河蜆擾動沉積物界面效應及其在水中代謝速率
2011-12-20 09:11:20商景閣王兆德申秋實范成新中國科學院南京地理與湖泊研究所湖泊與環境國家重點實驗室江蘇南京210008
中國環境科學 2011年6期
張 雷,商景閣,王兆德,申秋實,范成新 (中國科學院南京地理與湖泊研究所湖泊與環境國家重點實驗室,江蘇南京 210008)
河蜆擾動沉積物界面效應及其在水中代謝速率
張 雷,商景閣,王兆德,申秋實,范成新*(中國科學院南京地理與湖泊研究所湖泊與環境國家重點實驗室,江蘇南京 210008)
采用室內培養實驗,研究了河蜆擾動對沉積物耗氧速率與營養鹽通量的影響及河蜆在水中的呼吸與排泄速率.結果表明,河蜆增大了沉積物耗氧速率與溶解活性磷(SRP)、NH4+、NO3-向上覆水釋放通量.河蜆在沉積物-水系統中產生NO3
-速率與其在水中產生NO3
-速率不存在顯著差異.而河蜆在水中呼吸速率是其在沉積物-水中所產生凈耗氧速率的4.3倍,河蜆在水中排泄SRP、NH4+速率分別為其在沉積物-水中所產生凈SRP、NH4+釋放速率的7.3倍與20倍.這些顯著差異可能是由于河蜆在水中與沉積物中的活動形態不同.
河蜆;沉積物-水界面;氧氣;溶解活性磷;氨氮;硝酸根
底棲動物通過攝食、構筑洞穴、通風、排泄等活動,影響 O2在沉積物中的分布[1-2]與消耗速率[3-4]、以及營養鹽、重金屬在沉積物-水界面的遷移、轉化過程[5-8].河蜆(Corbicula fluminea)是一種穴居于淡水、咸淡水底沉積物表層的雙殼類軟體動物[9-10],可以在水底高密度聚集,成為底棲生物群落的優勢種,控制水底生物分布等[11-13].河蜆擾動對O2、溶解態營養鹽在沉積物-水界面遷移過程的影響[14-17],以及河蜆在水中的呼吸與排泄速率等方面的研究,受到一些學者的關注
[12,18-19],而這兩者之間又存有怎樣的關系尚不清楚.
本研究基于這一出發點,通過室內培養實驗,研究河蜆在沉積物-水系統中對沉積物耗氧速率、營養鹽在界面通量的影響,以及河蜆在水中的呼吸與排泄速率,旨在揭示2種不同環境下氧消耗速率與營養鹽釋放速率的差異.
1 材料與方法
1.1 野外采樣
河蜆在太湖分布廣泛,其中貢湖灣和西部湖區密度最高[20-21].2008年11月12日利用大口徑
* 責任作者, 研究員, cxfan@niglas.ac.cn重力采樣器(直徑110mm,長500mm)在太湖馬山南(31°21′45.4″N, 120°8′18.7″E)采集沉積物柱樣,同時采集湖水用于室內培養.利用彼得森采樣器采集表層沉積物、收集河蜆,并取一些沉積物與湖水將收集的河蜆暫養.所有樣品在采集后立即運回實驗室處理、培養.
1.2 實驗設計
將所采集沉積物柱樣從表層至層深12cm切分成6層,每層2cm.把相同層位的沉積物收集在一起,然后過 30目篩以去除其中河蜆等底棲生物、大顆粒物.將過篩后的各層沉積物分別混勻,按其原順序裝填到下端密封的有機玻璃管(直徑110mm×長170mm)中,每層深2cm.然后利用醫用輸液器把所采集湖水輕輕地引到沉積物上.共制得 6根沉積物柱樣,每根沉積物深 12cm,上覆水深5cm.將制得柱樣放在40 L培養水槽內,加湖水淹沒培養,每 3d更換一次湖水,并用微型曝氣頭充氣預培養 16d,以使沉積物性質穩定.將所采集河蜆放在鋪有沉積物的水槽中暫養.同時取0~2cm沉積物分析其基本理化性質.
