抗HP嗜酸乳桿菌與SGC-7901細胞的黏附及其對HP與SGC-7901細胞黏附的抑制效應
2012-02-24 07:46:46趙忠巖王江濱劉冬琦
中國實驗診斷學 2012年6期
關鍵詞:研究
趙忠巖,王江濱,劉冬琦,李 巖
(吉林大學1.中日聯誼醫院a.急救醫學科;b.消化內科,吉林 長春 130033;2.公共衛生學院)
*通訊作者
幽門螺桿菌是人類最常見的病原性細菌,是慢性胃炎、消化性潰瘍等疾病發生的主要病因,并與胃腺癌、胃粘膜相關淋巴組織淋巴瘤的發生密切相關,WHO已將其列為一類致癌因子[1]。由于目前根除HP治療方案存在許多弊端,近年來人們逐漸開始關注微生態療法在HP根除治療中的應用,有研究發現,益生菌中的某些菌株對HP的生長具有抑制作用[2],本實驗室經過大量的實驗研究從健康人糞便中篩選并建立了具有拮抗HP國際標準菌株SS1和NCTC11637作用的嗜酸乳桿菌L6,對其耐酸、耐堿等生物學特性及其在胃內存留時間和毒理進行了深入的研究[3],證明該株嗜酸乳桿菌具有穩定的抑制HP國際標準菌株作用。本研究旨在探討該株嗜酸乳桿菌拮抗幽門螺桿菌作用的可能機制。
1 資料與方法
1.1 嗜酸乳桿菌L6的建立及培養方法 L6由本實驗室通過從健康體檢者糞便中篩選而建立,采用固體培養法傳代培養,收集細菌后調整細菌濃度分別為9logCFU/ml、8logCFU/ml、7logCFU/ml。
1.2 SGC-7901細胞株及培養方法 SGC-7901細胞株購自中國科學院上海細胞生物研究所。細胞培養基為含10%胎牛血清的RPMI1640(Gibco BRL)完全培養液,細胞置于含5%CO2、95%空氣的CO2孵箱中,在37℃、95%濕度條件下培養。
1.3 L6與SGC-7901細胞的黏附及其對細胞存活率的影響 將SGC-7901細胞接種于內置載玻片的含20%胎牛血清的RPMI1640培養液的6孔培養板中,待細胞密度達80%時,用RPMI1640沖洗2次,分別加入終濃度為7logCFU/ml、8logCFU/ml和9logCFU/ml的嗜酸乳桿菌L6 PMI1640培養液,在37℃,5%CO2環境中培養,分別于1、6、12、和24 h終止培養,取出載玻片,PBS漂洗2次,自然干燥,甲醇固定15 min,革蘭染色,油鏡下隨機選擇50個細胞,計算其可視性細胞表面的細菌數,計算黏附指數(黏附細菌數/細胞數×100%)的均數及標準差,同時選擇與L6粘附的SGC-7901細胞以臺盼藍進行染色并計數其細胞總數和存活細胞數,換算成細胞存活率(活細胞數/細胞總數x 100%)。
1.4 嗜酸乳桿菌L6對HP與SGC-7901細胞黏附的拮抗效應 將已有SGC-7901細胞貼壁生長的載玻片置于6孔培養板中,當SGC-7901細胞密度達80%時,以下述3種方法分別觀察嗜酸乳桿菌L6對HP的拮抗效應。(1)排除實驗:先將不同濃度的嗜酸乳桿菌L6分別加入6孔培養板上(37℃,5%CO2)30 min,洗滌去除未黏附的L6,然后再加入HP懸液,繼續孵育30 min后計數HP黏附指數。(2)競爭實驗:將HP懸液與不同濃度L6同時加入6孔培養板中混勻,1 h后洗滌3次,以后處理方法同上。(3)替換實驗:先將HP懸液加入6孔培養板中(37℃,5%CO2),30 min后,洗滌去除未黏附的HP,然后再加入不同濃度的L6,混勻后繼續孵育30 min,處理方法同上。
1.5 上清液中IL-8的測定 將SGC-7901細胞懸浮在24孔板RPMI1640培養液中,37℃,5%CO2環境中培養24 h后,將HP與不同濃度的嗜酸乳桿菌L6加入培養液中共同培養,分別于培養6、12、24 h后取上清液通過ELISA方法測定IL-8的含量,并與單獨加入HP和單獨加入9logCFU/ml嗜酸乳桿菌進行比較。
1.6 統計學分析 所有數據均采用方差分析及t檢驗進行統計學處理,P<0.05為有統計學意義。
2 結果
2.1 嗜酸乳桿菌L6具有與SGC-7901細胞黏附的能力,其黏附指數在一定時間范圍內隨作用時間的延長而增高,在黏附后12 h達高峰。此外,黏附指數還隨細菌濃度的增加而增高,與對照組比較有顯著性差異,P<0.05,見表1。
2.2 嗜酸乳桿菌 黏附 、12和 的7901細胞的存活率與未被黏附的細胞存活率相比無顯著差異,且細胞存活率不受細菌濃度及黏附時間的影響,見表2。
