中國新疆紫花苜蓿復合體3個種的遺傳多樣性及親緣關系研究
2012-06-08 08:22:24李飛飛崔大方羊海軍鄧超宏李慶艷
草業學報 2012年1期
李飛飛,崔大方,羊海軍,鄧超宏,李慶艷
(1.華南農業大學生命科學學院,廣東 廣州510642;2.華南農業大學林學院,廣東 廣州510642;3.華南農業大學公共基礎實驗教學中心,廣東 廣州510642)
苜蓿屬(Medicago)植物是豆科(Leguminosae)一個重要的類群,大多數苜蓿屬物種都是廣泛分布在溫帶草原上的優質牧草。而紫花苜蓿(M.sativa)、黃花苜蓿(M.falcata)以及多變苜蓿(M.varia)更是畜牧業主要的飼料來源[1-3]。
紫花苜蓿復合體包括9個可以自然雜交的分類群[4],包含有二倍體和四倍體物種。在我國僅分布有3個種,即紫花苜蓿、黃花苜蓿和多變苜蓿。3個種均為苜蓿屬多年生草本植物,紫花苜蓿花冠各色,淡黃,深藍或暗紫色、莢果螺旋2~4圈、中央無孔或近無孔,在全國各地都有栽培或呈半野生狀態;黃花苜蓿花冠黃色、莢果鐮形,分布于我國東北、華北和西北地區以及中亞及歐洲地區;多變苜蓿是紫花苜蓿與黃花苜蓿自然雜交產生的類型,其花冠淺紫色、黃色或其他、莢果螺旋0.5~2.0圈、中央有孔,在我國新疆和高加索、中亞細亞、西伯利亞一帶多有分布[5]。
由于這3個種形態相似,并且長期以來在不同的地理條件下雜交,子代和親本類型多樣,使其分類問題存在很大爭議。前蘇聯學者在20世紀初到50年代把它們分成許多種,而中國、美國、加拿大學者則支持將它們定為一個種即多變苜蓿。在Small和Jomphe[6]系統中將多變苜蓿以及Lesins和Lesins[7]系統中的紫花苜蓿、黃花苜蓿劃分為紫花苜蓿種下的3個亞種,分別是M.sativasubsp.varia、M.sativasubsp.sativa及M.sativasubsp.falcata。我國學者何詠松和吳仁潤[8]通過形態學特點和遺傳現象的觀察認為紫花苜蓿和黃花苜蓿屬同1個種的2個亞種或生物型或生態型。李擁軍和蘇加楷[9,10]采用種子貯藏蛋白技術及RAPD(隨機擴增DNA多態性,random amplified polymorphic DNA)標記分析發現新疆大葉苜蓿、新牧1號多變苜蓿和新疆黃花苜蓿之間的遺傳距離為0,支持將這3個苜蓿類型劃分為紫花苜蓿的3個亞種。而王俊杰[11]認為莢果形態在苜蓿屬植物“種”及以上分類地位的界定上是十分有效的,而在種下分類單位的界定上則難以發揮作用,因此不支持Small和Jomphe[6]系統中的劃分,并且將中國黃花苜蓿野生種群的莢果劃分為13個類型。因此,紫花苜蓿、黃花苜蓿及多變苜蓿這3個種及其種下類型的分類地位需進一步研究確定。
近十余年來,許多分子標記包括簡單串聯重復序列SSR(簡單重復序列,simple sequence repeats)[12-14],RAPD[15,16],RFLP(限制性片段長度多態性,restriction fragment length polymorphism)[17]以及細胞器基因片段[18,19]都曾對苜蓿屬植物進行過研究。但這些研究中絕大部分以栽培的紫花苜蓿為主要研究材料,對野生分布的多變苜蓿及黃花苜蓿研究較少,而將共顯性與顯性分子標記結合對廣泛分布于新疆地區的紫花苜蓿復合體3個種的研究還未有報道。因此本研究利用SSR及ISSR(簡單重復序列區間,inter-simple sequence repeat)分子標記技術,比較分析在中國新疆分布的紫花苜蓿、黃花苜蓿和多變苜蓿3個種共10個居群的遺傳多樣性及其親緣關系,為苜蓿屬植物的分類學問題以及苜蓿屬植物資源的合理開發利用和保護提供分子依據。