在17d,將6根柱樣隨機分成2組,每組3根.其中一組為對照組(C);另外一組為河蜆擾動組(B).在17d測定沉積物耗氧速率;第18d,測定氨氮(NH4+)、硝酸根(NO3-)與溶解活性磷(SRP)在沉積物-水界面通量.根據前期調查所得太湖最高河蜆密度(1072ind./m2)與生物量(2918g/m2)[17],在實驗第19d,向河蜆組各柱樣表面輕輕引入10只個體中等大小(2.48~3.22g/只,體長 1.82~2.19cm)河蜆,每組10只河蜆總濕重為(29.0±0.1)g.柱樣繼續放在水槽中淹沒充氣培養20d,每3d更換一次湖水.在第37,38d分別測定耗氧速率與界面營養鹽通量.實驗第39d,將河蜆從各柱樣中分別挑出,依然保持3組,測定其在水中呼吸速率與排泄速率(在后文圖中用R表示).在整個實驗過程中,實驗室溫度保持在(15±1)℃,與采樣時沉積物-水界面處溫度一致.
測定沉積物耗氧速率、沉積物-水界面營養鹽通量時,首先將水槽水位調至低于培養管上端.測耗氧速率時,利用虹吸法從上覆水中下部吸取上覆水進入微碘量瓶(50mL),讓水溢流50mL以排除空氣干擾,然后迅速固定水中溶解氧(DO).同時向柱樣中補充100mL水槽中的培養水,用一具O型密封圈塞子將上覆水端密封.2h后再次取樣分析 DO.界面營養鹽通量通過測定上覆水中NH4+、NO3
-、SRP隨時間的變化實現.每次用注射器在沉積物-水界面上方0.5cm處抽取15mL上覆水,并用15mL培養水補充被抽掉的上覆水.迅速將水樣用0.45μm醋酸纖維濾膜過濾,4℃冷藏待分析.測定通量時,每 0.5h取樣一次,每測定一次通量取樣5次.為減小浮游生物等干擾,耗氧速率及營養鹽通量測定在黑暗環境中進行.河蜆在水中呼吸、排泄速率的測定方法為:將河蜆沖洗干凈,3組河蜆分別放入3只500mL三角瓶中,加入湖水密塞;同時另取3只500mL三角瓶加滿湖水作為空白對照樣,黑暗處放置 2h,測定密封前后湖水中O2、NH4+、NO3-、SRP的濃度.
1.3 樣品分析
將新鮮沉積物在60℃條件下烘至恒重,根據前后質量差計算沉積物含水率.將烘干的沉積物磨碎、過100目篩后放550℃馬弗爐中灼燒5h,通過質量差計算燒失重(LOI).沉積物中總氮與總磷分別采用堿性過硫酸鉀消解法與SMT法[22].水中DO測定采用微碘量法[23].水中SRP、NH4+、NO3
-分別采用鉬銻抗光度法、納氏試劑分光光度法與紫外分光光度法測定[24].
1.4 數據處理與統計分析
測定通量過程中不斷用培養水補充被抽掉的上覆水,則需對所測定上覆水濃度根據式(1)進行校正:
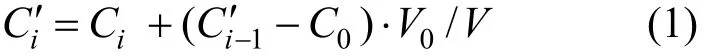
式中:Ci′是第 i(≥2)次校正后的上覆水濃度, μmol/L;Ci是第 i(≥1) 次測定出來的上覆水濃度, μmol/L; C0是培養水濃度,μmol/L;V與V0分別是柱樣上覆水體積(L)與每次所抽取水樣體積(L).將所獲得校正數據對時間作圖,得濃度對時間的回歸斜率;然后利用式(2)計算通量:

式中: F表示溶解物質在沉積物-水界面通量, μmol/(h?m2); k為該溶解物質在上覆水中濃度對時間的回歸斜率,μmol/(L·h); V為上覆水體積,L; A為沉積物柱樣表面積,m2.在沉積物耗氧速率與河蜆在水中代謝速率計算中略去濃度校正步驟,直接利用式(2)計算.
沉積物耗氧速率、界面通量與呼吸、代謝速率結果均表示為:平均值 ± 標準偏差 (n=3)的形式.河蜆在沉積物-水界面所產生的擾動效應(耗氧速率,SRP、NH4+與NO3-通量)、及其在水中呼吸、排泄速率均表示為μmol/(h?g)的形式,其中河蜆重量為去殼干重,根據所添加河蜆濕重、及河蜆去殼干重與濕重比為 0.024[17]換算得出.采用單因子方差分析(one-way ANOVA)檢驗C組與B組、B-C與R之間的差異,統計分析利用軟件SPSS(13.0)完成.P>0.05表示差異不顯著, P<0.05
表示差異顯著,P<0.01表示差異高度顯著,
P<0.001表示差異極顯著.
