tyson G H, Sabo J L, Hoffmann M, et al. Novel linezolid resistance plasmids in Enterococcus from food animals in the USA[J]. J Antimicrob Chemother, 2018, 73(12): 3254-3258.
Wang Y, Li X, Fu Y, et al. Association of florfenicol residues with the abundance of oxazolidinone resistance genes in livestock manures[J]. J Hazard Mater, 2020, 399: 123059.
Jung Y H, Cha M H, Woo G J, et al. Characterization of oxazolidinone and phenicol resistance genes in non-clinical enterococcal isolates from Korea[J]. J Glob Antimicrob Res, 2021, 24: 363-369.
