岡田酸抑制神經元軸突生長
2011-06-14 06:26:40鄭紅云童永清
卒中與神經疾病 2011年6期
關鍵詞:海馬
鄭紅云 李 艷 童永清
神經元的極性是指絕大多數神經元存在一個長的軸突和幾個短的樹突。對于一個神經元來說,樹突往往是接受信息的部位,軸突往往是輸出信息的部位,因此神經元的極性是保證信息在神經系統內部及神經系統與其他系統之間有序流動的物質基礎。但是神經元極性形成的分子機制仍是一個有趣且具挑戰性的問題。
胎鼠海馬神經元培養是研究神經元極性的良好模型[1]。海馬神經元極性的建立經歷層狀偽足、無極性的突起以及軸突和樹突形成三個階段[2]。研究表明,微管對軸突和樹突的分化成熟起著關鍵作用[3];進一步研究揭示微管相關蛋白tau和微管相關 蛋 白 1B(microtubule-associated protein 1B,MAP1B)促進神經元軸突的形成[4];腦衰蛋白反應調節蛋白2(collapsin response mediator protein-2,CRMP-2)可結合管蛋白,促進微管聚集,進而調節軸突和樹突極性形成[5]。最近2項研究表明糖原激酶合酶3((glycogen synthase kinase 3,GSK-3()通過調節 CRMP-2磷酸化狀態影響神經元極性[6,7]。大量事實表明蛋白質磷酸化狀態與神經元極性形成關系密切,而蛋白質磷酸化狀態受蛋白磷酸激酶和磷酸酯酶的雙向調節。近年來研究報道了激酶對神經元極性的影響,而蛋白磷酸酯酶對神經元極性的影響尚無研究。
本實驗采用蛋白磷酸酯酶(PP2A)特異性抑制劑OA在海馬神經元極性形成的早期處理海馬神經元,探討PP2A在神經元極性形成中是否起作用,為神經元發生的機制提供新的理論基礎。
1 材料與方法
1.1 動物 孕18 d的SD大白鼠(由華中科技大學同濟醫學院實驗動物學部提供)。
1.2 試劑及抗體 OA購于Calbiochem公司(La Jalla,CA,USA),DMSO購于sigma公司。MAP2和Tau-1購于Chemicon公司。
1.3 原代海馬神經元細胞的培養 無菌條件下取出孕18 d胎鼠的完整腦組織,剔除腦膜和血管,鈍性分離出雙側海馬;將海馬剪成1 mm3的組織塊(以上均在冰上進行),置入解剖液(含3%~6%葡萄糖D-Hanks)中,0.125%胰蛋白酶(Gibco,USA)37℃消化15 min,加入種植培養液(含10%胎牛血清的DMEM培養基)終止反應,滴管吹打后經200目篩網濾過,1500 r/min離心5 min,加入適當的種植培養液重懸,細胞計數,調至1x107/L,接種于預鋪多聚賴氨酸的培養板或培養瓶中,于37℃、5%CO2培養箱中培養;12 h后換成維持培養基(含5%小牛血清、2%B27的DMEM培養基)。
1.4 OA處理海馬神經元 海馬神經元原代培養24 h后分別給予不同濃度的OA處理細胞24 h,普通光學顯微鏡下觀察海馬神經元形態變化。
1.5 免疫熒光雙重標記 海馬神經元原代培養24 h,即給予OA處理細胞24 h后吸出培養基,0.01 M磷酸鹽緩沖液(phosphate buffered saline,PBS)漂洗后加入100%甲醇-20 ℃固定5 min,PBS-0.2%Triton漂洗,然后 PBS-0.5%Triton破膜5~10 min,PBS-0.2%Triton漂洗,5%牛血清白蛋白(bovine serum albumin,BSA)封閉1 h,分別滴加鼠抗Tau-1(1∶200)和兔抗微管相關蛋白2(microtubule-associated protein-2,MAP-2)(1∶1000),于4℃孵育過夜,漂洗后加入熒光標記Ⅱ抗于室溫孵育1 h,漂洗后于共聚焦顯微鏡下觀察。
1.6 圖像分析和統計學處理 采用統計產品與服務解決方案軟件(statistical product and service solutions 11.0,SPSS 11.0),全部數據采用平均數±標準差±s)表示,采用Image-Proplus軟件統計突起長度。
2 結 果
2.1 正常海馬神經元極性形成的形態觀察