2 結果
表層 0~2cm沉積物含水率 49.3%,LOI為4.72%,總氮、總磷含量分別為 12.8,51.2μmol/g.被引入到沉積物表面上后,河蜆開始挖穴.其活動破壞了沉積物表層原有結構,使得一些厭氧沉積物被上覆水所氧化.隨著其對環境的適應,河蜆逐漸穩定下來,開始穴居生活,偶爾可以觀察到其運動或伸出虹吸管.在實驗后期測定河蜆在水中呼吸與排泄速率時,發現河蜆瓣膜經常處于張開狀態.

圖1 引入河蜆前沉積物耗氧速率與營養鹽通量Fig.1 Sediment oxygen uptake rates and nutrient fluxes before the introduction of C. fluminea
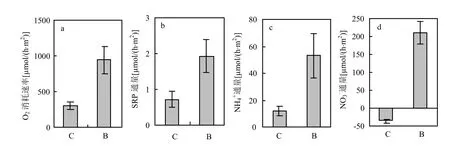
圖2 引入河蜆后沉積物耗氧速率與營養鹽通量Fig.2 Sediment oxygen uptake rates and nutrient fluxes after the introduction of C. fluminea
在引入河蜆前(圖1),對照組與河蜆擾動組沉積物耗氧速率不存在顯著差異(P>0.05). SRP、NH4+在沉積物-水界面通量向上,呈釋放狀態,兩組之間差異不顯著(P>0.05).與SRP、NH4+呈向上釋放不同, NO3-通量向下,表現為沉積物吸收上覆水中NO3-,對照組與河蜆擾動組差異也不顯著(P >0.05).
河蜆顯著改變了沉積物耗氧速率及營養鹽在沉積物-水界面通量(圖 2).河蜆擾動組沉積物耗氧速率是對照組的 3.1倍,兩者差異顯著(P<0.01).河蜆顯著促進了 SRP、NH4+向上覆水的釋放,河蜆組SRP、NH4+通量分別是對照組的2.6倍(P<0.05)與 4.4倍(P<0.05).同時,河蜆改變了NO3-在沉積物-水界面通量方向,促進 NO3-向上覆水釋放(P<0.001).
河蜆在沉積物中所增大 O2消耗速率為9.2μmol/(h?g),僅為其在水中呼吸速率[39.9μmol/ (h?g)]的0.23倍(P<0.001)(圖3).河蜆在沉積物-水系統中所產生凈 SRP釋放速率為 0.017μmol/ (h?g),而其在水中排泄SRP速率則為此速率的20倍(P<0.01).河蜆在水中排泄 NH4+速率為4.3μmol/(h?g),為其在沉積物-水界面所產生凈NH4+釋放速率[0.59μmol/(h?g)]的 7.3倍(P<0.01).河蜆在沉積物-水界面所產生凈NO3-釋放速率為(3.5±0.45)μmol/(h?g),其在水中所產生硝酸根速率為(3.0±0.65)μmol/(h?g),兩者差異不顯著(P >0.05).

圖3 河蜆在沉積物-水系統、水中分別產生的耗氧速率、營養鹽釋放速率Fig.3 Sediment oxygen uptake and nutrient release rates caused by C. fluminea in sediment-water system and water separately
3 討論
實驗結果表明,15℃時河蜆在水中呼吸速率為39.9μmol/(h?g),與Hakenkamp等[16]在18℃時所得49.4μmol/(h?g)在同一數量級內,兩者之間的差異可能是因為實驗溫度不同.隨著溫度升高,河蜆呼吸速率增大,例如:溫度由 20°C 升高到28°C,一只濕重 1.7g河蜆的呼吸速率增大了40%[18].河蜆增大了沉積物耗氧速率(圖 2a),這與關于其他雙殼動物的研究結果一致[3,25].底棲動物增大沉積物耗氧速率的原因包括以下 2個方面:一是底棲動物自身的呼吸作用,二是生物擾動所引起的厭氧沉積物氧化、沉積物中微生物活性增強等[4].底棲動物的呼吸速率常常通過與本實驗類似的方法測定,然后將有擾動的沉積物耗氧速率減去對照樣耗氧速率與底棲動物呼吸速率,即得生物擾動本身所引起沉積物耗氧速率的增加量[4,8,26].因此,底棲生物的呼吸速率應該小于其在沉積物中所引起的耗氧速率增大量.但本實驗結果卻表明河蜆在水中的呼吸速率遠大于河蜆所引起沉積物耗氧速率的增加量(圖 3a).產生這一結果的原因可能是因為河蜆在沉積物-水系統與其在水中的生活、攝食方式不同.
