細粒棘球絳蟲Eg95抗原表位的生物信息學預測*
2011-08-21 10:23:36李玉嬌賈海英馬秀敏丁劍冰
中國人獸共患病學報 2011年10期
關鍵詞:區域
李玉嬌,王 晶,趙 慧,賈海英,李 博,馬秀敏,溫 浩,丁劍冰,
2.新疆醫科大學附屬中醫醫院,烏魯木齊 830000;
3.新疆自治區人民醫院,烏魯木齊 830000;
4.新疆醫科大學基礎醫學院,烏魯木齊 830011
細粒棘球絳蟲Eg95抗原表位的生物信息學預測*
李玉嬌1,王 晶2,趙 慧1,賈海英3,李 博2,馬秀敏4,溫 浩1,丁劍冰1,4
目的 應用軟件和在線網絡分析Eg95的蛋白二級結構,預測B細胞表位和T細胞表位,為確定和篩選優勢表位,研制安全、高效的表位疫苗奠定基礎。方法 采用DNAStar軟件和在線網站IEDB、SYFPEITHI等對Eg95的B細胞表位和 T細胞表位進行預測。結果 Eg95存在潛在的抗原表位,B細胞表位區域:16-38,41-48,50-67,68-90,92-100,103-113,120-132。分值高的 T細胞表位區域:6-14,14-22,48-56,4-12,98-106,142-150,146-154。結論 運用生物信息學方法確定Eg95存在7個抗原表位,對進一步研究Eg95的抗原性和研發優勢表位疫苗具有重要的意義。
Eg95;抗原表位;生物信息學
2.新疆醫科大學附屬中醫醫院,烏魯木齊 830000;
3.新疆自治區人民醫院,烏魯木齊 830000;
4.新疆醫科大學基礎醫學院,烏魯木齊 830011
細粒棘球蚴病(echinococcosis)又稱囊型包蟲病(cystic echinococcosis,CE),是由細粒棘球絳蟲(Echinococcusgranulosus,Eg)幼蟲引起的一種人獸共患慢性寄生蟲病,呈全球性分布。我國是細粒棘球蚴病發病最高的國家之一,至今已在25個省、市、自治區發現有人、畜原發性棘球蚴病,主要流行在西部的新疆、青海、甘肅、寧夏等牧區和半牧區[1]。經過流行病學統計,2004-2008年總體上報告包蟲病病例數呈增加趨勢[2]。目前對于包蟲病的治療主要以手術治療為主,藥物治療為輔,效果欠佳。采用給犬定期驅蟲和宣傳教育為主的綜合性防治措施的效果也不理想,因此人們一直在研究更有效的防治措施。免疫預防是防止包蟲病流行比較理想的途徑。采用現代分子生物學技術對細粒棘球絳蟲的有效免疫原成分進行篩選和克隆,制備基因工程疫苗,為包蟲病的免疫預防和免疫診斷開辟了新途徑[3]。Lightowlers等[4]發現Eg95重組蛋白疫苗免疫中間宿主(羊),可抵抗六鉤蚴的感染獲得95%~100%的免疫保護作用,其中86%得到完全保護,認為Eg95是較理想的保護性抗原。