普通光學顯微鏡下可見種植4 h后少部分細胞貼壁,細胞表面呈現單個或多個層狀偽足;種植12 h后大部分細胞貼壁,多數神經元出現無分支的突起;種植24 h后細胞完全貼壁,可見雙突起、三個突起和多個突起的神經元,其中一個突起迅速生長成為軸突;種植48 h后其它的突起發展成為樹突,并且突起之間交錯成網(圖1)。
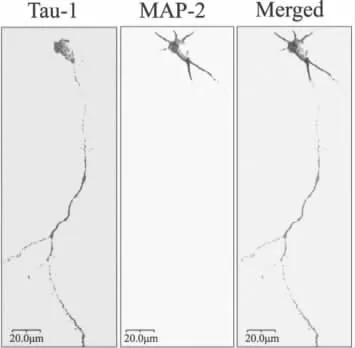
圖1 原代海馬神經元培養48 h后固定做免疫熒光,分別采用軸突特異性標記物Tau-1和樹突特異性標記物MAP2標記神經元(Bar=20μm)
2.2 OA抑制原代海馬神經元軸突生成
普通光學顯微鏡下可見DMSO對照組神經元的軸突和樹突已經形成,神經元極性完全建立;采用PP2A抑制劑OA處理24 h后各濃度組神經元突起的生長均受到了不同程度的抑制,且隨OA濃度的增加抑制作用逐漸增強,呈現劑量依賴性(圖2)。與對照組(平均長度為185μm)相比,不同濃度OA處理后神經元軸突平均長度分別為150μm、100 μm、80μm、75μm、50μm。
3 討 論
神經元極性形成對其功能的正常發揮至關重要,探索神經元極性形成的分子基礎尤為迫切。本實驗結果可見正常海馬神經元培養48 h后極性已完全形成,而培養24 h后用PP2A特異性抑制劑OA處理后普通光學顯微鏡下可見隨OA濃度的增加,對神經元突起生長的抑制作用逐漸增強,呈劑量依賴性,該結果提示PP2A參與了神經元極性的形成。
OA是一種廣泛使用的PP2A選擇性抑制劑。最新研究表明OA處理皮質神經元可導致tau蛋白和CRMP-2均發生不同程度的磷酸化,并首次提出CRMP-2磷酸化狀態受PP2A活性的調節[8]。微管相關蛋白tau、MAP1B以及CRMP-2可結合管蛋白,促進微管聚集,進而調節軸突和樹突極性形成[9,10]。在發育的新生海馬神經元軸突末端以及生長錐,tau蛋白、MAP1B以及CRMP-2主要以非磷酸化形式存在[11];GSK-3β磷酸化 CRMP-2降低其與管蛋白的結合能力,抑制微管的聚集和神經元軸突生長[12],而PP2A廣泛存在于微管相關蛋白區域[13],可以去磷酸化tau蛋白、MAP1B和 CRMP-2而促進微管聚集。因此,tau蛋白、MAP1B和CRMP-2可能作為PP2A下游靶分子以調節神經元極性。
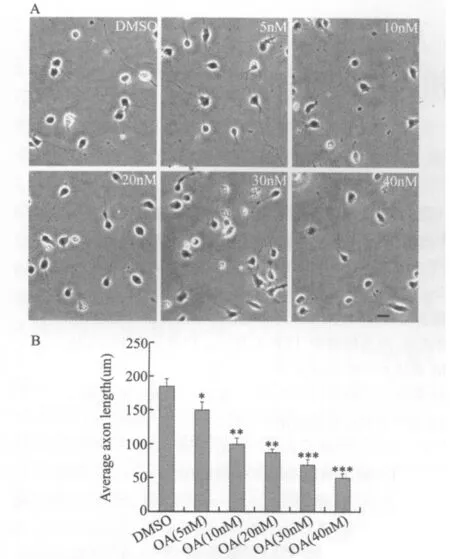
圖2 不同濃度OA處理后對神經元軸突形成的影響
在常見神經退行性疾病如Alzheimer病(AD)患者腦中PP2A活性明顯降低,tau蛋白以高度磷酸化的形式存在[14],進而出現神經元極性的喪失并最終導致神經元的大量丟失,因此上調PP2A對促進AD患者腦中神經元正常生存意義重大。因此,該研究為神經元極性形成的分子機制提供了新資料,同時為神經退行性疾病的治療提供了新的理論基礎。
1 Arimura N,Kaibuchi K.Neuronal polarity:from extracellular signals to intracellular mechanisms.Nat Rev Neurosci,2007,8(3):194-205.
2 Szu-Yu Ho,Rasband MN.Maintenance of neuronal polarity.Dev Neurobiol,2011,71(6):474-482.
3 Buck KB,Zheng JQ.Growth cone turning induced by direct local modification of microtubule dynamics.J Neurosci,2002,22(21):9358-9367.
4 Tahirovic S,Bradke F.Neuronal polarity.Cold Spring Harb Perspect Biol,2009,1(3):16-44.
5 Arimura N.Role of CRMP-2 in neuronal polarity.J Neurobiol,2004,58(1):34-47.