1 材料與方法
1.1 實驗材料及引物
實驗材料于2007-2008年在野外采集,共有3個種10個居群,每個居群選取19份單株,共采集190份單株,記錄編號后分別放入裝有變色硅膠的封口袋中干燥。居群編號、采集地及采集地經緯度見表1。40對SSR引物來源于Bernadette等[20]發表的紫花苜蓿SSR引物,37個ISSR引物來源于UBC大學公布的100個通用引物。

表1 樣品采集地信息Table 1 Sample list of location
1.2 實驗方法
于2008年將野外采集的苜蓿屬植物每份單株摘取適量葉片,采用改良CTAB(hexadecyltrimethy-ammonium bromide)法提取植物總DNA,并用1.0%的瓊脂糖電泳檢測DNA的濃度和純度。
SSR反應體系:總體積20μL,包括10×Buffer(含 Mg2-)2μL,10mmol/L dNTP 0.3μL,10μmol/L引物各1μL,Taq酶(5U/μL)0.2μL,30ng/μL DNA 模板2μL,去離子水13.5μL。SSR反應程序:94℃變性5 min;94℃變性30s,54℃退火1min,72℃延伸1min,35個循環;72℃延伸10min;4℃保存。ISSR反應體系:總體積20μL,包括10×Buffer(含 Mg2-)2μL,10mmol/L dNTP 0.25μL,10μmol/L 引物1μL,Taq酶(5 U/μL)0.2μL,30ng/μL DNA模板2μL,去離子水14.55μL。ISSR反應程序:94℃變性5min;94℃變性30s,54℃退火45s,72℃延伸1.5min,42個循環;72℃延伸7min;4℃保存。
SSR擴增產物采用6%的聚丙烯酰胺凝膠電泳檢測,ISSR擴增產物采用2%瓊脂糖凝膠電泳檢測。電泳結果至凝膠成像儀拍照觀察,并記錄結果。
1.3 數據統計分析
由于紫花苜蓿和多變苜蓿為同源四倍體,而黃花苜蓿既含有四倍體又含有二倍體[21],所以二倍體等位基因的頻率統計公式并不適合本研究,劉志鵬等[22]曾采用一種將無效等位基因包含在內的統計方法統計四倍體,但由于這種方法還未完善,部分統計指數還需進一步研究。因此本實驗統一采用0/1法統計,以擴增條帶在相對遷移位置有無,賦以“1”或“0”,生成分子數據矩陣。利用PopGene 1.32軟件[23]進行電泳譜帶差異和各項遺傳指數的統計分析,包括:多態位點百分率(PPB)、有效等位基因數(ne)、Nei’s基因多樣性指數(h)、遺傳分化系數(Gst)、基因流(Nm)、Shannon信息指數(I)、遺傳距離(GD)。利用 GeneALEx軟件[24]AMOVA 計算方差分量,及Mental檢測遺傳距離和地理距離相關系數(r)。SSR/ISSR引物多態信息含量采用計算公式:
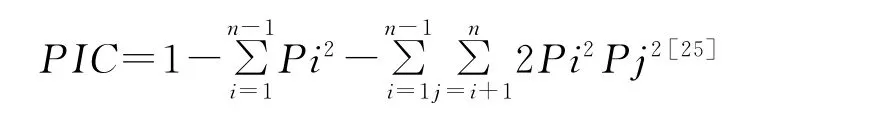
式中,Pi為微衛星位點上第i個等位基因頻率(allelic frequency),Pj為第i+1個等位基因頻率。利用POPTREE2[26]軟件采用非加權平均聚類法(UPGMA)繪制聚類圖,并重復10 000次計算自展支持率[27]。
2 結果與分析
2.1 SSR及ISSR位點多態性
40對SSR引物中篩選出15對多態性好、具清晰條帶的SSR引物用于進一步研究,在10個居群190份材料中共獲得66個位點,64個多態性位點,位點數范圍為2(MTIC258)~11(MTIC248),平均位點數達到4.4個,平均多態性位點數達4.3個,多態位點百分比達到96.97%;10個ISSR引物共獲得74個位點,多態位點百分比達到100%,位點數范圍為2(891)~13(822),平均多態性位點數達7.4個(表2)。