賈海英等[5]將Eg95與pET28a重組融合構建了pET28a-Eg95重組質粒,經過Western blot實驗,其中11例陽性患者的血清中有8份都與pET28a-Eg95重組蛋白有陽性反應,其中5份正常人的血清不與其發生陽性反應,該結果表明Eg95抗原在CE病人體內有較高免疫原性。隨著分子生物學技術的發展,表位疫苗[6](Epitope vaccine)成為分子疫苗的研究熱點,它是根據抗原表位氨基酸序列制備而成的疫苗,包括合成肽疫苗[7](synthetic peptide vaccine)、重組表位疫苗[8](Recombinant epitope-based vaccine)及表位 核 酸 疫 苗[9](Epitope DNA vaccine,minigenes/epigenes)等形式,是目前研制抗感染性疾病、惡性腫瘤、寄生蟲病疫苗的發展方向,研制表位疫苗面臨的最大挑戰就是如何找到抗原上免疫原性最強的區域,即表位所在的位置[10]。Eg95分子是一種分子量為24.5kDa的天然六鉤蚴抗原,本研究運用生物信息學方法進行Eg95的B細胞和T細胞抗原表位預測,為包蟲病診斷和表位疫苗的研制奠定基礎。
1 材料與方法
1.1 材料
1.1.1Eg95氨基酸序列 根據GenBank的Eg95基因序列,經過多重序列比對,確定最終進行分析的Eg95基因序列(GenBank登錄號:HM345607),長471bp,用DNAMAN軟件推導出Eg95抗原氨基酸序列,它編碼156個氨基酸殘基:MAFQLCLILFATSVLAQEYKGVGKGQGQQETPLRNHFNL TPVGSQGIRLSWEVQHLSDLKGTDISLRAVN PSDPLVCKRQTAKFSDGQLAVGELKPSTLYK MTVEAVKAKKTILGFTVDIETPRAGKKEST VMTSGSALTSAIAGFVFSCIVVVLT
1.1.2 蛋白質二級結構預測軟件TMHMM Sever2.0 (http://www.cbs.dtu.dk/services/ TMHMM)分析Eg95抗原的跨膜序列,SOPMA Sever(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)預測Eg95的蛋白二級結構。
1.1.3 抗原表位預測軟件 B細胞表位:DNAStar軟件及在線預測軟件IEDB (http://tools.immuneepitope.org/main/index.html)。T 細胞表位:(SYFPEITHI http://www.syfpeithi.de)和(BIMAS http://bimas.dcrt.nih.gov/ molbio/hla_bind/)。
1.2 方法