2.3 嗜酸乳桿菌L6對HP與SGC-7901細胞的黏附具有明顯的拮抗作用,且這種拮抗作用隨細菌濃度的增加而逐漸增強,與單純HP組比較差異顯著,P<0.05;與L6的作用時間早晚有一定關系,排除實驗比替換實驗明顯增強,兩者比較差異顯著,P<0.05,見表3。
2.4 嗜酸乳桿菌L6可以明顯拮抗HP誘導SGC-7901細胞產生IL-8,且隨著細菌濃度的增加和作用時間的延長,其對HP誘導的IL-8分泌的拮抗作用逐漸增強,12 h后,濃度為9logCFU/ml組明顯高于濃度為7logCFU/ml組和濃度為8logCFU/ml組,各組比較差異顯著,P<0.05;而嗜酸乳桿菌L6本身幾乎不刺激上皮細胞分泌IL-8,見表4。

表1 嗜酸乳桿菌L6與SGC-7901細胞粘附能力分析
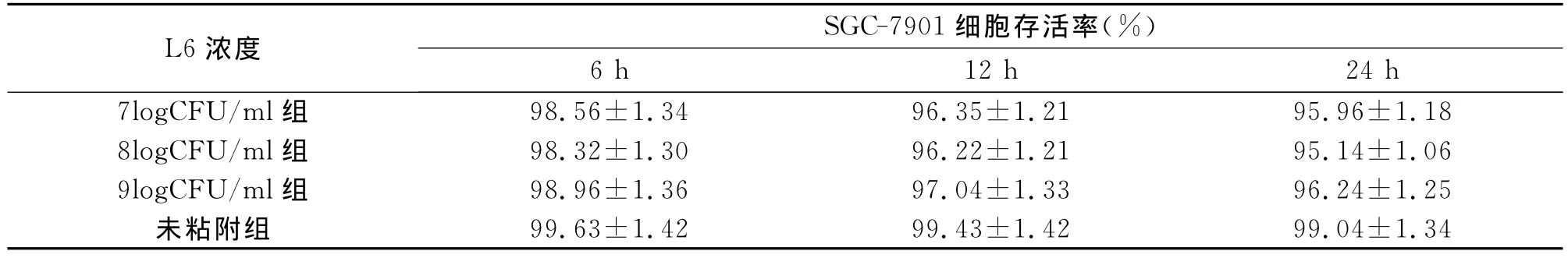
表2 嗜酸乳桿菌L6粘附SGC-7901細胞后細胞存活率的比較

表3 嗜酸乳桿菌L6對HP與SGC-7901細胞粘附拮抗作用分析
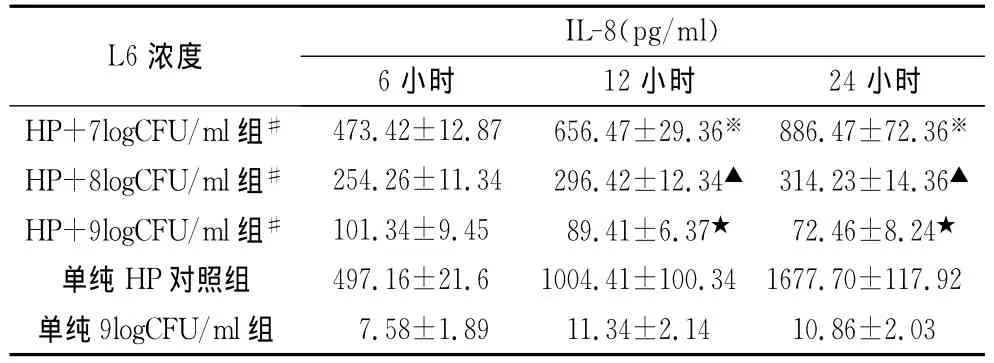
表4 嗜酸乳桿菌L6對HP誘導IL-8分泌的抑制效應
3 討論
幽門螺桿菌(HP)是人類最常見的病原性細菌,是慢性胃炎、消化性潰瘍等疾病發生的主要病因,并與胃腺癌、胃粘膜相關淋巴組織淋巴瘤的發生密切相關,WHO已將其列為一類致癌因子。HP對胃粘膜造成損害的前提是必須黏附于胃粘膜上皮細胞,進一步引起上皮細胞的某些行為改變,如分泌炎性細胞趨化因子,激活中性粒細胞等,同時胃上皮細胞還可作為抗原呈遞細胞引發T淋巴細胞的活化,最終導致中性粒細胞和淋巴細胞在胃粘膜內浸潤,引起胃粘膜炎癥反應[4,5]。HP在人類的感染率已高達50%以上,其感染引起的炎癥反應是其重要的致病機制之一[6]。由于HP為非侵入性細菌,其所引起的炎癥反應是通過第二信使來完成的,關于第二信使目前研究較多的是炎癥細胞趨化因子,如IL-2、IL-6、IL-8及腫瘤壞死因子等,特別是IL-8研究的比較深入,是目前比較肯定的炎癥介質,其誘導的炎細胞聚集是HP引起胃粘膜病變的關鍵因素,并且有研究進一步證明,只有活的HP才能誘發IL-8的分泌,且HP與胃粘膜上皮細胞的黏附是誘發上皮細胞IL-8分泌的先決條件[7]。從以上的研究中我們可以看出,阻斷HP的黏附是治療HP感染的重要環節。
益生菌是通過改善胃腸內菌群的平衡,對宿主起到有益作用的活性微生物,具有抵御外源細菌定植所表現出的定植抗性,即益生菌通過黏附素與粘膜上皮細胞受體黏附,然后占位定植,以阻止病原菌與粘膜受體結合從而抑制病原菌的黏附[8]。黏附是益生菌發揮作用的首要條件,因此,益生菌的定植抗性是衡量其生態效應的重要指標。
嗜酸乳桿菌是人類胃腸道內主要的益生菌,有研究表明,其有明顯的拮抗HP的作用,但其作用機制尚不清楚,可能與嗜酸乳桿菌對胃上皮細胞的高度親和力有關,由于嗜酸乳桿菌的快速生長使得它能分泌大量的乳酸,可以直接對黏附在上皮細胞的HP起抑制作用,而不干擾胃粘膜的粘液層[10]。動物實驗研究發現乳酸桿菌能阻止HP在鼠胃內的定植,這可能與嗜酸乳桿菌阻礙了HP的黏附有關[10]。嗜酸乳桿菌對胃粘膜表面的選擇性黏附被認為是影響其在有胃液存在的環境中定植的一個重要因素,嗜酸乳桿菌產生的乳酸、乙酸、乙酰醋酸等有機酸在消化道粘膜的分布明顯多于消化道內容物的分布,這些黏附在消化道粘膜表面的有機酸,對消化道病菌的黏附也起到一定的屏障作用[13-14]。
本研究以黏附和黏附抑制為切入點,選用胃粘膜上皮細胞為靶細胞,觀察了嗜酸乳桿菌對胃黏膜上皮細胞的粘附情況,結果顯示,本實驗室篩選的嗜酸乳桿菌L6對胃黏膜上皮細胞具有粘附性,隨濃度的增加粘附性逐漸增強,且粘附后不影響細胞的存活率;L6對HP與胃黏膜上皮細胞的粘附起拮抗作用,且在HP與胃黏膜細胞粘附之前應用嗜酸乳桿菌的拮抗作用更明顯;嗜酸乳桿菌L6還可抑制HP誘導胃上皮細胞分泌Il-8,而其本身并不引起IL-8的分泌,為嗜酸乳桿菌L6在微生態制劑開發中的價值提供依據。
[1]Sakai T,Ognra Y,Narita J,et al.Simultaneous early adenocarcinoma and mucosa-associated lymphoid tissue(MALT)lymphoma of the stomach associated with Helicobacter pylori infection[J].Gastric Cancer,2003,8:191.