河蜆是一種雙食性底棲動物,在沉積物-水系統中,它們既可以通過虹吸管從水中濾食有機顆粒,也可以通過斧足從沉積物中攝食有機質,而在水中,它只能通過濾食維持生命[14,16].既能夠通過斧足攝食又能濾食的河蜆,比僅能濾食的河蜆長的快[16],同時有研究表明單獨濾食所提供能量不超過河蜆總能量需求的51%[27].為保持其身體代謝的高能量需求,在水中的河蜆必須張開瓣膜加大其攝食力度.在沉積物中,河蜆通過間歇新陳代謝維持生命,它們通常持續閉合瓣膜 5~10h,閉合時間長達幾天的情況也有發生[28].但在水中,河蜆閉合瓣膜的時間不會超過7min[29].這與實驗中觀察河蜆在沉積物中與水中的活動情況一致.而河蜆張開瓣膜時其呼吸、代謝速率是瓣膜閉合時的 10倍[28].因此,河蜆在水中的呼吸速率要比其在沉積物中高出許多.從圖 3a的數據分析,河蜆在水中呼吸速率至少為其在沉積物中呼吸速率的4.3倍.
Wittmann等[19]研究結果表明,1只長 2.0cm (本研究使用河蜆平均體長)河蜆排泄 SRP與NH4+速率分別為 1.1,14.9μmol/d.基于上述數據,假設相同體長河蜆的重量、排泄量相同,那么河蜆排泄 SRP與 NH4+的速率可以轉換為 0.69, 9.48μmol/(h?g).本研究所得河蜆在水中排泄SRP、NH4+速率分別為 0.34,4.3μmol/(h?g)(圖3b,c),相對偏小.這可能是因為本研究實驗溫度15℃,Wittmann等研究溫度為 19℃,與其呼吸速率隨溫度的變化一樣,溫度升高河蜆排泄SRP與NH4+速率也會顯著增大[12,18-19].
NH4+、SRP在沉積物-水界面釋放通量不僅受河蜆排泄速率的影響,同時受河蜆擾動過程的影響.生物擾動能促進有機質礦化,產生的SRP、NH4+富集在間隙水中或釋放到上覆水中[8].河蜆擾動增大沉積物-水界面面積,有利于間隙水中SRP、NH4+向上覆水擴散;同時擾動可以在間隙水與上覆水之間產生平流作用,這一過程也有利于兩者的釋放[30].雖然河蜆擾動能夠增加氧化沉積物體積,伴隨加強的硝化、二價鐵氧化等過程存在減小間隙水中NH4+、SRP濃度的可能,但由于機質礦化產生SRP、NH4+過程的存在,減小程度往往不顯著[17].因此,總體而言河蜆自身代謝及其擾動本身對沉積物所產生的影響均是有利于SRP、NH4+向上覆水釋放.由于河蜆在水中代謝速率約為其在沉積物中的 10倍[28],其在水中排泄速率也得到加強.這正是河蜆在水中排泄NH4+、SRP速率為其在沉積物中所產生凈NH4+、SRP釋放速率7.3倍與20倍的原因.
河蜆在水中的活動增大了NO3-濃度(圖3d),這在前人研究中未被關注.其他一些雙殼類動物在排泄過程中產生NO3-[31],這可能是因為它們殼與內部組織上繁殖有大量的硝化細菌[32].河蜆改變了NO3-在沉積物-水界面通量方向,促進NO3-向上覆水釋放(圖 2d).這主要是因為河蜆的擾動及排泄為硝化反應提供了充足的氨氮,同時河蜆擾動加深了O2在沉積物中的滲透,擴展了硝化細菌活動范圍并改善硝化活動進行的環境條件[15].結果還表明,河蜆在沉積物-水系統中產生的凈NO3-釋放速率與其在水中產生 NO3-的速率不存在顯著差異(圖 3d).這可能是因為河蜆在水中升高NO3-濃度是因為其殼與內部組織上附著硝化細菌,即通過強化硝化反應實現水中NO3-濃度升高,與河蜆自身排泄關系不大.
本實驗結果表明,不可用在水中測得河蜆呼吸、排泄速率代替其在沉積物中所產生的耗氧速率與營養鹽通量;在研究河蜆對沉積物-水界面氧消耗速率及營養鹽通量的影響時,也不可用在水中測定的呼吸、排泄速率來代替其在沉積物中的呼吸、排泄速率.如何確定河蜆及類似底棲動物在沉積物中呼吸、排泄速率,及其活動本身對沉積物耗氧速率及界面營養鹽循環過程的貢獻,需在后續研究中加強.