1.2.1 采用DNAStar軟件的Protean程序以及在線預測網站IEDB (http://tools.immuneepitope.org/main/index.html)結合親水性參數、可及性參數、抗原性指數、柔韌性參數及二級結構方案對Eg95抗原的B細胞表位進行綜合預測。
1.2.2 通過在線網絡資源的使用在以下兩個網站:(SYFPEITHI http://www.syfpeithi.de)和(BIMAS http://bimas.dcrt.nih.gov/molbio/hla_bind/)通過其各種算法對Eg95抗原的T細胞表位進行預測。
2 結 果
2.1 B細胞表位預測結果
2.1.1 Eg95跨膜結構預測 用 TMHMM 對Eg95的跨膜序列進行分析,Eg95的跨膜區在133-155位區域,其中1-132位區域顯示為膜外區,156為膜內區(見圖1)。
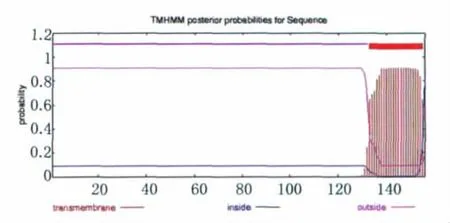
圖1 TMHMM分析的Eg95跨膜結構Fig.1 Transmembrane struture of Eg95using TMHMM analysis
2.1.2Eg95抗原蛋白質二級結構預測 利用SOPMA Sever對Eg95的二級結構進行預測,發現Eg95基因中柔性結構占氨基酸總數的43.59%,其中無規則卷曲占40.38%,β轉角為3.21%。α螺旋和β折疊分別占到32.05%和24.36%,各種結構在Eg95抗原中的分布情況見圖2。

圖2 SOPMA分析的Eg95二級結構Fig.2 Secondary structure of Eg95using SOPMA analysis
2.1.3 用DNAStar軟件對Eg95B細胞表位預測DNAStar軟件對Eg95的預測結合了以下有代表性的參數綜合預測方案(見圖3):1)以 Hopp和Woods方案預測的Eg95親水性區域為:15-40,47-65,69-87,96-101,120-134。2)用Emini蛋白可及性方案分析,分值比較高的區域為:24-37,79-86,94-111,121-131。3)經過 Karplus-Sohulz柔性區域的分析,Eg95可塑性較強的區域為:18-35,40-48,57-65,70-75,78-88,93-99,108-112,120-139。4)采用Jameson-Wolf方法對Eg95的抗原性指數進行預測,抗原性指數較高的表位區域為:17-38,41-48,55-67,68-90,92-99,103-113,120-133,136-138。聯合以上的參數和Eg95的二級結構進行綜合分析,得出Eg95有7個潛在的的B細胞抗原表位,分別為:17-38,41-48,57-67,68-90,92-99,104-113,120-133。
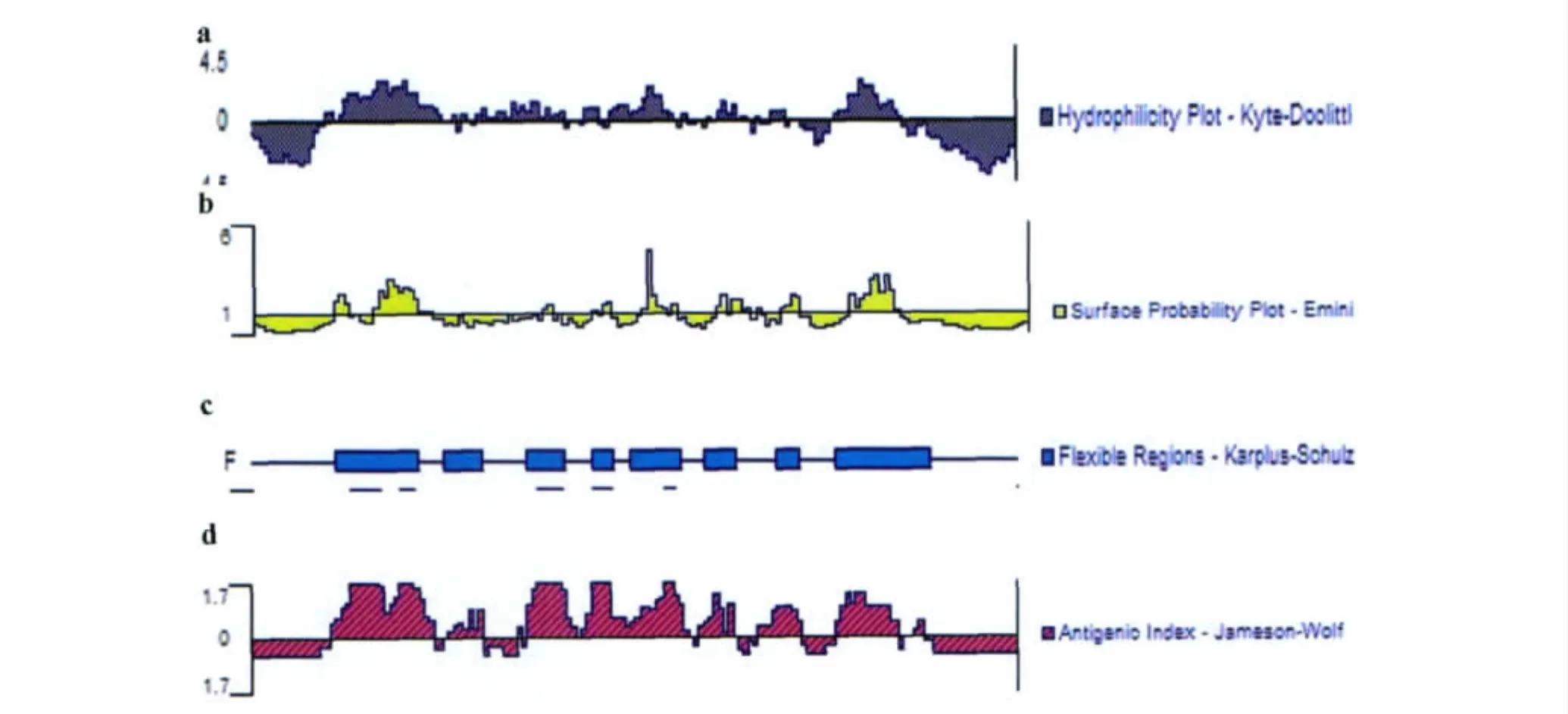