[2]Brzozowski T,Konturek PC,Sliwowski Z,et al.Interaction of nonsteroidal anti-inflammatory drugs(NSAID)with Helicobacter pylori in the stomach of humans and experimental animals[J].J Physiol pharmacol,2006,57 (Supple 3):67.
[3]王錦鴻,王江濱,李 巖,等.抗HP嗜酸乳桿菌的篩選及其相關生物學特性的研究[J].中國免疫學雜志,2007,(4):1116.
[4]Pillinqer MH,Marianovic N,Kim SY,et al.Helicobacter pylori stimulates gastric epithelial cell MMP-1 secretion via Cag A-dependent and independent ERK activation[J].J Biol Chem,2007,282(26):18722.
[5]Couturier MR,Tasca E,Momtecuccoc C,et al.Interaction with Cag F is required for translocation of Cag A into the host via the Helicobacter pylori type IV secretion system[J].Infect Immun,2006,74(1):273.
[6]Sunir AE,et al.Typing of Campylobacter pylori by bacterial DNA Brestriction endonuclease analysis and determination of plasmid profile[J].J Clin Microbiol,1990,28:83.
[7]Ding S Z,Cho C H,Lam S K.Helicobacter pylori induces interleukin-8 expression in endothelial cells and the signal path way is protein tyrosine kinase dependent[J].Biochem Biophys Res Commun,1997,240(3):561.
[8]郭興華.益生菌基礎與臨床[M].北京:科學技術出版社.2002:36.
[9]Ryan KA,Daly P,Li Y,er al.Strain-specific inhibition of Helicobacter pylori by Lactobacillus salivarius and other lactobacilli[J].J Antimicrob Chemother,2008,61(4):831.
[10]Sqouras DN,Panayotopoulou EG,Martinez-Gonzalez B,et al.Lactobacillus johnsonii La1 attenuates Helicobacter pylori-associated gastritis and reduces levels of proinflammatory chemokines in C57BL/6 mice[J].Clin Diagn Lab Immunoll,2005,12(12):1378.
[11]Collado MC,Jalonen L,Meriluoto J,et al.Protection mechanism of probiotic combination against human pathogens:in vitro adhesion to human intestinal mucus[J].Asia Pac J Clin Nutr,2006,15(4):570.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