4 結論
4.1 河蜆增大了沉積物耗氧速率、增大了沉積物中SRP、NH4+、NO3-向上覆水釋放通量.
4.2 與河蜆在沉積物-水系統中所產生的凈耗氧速率、凈SRP、NH4+釋放速率相比,河蜆在水中呼吸速率、SRP與 NH4+排泄速率分別為其4.3、20與7.3倍,差異顯著.而河蜆在沉積物-水系統中所產生的凈 NO3-釋放速率與其在水中產生NO3-的速率不存在顯著差異.
[1] Wang F, Tessier A, Hars L. Oxygen measurements in the burrows of freshwater insects [J]. Freshwater Biology, 2001,46(2):317-327.
[2] Polerecky L, Volkenborn N, Stief P. High temporal resolution oxygen imaging in bioirrigated sediments [J]. Environmental Science and Technology, 2006,40(18):5763-5769.
[3] Michaud E, Desrosiers G, Mermillod-Blondin F, et al. The functional group approach to bioturbation: The effects of biodiffusers and gallery-diffusers of the Macoma balthica community on sediment oxygen uptake [J]. Journal of Experimental Marine Biology and Ecology, 2005,326:77-88.
[4] Glud R N. Oxygen dynamics of marine sediments [J]. Marine Biology Research, 2008,4:243-289.
[5] Andersson G, Granéli W, Stenson J. The influence of animals on phosphorus cycling in lake ecosystems [J]. Hydrobiologia, 1988,170:267-284.
[6] 路永正,閻百興.顫蚓擾動作用對鉛鎘在沉積物-水相中遷移的影響 [J]. 中國環境科學, 2010,30(2):251-255.
[7] Vanni M J. Nutrient cycling by animals in freshwater ecosystems [J]. Annual Review of Ecology and Systematics, 2002,33:341-370.
[8] Welsh D T. It’s a dirty job but someone has to do it: the role of marine benthic macrofauna in organic matter turnover and nutrient recycling to the water column [J]. Chemistry and Ecology, 2003,19:321-342.
[9] 張虎才,陳 玥,樊紅芳,等.河蜆分布的氣候環境及殼體穩定同位素 [J]. 海洋地質與第四紀地質, 2007,27(3):77-84.
[10] 劉 敏,熊邦喜.河蜆的生態習性及其對重金屬的富集作用 [J].安徽農業科學, 2008,36(1):221-224.
[11] Cherry D L, Roy R L, Lechleitner R A, et al. Corbicula fouling and control measures at the Celco Plant, Virginia [J]. American Malacological Bulletin, 1986,2:69-81.
[12] Lauritsen D D, Mozley S C. Nutrient excretion by the Asiatic clam Corbicula fluminea [J]. Journal of North American Benthological Society, 1989,8:134-139.
[13] Poff N, Palmer M, Angermeier P, et al. Size structure of the metazoan community in a Piedmont stream [J]. Oecologia, 1993, 95(2): 202-209.
[14] Vaughn C C, Hakenkamp C C. The functional role of burrowing bivalves in freshwater ecosystems [J]. Freshwater Biology, 2001,46:1431-1446.
[15] 陳振樓,劉 杰,許世遠,等.大型底棲動物對長江口潮灘沉積物-水界面無機氮交換的影響 [J]. 環境科學, 2005,26(6):43-50.
[16] Hakenkamp C C, Palmer M A. Introduced bivalves in freshwater ecosystems: the impact of Corbicula on organic matter dynamics in a sandy stream [J]. Oecologia, 1999,119:445-451.
[17] 張 雷.生物擾動下湖泊沉積物-水界面特征變化與磷的遷移轉化 [D]. 南京:中國科學院南京地理與湖泊研究所, 2010.
[18] 劉其根,沈和定,周洪琪,等.河蜆的耗氧率與排氨率 [J]. 上海水產大學學報, 1999,8(4):298-303.
[19] Wittmann M, Reuter J, Schladow G, et al. Asian clam (Corbicula fluminea) of Lake Tahoe: Preliminary scientific findings in support of a management [EB/OL]. http://169.237.166.248/ research/AsianClam2009.pdf.
[20] 蔡永久,龔志軍,秦伯強.太湖大型底棲動物群落結構及多樣性[J]. 生物多樣性, 2010,18(1):50-59.