圖3 DNAStar分析Eg95的各項參數Fig.3 The parameters of Eg95using DNAStar analysis
2.1.4 在線網絡IEDB預測Eg95抗原B細胞表位 通過在線網絡IEDB對Eg95進行表位預測(見圖4):1)Chou& Fasmanβ轉角的預測,分值較高的區域為:19-49,56-65,68-78,82-91,93-100,121-140。2)對Eg95進行Emini表面可及性的分析,分值較高的區域為:16-22,23-37,68-73,78-88,92-103,107-111,120-130。3)在Karplus &Schulz柔韌性區域的預測中,Eg95分值高的區域為:18-37,40-47,57-65,70-75,78-89,92-100,109-113,120-139。4)通過Kolaskar &Tongaonkar抗原性指數預測,可能存在的表位區域為:4-22,39-48,50-59,65-80,86-94,101-121。5)Parker的親水性預測顯示Eg95的高親水性區域為:16-34,40-47,55-65,68-76,78-90,103-112,119-140。6)通過 Bepipred預測抗原表位,Eg95存在多個Bepipred線性抗原表位區域為:19-33,41-44,69-76,82-86,120-134。綜合以上參數分析,得出Eg95的潛在抗原表位區域也為 7個,分別是:16-37,41-47,50-65,68-90,92-100,103-112,120-139。
通過綜合以上兩個分析軟件的結果,將兩個軟件預測為表位的相同區域確定為最終Eg95的B細胞抗原表位區域,分別為:16-38,41-48,50-67,68-90,92-100,103-113,120-132。
2.2 T細胞表位預測結果
2.2.1 SYFPEITHI預測Eg95抗原T細胞表位
在進行T細胞表位的預測后,選取了分值高的13個T細胞抗原表位,預測的結果見表1。
2.2.2 在線網絡預測Eg95抗原T細胞表位 為了提高預測的準確性,同時用在線預測工具(http://bimas.dcrt.nih.gov/molbio/hla_bind/)對Eg95的T細胞表位進行預測,結果見表2。
綜合兩個在線軟件的預測,確定了在兩種方法中分值都比較高的七個表位為Eg95的T細胞表位區域,分別是:6-14,14-22,48-56,4-12,98-106,142-150,146-154。
3 討 論
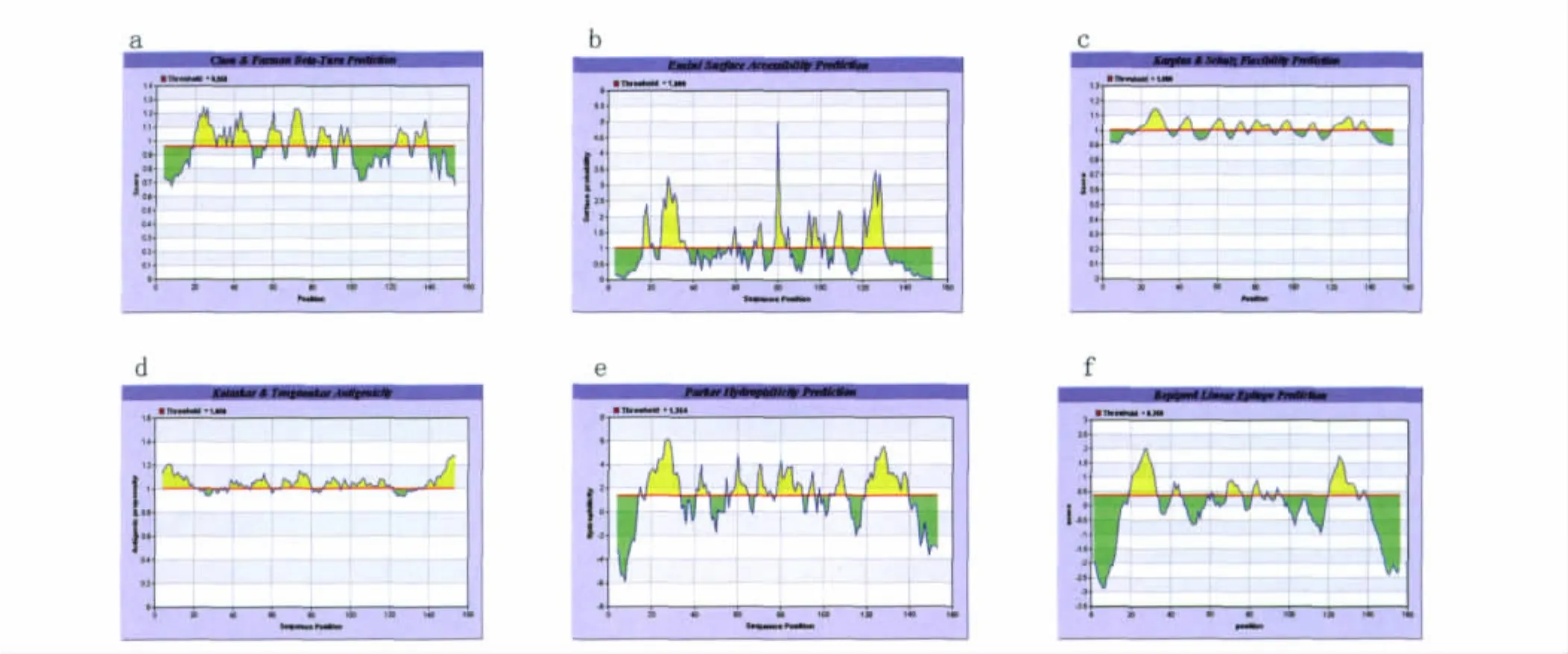
圖4 IEDB分析的Eg95的B細胞抗原表位Fig.4 The parameters of Eg95using IEDB analysis