6 Jiang H,et al.Both the establishment and the maintenance of neuronal polarity require active mechanisms:critical roles of GSK-3beta and its upstream regulators.Cell,2005,120(1):123-135.
7 Yoshimura T.GSK-3beta regulates phosphorylation of CRMP-2 and neuronal polarity.Cell,2005,120(1):137-149.
8 Hill JJ.Identification of okadaic acid-induced phosphorylation events by a mass spectrometry approach.Biochem Biophys Res Commun,2006,342(3):791-799.
9 Hirai S.Axon formation in neocortical neurons depends on stagespecific regulation of microtubule stability by the dual leucine zipper kinase-c-Jun N-terminal kinase pathway.J Neurosci,2011,31(17):6468-6480.
10 Hoogenraad CC,Bradke F.Control of neuronal polarity and plasticity--a renaissance for microtubules?Trends Cell Biol,2009,19(12):669-676.
11 Yoshimura T,Arimura N,Kaibuchi K.Molecular mechanisms of axon specification and neuronal disorders.Ann N Y Acad Sci,2006,1086:116-125.
12 Ni MH.GSK-3 mediates the okadaic acid-induced modification of collapsin response mediator protein-2 in human SK-N-SH neuroblastoma cells.J Cell Biochem,2008,103(6):1833-1848.
13 Awotunde OS.Interaction of maize(Zea mays)protein phosphatase 2A with tubulin.Acta Biochim Pol,2003,50(1):131-138.
14 Small SA,Duff K.Linking Abeta and tau in late-onset Alzheimer's disease:a dual pathway hypothesis.Neuron,2008,60(4):534-542.
猜你喜歡
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
幼兒智力世界(2022年3期)2022-03-12 03:19:19
娃娃樂園·綜合智能(2020年8期)2020-08-28 00:32:14
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
作文周刊·小學二年級版(2018年9期)2018-04-18 10:01:40
小學生導刊(2018年1期)2018-03-15 08:02:37
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
科技知識動漫(2016年6期)2016-06-24 21:04:53
大灰狼(2015年6期)2015-07-16 21:01:00
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