[21] 蔡 煒,蔡永久,龔志軍,等.太湖河蜆時空格局 [J]. 湖泊科學, 2010,22(5):714-722.
[22] Ruban V, López-Sánchez J F, Pardo P, et al. Selection and evaluation of sequential extraction procedures for the determination of phosphorus forms in lake sediment [J]. Journal of Environmental Monitoring, 1999,1(1):51-56.
[23] USEPA-LG501-2007 Standard operating procedure for dissolved oxygen micro method, Winkler titration [S].
[24] 國家環境保護總局水和廢水檢測分析方法編委會. 水和廢水檢測分析方法 [M]. 4版.北京:中國環境科學出版社, 2002: 243-285.
[25] Karlson K, Hulth S, Ringdahl K, et al. Experimental recolonisation of Baltic Sea reduced sediments: survival of benthic macrofauna and effects on nutrient cycling [J]. Marine Ecology Progress Series, 2005,294:35-49.
[26] Svensson J M, Leonardson L. Effects of bioturbation by tube-dwelling chironomid larvae on oxygen uptake and denitrification in eutrophic lake sediments [J]. Freshwater Biology, 1996,35(2):289-300.
[27] Boltovskoy D, Izaguirre I, Correa N. Feeding selectivity of Corbicula fluminea (Bivalvia) on natural phytoplankton [J]. Hydrobiologia, 1995,312:171-182.
[28] Ortmann C, Grieshaber M K. Energy metabolism and valve closure behaviour in the Asian clam Corbicula fluminea [J]. The Journal of Experimental Biology, 2003,206:4167-4178.
[29] Saloom M E, Duncan R S. Low dissolved oxygen levels reduce anti-predation behaviours of the freshwater clam Corbicula fluminea [J]. Freshwater Biology, 2005,50:1233-1238.
[30] Kristensen E. Organic matter diagenesis at the oxic/anoxic interface in coastal marine sediments, with emphasis on the role of burrowing animals [J]. Hydrobiologia, 2000,426(1):1-24.
[31] Magni P, Montani S, Takada C, et al. Temporal scaling and relevance of bivalve nutrient excretion on a tidal flat of the Seto Inland Sea, Japan [J]. Marine Ecology Progress Series, 2000,198: 139-155.
[32] Welsh D T, Castadelli G. Bacterial nitrification activity directly associated with isolated benthic marine animals [J]. Marine Biology, 2004,144:1029-1037.
Bioturbation effects of Corbicula fluminea on sediment-water interface progresses and its metabolic rate in water.
ZHANG Lei, SHANG Jing-ge, WANG Zhao-de, SHEN Qiu-shi, FAN Cheng-xin*(State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China). China Environmental Science, 2011,31(6):1001~1006
A laboratory incubation experiment was carried out to investigate the effects of Corbicula fluminea bioturbation on sediment oxygen uptake and nutrient fluxes across the sediment-water interface, and the respiratory rate and excretion rate of C. fluminea in water. Sediment oxygen uptake rate and SRP, NH4+and NO3-fluxes from sediment to the overlying water were promoted by C. fluminea bioturbation. The producing rate of NO3-from C. fluminea activities in water was not significantly different from that in the sediment-water system. The respiratory rate of C. fluminea in water was 4.3 times as much as the net oxygen uptake caused by C. fluminea in the sediment-water system. Furthermore, the excretion rates of SRP and NH4+in water were 7.3 and 20 times as much as their net release rates caused by C. fluminea in the sediment-water system, respectively. The above differences between in water and the sediment-water system may be likely due to different living styles of C. fluminea in the two environments.
Corbicula fluminea;sediment-water interface;oxygen;soluble reactive phosphorus;ammonium;nitrate
X174
A
1000-6923(2011)06-1001-06
2010-10-08
國家自然科學基金資助項目(40730528,50979102);江蘇省社會發展項目(BE2009603);江蘇省自然科學基金項目(BK2009332)
張 雷(1982-),男,安徽宿州人,中國科學院南京地理與湖泊研究所博士研究生,主要從事湖泊沉積物-水界面環境地球化學研究.發表論文10篇.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
當代陜西(2020年13期)2020-08-24 08:22:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
制造技術與機床(2017年5期)2018-01-19 02:49:17
金秋(2017年4期)2017-06-07 08:22:16
中國材料進展(2016年10期)2016-12-26 06:50:20
濰坊學院學報(2016年2期)2016-12-01 13:00:11
新聞傳播(2015年11期)2015-07-18 11:15:04