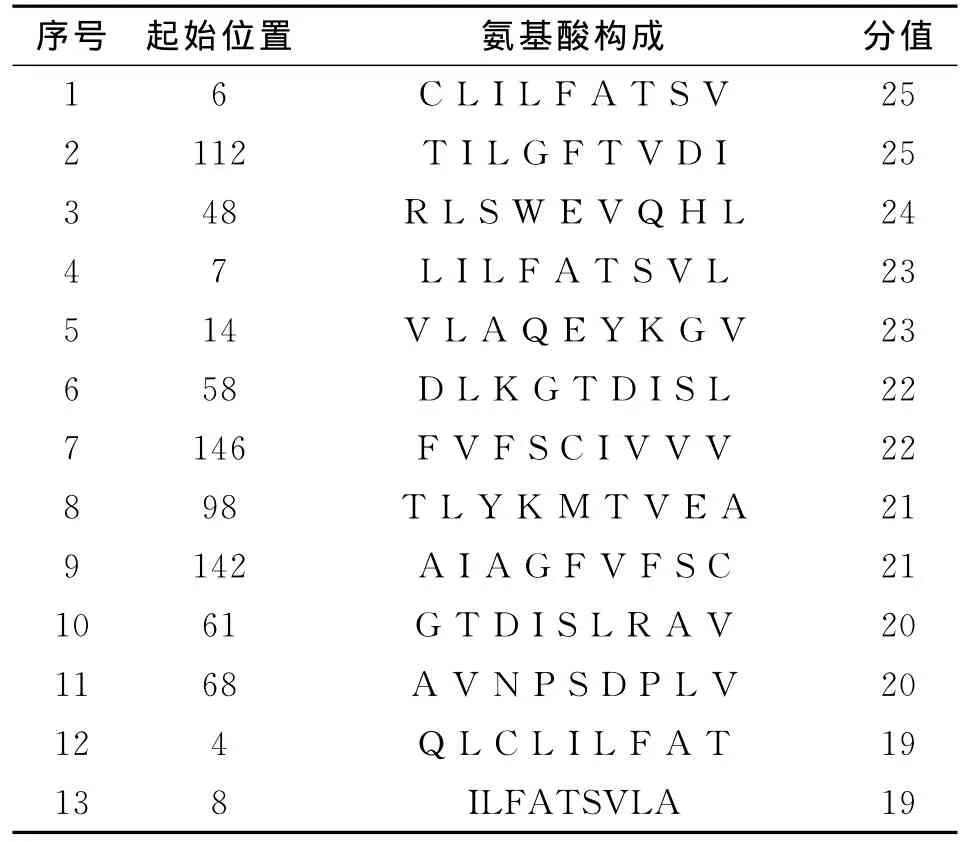
表1 SYFPEITHI預測的MHCⅠ類HLAA 0201限制性T細胞表位Table 1 HLA-A*0201nonamers T cell epitope using SYFPEITHI
寄生蟲病仍是發展中國家亟待解決的公共衛生問題,研制抗寄生蟲病疫苗是人們關注的熱點和難點問題。包蟲病嚴重影響我國西部牧區的經濟發展和人民的身體健康,而且新的感染和發生區域也在不斷擴大,因此通過免疫預防途徑控制和消滅包蟲病將是行之有效的方法。20世紀80年代Strohmaier[11]等發現口蹄疫病毒(FMDV)的 146-154及200-213氨基酸肽段含有免疫性位點,從而找到了一種新型的疫苗,即表位疫苗。表位(epitope)是抗原分子中決定抗原特異性的特殊化學基團,又稱抗原決定簇(antigenic determinant),它是T細胞抗原受體(Tcell receptor,TCR)和B細胞抗原受體(B cell receptor,BCR)及抗體特異性結合的基本單位,分為T細胞表位和B細胞表位。
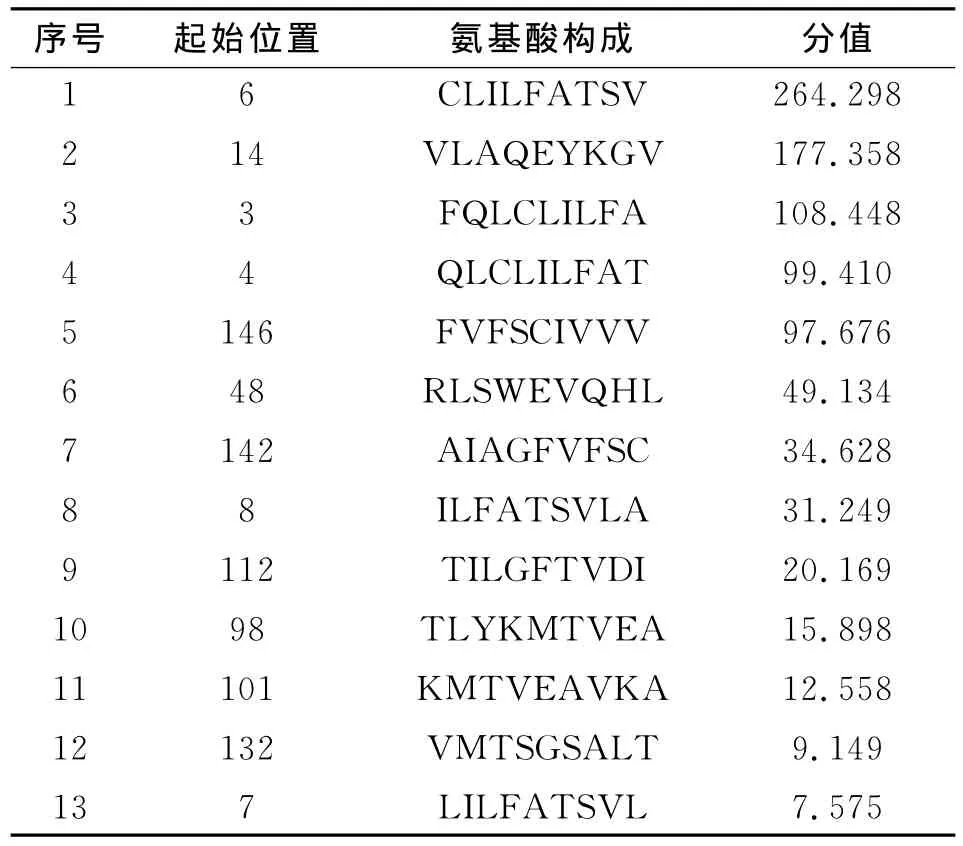
表2 在線網絡預測的MHCⅠ類HLAA 0201限制性T細胞表位Table 2 HLA-A*0201nonamers T cell epitope using Internet
分子生物學和免疫學研究快速向前發展,如何利用細胞表位預測新方法,高效快速地獲得表位的信息,已經成為抗包蟲病疫苗研究中的重要內容。傳統的篩選方法,包括噬菌體肽庫,核磁共振技術,質譜技術,SPR等多種手段都需要做大量的工作,花費大量的經費,但是預測的準確性卻有限,這讓人們感到困擾。隨著計算機知識的普及和網絡技術的發展,預測表位的參數計算變得容易操作。以往表位預測多采用單參數預測,有其局限性,隨著生物信息網絡技術的發展和數據庫的擴展,多參數多方法的綜合預測已成為當前表位預測的主流,可顯著提高預測的準確性[12]。李晉濤[13]運用多種方法對Eppin的二級結構和表面特性,如親水性、理化性質、可及性、免疫原性和可塑性等方面進行分析,得出其存在多個潛在的抗原表位。經過體外實驗證明,所預測抗原表位區域基本能與免疫抗血清發生抗原特異性反應。李章球[14]應用SYFPEITHI等7種軟件對人、鼠源血小板糖蛋白GPⅡb/Ⅲa抗體蛋白進行 HLA-A*0201、HLA-A*1101和 HLA-A*2401限制性表位預測,發現了GPⅡb/Ⅲa抗體蛋白的T細胞表位。
本研究利用生物信息網絡技術對Eg95抗原的B細胞表位預測,發現其跨膜結構中有一個跨膜區,一般比較穩定,不容易發生氨基酸的變異和表位形成。抗原的柔性區域主要集中在膜外區,本研究氨基酸序列也證實了這一結果。用TMHMM對Eg95抗原的跨膜序列進行分析,膜外區占了大部分的區域,說明這些區域產生抗原表位的可能性也比較大。用SOPMA Sever進行預測,發現Eg95氨基酸序列中柔性結構占氨基酸總數的43.59%,1-132區域是抗原表位富集的區域,表明Eg95的抗原表位也集中在該區域內。
在二級結構的預測中,蛋白質二級結構預測方案認為蛋白質二級結構分析與蛋白質表位的分布關系密切,無規則卷曲和β轉角主要位于蛋白質的表面,結構突出,有利于與抗體嵌合,成為抗原表位的可能性也較大[15]。本研究用SOPMA Sever對Eg95進一步分析發現其無規則卷曲占40.38%,β轉角為3.21%,表明在Eg95的這些區域中,抗原表位將容易產生并發揮特異性反應。
用抗原表位預測的經典方案對Eg95進行多參數的預測,(1)Hopp和 Woods為代表的蛋白質的親水性方案(hydrophilicity),此方案認為蛋白質抗原各氨基酸殘基可分為親水殘基和疏水殘基兩類。在機體內,疏水性殘基一般埋在蛋白內部,而親水性殘基位于表面,因此蛋白的親水部位與蛋白抗原表位有密切的聯系[16]。(2)Emini蛋白可及性方案(accessibility),指蛋白質抗原中氨基酸殘基被溶劑分子接觸的可能性,它反映了蛋白質抗原內、外各層殘基的分布情況[17]。(3)Karplus-Sohulz柔性區域預測方案(flexibility)認為蛋白抗原構象不是剛性不變的,其多肽鏈骨架有一定程度的活動性,也就是蛋白質的柔性區域。活動性強的氨基酸殘基即可塑性大的位點,易形成抗原表位[18]。(4)Jameson-Wolf的抗原性指數方案(Antigenic index)通過對20個已研究得很透的蛋白質的69個連續位點的606個氨基酸統計分析,用各氨基酸殘基在已知B細胞表位中出現的百分率與其通常在蛋白質中出現的百分率比值的對數建立了抗原性刻度,并以此計算蛋白中各亞序列的抗原性[19]。(5)β轉角的預測方案認為β轉角是蛋白質二級結構的基本類型之一,由4個氨基酸殘基組成,其中第一個殘基的CO基團和第四個殘基的NH基團之間形成氫鍵,使多肽鏈的方向發生U形改變。正因為這樣的改變,使表位容易在β轉角存在的區域形成。這些方案的綜合運用,將會提高B細胞表位預測的準確性。
在T細胞的預測中,MHCI類抗原表位預測的準確率較高,可達90%左右,MHCI類表位是由9個氨基酸殘基構成的短肽,漢族人種最常見的是HLAA 0201限制的抗原表位[20]。因此本研究對Eg95進行了HLAA 0201MHCI類9肽限制性表位預測。研究者們發現根據蛋白質氨基酸的分布,運用各種軟件對氨基酸的構成和分布進行計算得出一個T細胞表位分值,以表格的形式顯示[21]。本研究將兩種預測法分值排在前13位的表位進行綜合分析,發現兩種不同方法的結果有許多重復之處,說明綜合兩種方法的結果能夠提高預測方法的準確性。聯合各參數預測Eg95含有7個潛在的B細胞表位分別是16-38,41-48,50-67,68-90,92-100,103-113,120-132,7個分值較高的 T細胞表位是6-14,14-22,48-56,4-12,98-106,142-150,146-154。
本研究通過生物信息學預測的這些表位區域有利于下一步研究和篩選Eg95抗原B細胞和T細胞優勢抗原表位,構建優勢抗原表位疫苗,為獲得高效的保護性疫苗和預防包蟲病的流行奠定實驗基礎。
[1]劉春燕,馬秀敏,丁劍冰,等.我國棘球絳蟲感染的不同宿主狀況[J].中國人獸共患病學報,2009,25(6):586-588.
[2]王立英,伍衛平,朱雪花.2004-2008年全國包蟲病疫情分析[J].中國人獸共患病學報,2010,26(7):699-701.
[3]鮑佳春,袁鳳來,陸偉國.包蟲病治療進展[J].中國血吸蟲病防治雜志,2010,22(2):197-199.
[4]Ligtowlers MW,Lawrence SB,Gauci CG,et al.Vaccination against hydatidosis using a defined recombinant antigen[J].Parasite Immunol,1996,18(9):457-462.
[5]賈海英,馬秀敏,丁劍冰,等.細粒棘球蚴Eg95重組蛋白表達及其血清學反應的研究[J].中國人獸共患病學報,2009,25(4):344-347.
[6]Ben YT,Arnon R.Towards an epitope based human vaccine for influenza[J].Hum Vaccin,2005,1(3):95-101.
[7]Corradin G,Swpertini F,Verdini A.Medicinal application of long synthetic peptide technology[J].Expert Opin Biol Ther,2004,4(10):1629-1639.
[8]駱利敏,李明,夏虎,等.乙型肝炎病毒多表位抗原基因的設計、合成及表達[J].中華微生物學和免疫學雜志,2004,24(6):426-432.
[9]Wang X,Yang Q,Wang P,et al.Derp2-mutant gene vaccine inhibits airway inflammation and up-regulates Toll-like receptor 9 in an allergic asthmatic mouse model[J].Asian Pac J Allergy Immunol,2010,28(4):287-293.
[10]You L,Brusic V,Gallagher M,et al.Using Gaussian process with test rejection to detect T-Cell epitopes in pathogen genomes[J].IEEE/ACM Trans Comput Biol Bioinform,2010,7(4):741-751.
[11]Schuler MM,Nastke MD,Stevanovikc S.SYFPEITHI:database for searching and T-cell epitope prediction[J].Methods Mol Biol,2007,409:75-93.
[12]Sompuram SR,Bastas G,Vani1K,et al.Accurate identification of paraprotein antigen targets by epitope reconstruction[J].Blood,2008,111(1):302-308.
[13]李晉濤,陳正瓊,梁志清,等.Eppin抗原的二級結構分析B細胞表位預測[J].第三軍醫大學學報,2008,30(24):2254-2257.
[14]李章球,張梅霞,胡海燕,等.GPⅡb/Ⅲa抗體的二級結構及T、B細胞表位的預測[J].生物醫學工程學雜志,2010,27(5):1146-1151.
[15]Sikic K,Tomic S,Carugo O.Systematic comparison of crystal and NMR protein structures deposited in the protein data bank[J].Open Biochem J,2010,3(4):83-95.
[16]Sy SM,Chen J,Huen MS.The 53BP1-EXPAND1connection in chromatin structure regulation[J].Nucleus,2010,1(6):472-474.
[17]Liliana F',Bjarni H,Oliver K,et al.Epitope prediction algorithms for peptide-based vaccine design [J].Proc Comput Sys Bioinform,2003,33:7690-7695.
[18]Casella G,Ferrante F,Saielli G.A.A DFT study of the Karplus-type dependence of vicinal(3)J(Sn-C-X-C),X=N,O,S,in organotin(iv)compounds:application to conformationally flexible systems[J].Org Biomol Chem,2010,8(12):2711-2718.
[19]Ie SI,Thedja MD,Roni M,et al.Prediction of conformational changes by single mutation in the hepatitis B virus surface antigen(HBsAg)identified in HBsAg-negative blood donors[J].Virol J,2010,7:326-334.
[20]Yan C,Wang R,Li J,et al.HLA-A gene polymorphism defined by high-resolution sequence-based typing in 161Northern Chinese Han people[J].Gen Prot Bioinform,2003,1(4):304-309.
[21]Tong JC,Tan TW,Ranganathan S.Methods and protocols for prediction of immunogenic epitopes[J].Briefings in Bioninformatics,2007,8(2):96-108.
Bioinformatics prediction onEg95antigen epitopes ofEchinococcusgranulosus
LI Yu-jiao,WANG Jing,ZHAO Hui,JIA Hai-ying,LI Bo,MA Xiu-min,WEN Hao,DING Jian-bing
(MedicalResearchCenterandXinjiangKeyFirst,FirstAffiliatedHospital ofXinjiangMedicalUniversity,Urumqi830011,China)
The protein secondary structure was analyzed by TMHMM Sever2.0and SOPMA Sever,and B cell and T cell epitopes ofEg95were predicted by DNAStar,IEDB and SYFPEITHI.Many distinct antigenic epitopes ofEg95were identified by computation,and the possible B cell epitopes of which were in the regions of 16-38,41-48,50-67,68-90,92-100,103-113,and 120-132.The high score of T cell epitopes were in the regions of 6-14,14-22,48-56,4-12,98-106,142-150,and 146-154.These results indicated that there are 7antigen epitopes inEg95antigen determined by bioinformatics methods,which are significant in the further research ofEg95antigenic and development of dominant epitope vaccine.
Eg95;antigen epitope;bioinformatics
R383.3
A
1002-2694(2011)10-0892-05
*國家自然科學基金項目 (30901374,81060135,30560146,30860263)資助
丁劍冰,Email:djbing002@163.com
1.新疆醫科大學第一附屬醫院新疆重點實驗室,烏魯木齊 830011;
2010-11-17;
2011-03-23
猜你喜歡
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
科學(2020年5期)2020-11-26 08:19:22
軟件(2020年3期)2020-04-20 01:45:18
商周刊(2018年15期)2018-07-27 01:41:20
敦煌學輯刊(2018年1期)2018-07-09 05:46:42
北京教育·普教版(2017年1期)2017-02-05 13:26:23
新疆農墾科技(2016年2期)2016-08-21 13:50:16
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
新疆財經大學學報(2015年3期)2015-12-10 03:49:15