表2 15對SSR引物及10個ISSR引物在總群體中的遺傳多樣性信息Table 2 Genetic diversity informations of 15pairs of SSR primers and 10ISSR primers in total group
多態信息含量是衡量微衛星位點多態性高低的較好的指標,當PIC>0.50時,具有高度多態性;當0.25<PIC<0.50時,具有中度多態性;當PIC<0.25時,具有低度多態性。SSR標記中15個位點中9個位點具有高度多態性,6個位點具有中度多態性,其中MTIC248引物PIC值最高(0.84),MTIC258引物PIC值最低(0.34),平均值為0.54。ISSR標記中10個位點中6個位點具有高度多態性,4個位點具有中度多態性,其中881引物PIC值最高(0.81),891引物最低(0.36),平均值為0.65。
2.2 紫花苜蓿復合體10個居群遺傳多樣性分析
SSR標記下10個居群的遺傳變異顯示(表3),不同居群內的多態位點百分率不同,范圍為57.58% (FE)~75.76% (VG),平均值為68.79%,總群體為96.97%;各居群Shannon信息指數為0.24(FE)~0.33(VG),平均值為0.30,總群體為0.35;各居群 Nei’s基因多樣性指數為0.16(FE)~0.21(VG),平均值為0.19,10個群體基本接近,變異幅度不大,總群體為0.22。10個居群中多變苜蓿居群VG以上3個指數均最高,可見在SSR分子標記反映的結果中多變苜蓿居群VG遺傳多樣性最豐富,黃花苜蓿居群FE的3個指數均最低;多變苜蓿5個居群中3個指數最高的為居群VG,最低的為居群VQ,黃花苜蓿4個居群中3個指數最高的為居群FB,最低的為居群FE;多變苜蓿總群體多態位點百分率(92.42%)、Shannon信息指數(0.35)、Nei’s基因多樣性指數(0.22)均大于黃花苜蓿總群體指數(PPB=86.36%、I=0.32、h=0.20);紫花苜蓿復合體總群體基因分化系數為0.12,基因流為3.62,多變苜蓿居群間基因流 (7.75)大于黃花苜蓿居群間基因流(5.21)。
ISSR標記下10個居群的多態位點百分率為47.30% (SH)~75.68% (FH),平均值為66.49%,總群體為100%;各居群Shannon信息指數為0.19(SH)~0.30(FH),平均值為0.26,總群體為0.33;各居群 Nei’s基因多樣性指數為0.12(SH)~0.19(FH),平均值為0.17,10個群體非常接近,總群體為0.20。與SSR分析結果不同的是10個居群中黃花苜蓿居群FH以上3個指數均最高,紫花苜蓿3個指數均最低;多變苜蓿5個居群中居群VB以上3個指數最高,居群VG最低,黃花苜蓿4個居群中居群FH以上3個指數最高,居群FE最低;多變苜蓿總群體多態位點百分率 (93.24%)大于黃花苜蓿總群體(91.89%),但多變苜蓿總群體Shannon指數(0.30)和 Nei’s基因多樣性指數 (0.18)略小于黃花苜蓿(I=0.32,h=0.20);總群體基因分化系數為0.18,基因流為2.34,基因流低于SSR標記的分析結果,但多變苜蓿居群間基因流(5.43)同樣高于黃花苜蓿(4.94)。黃花苜蓿居群FH在2種分子標記下都存在特有位點,但在ISSR標記下紫花苜蓿存在1個特有位點。
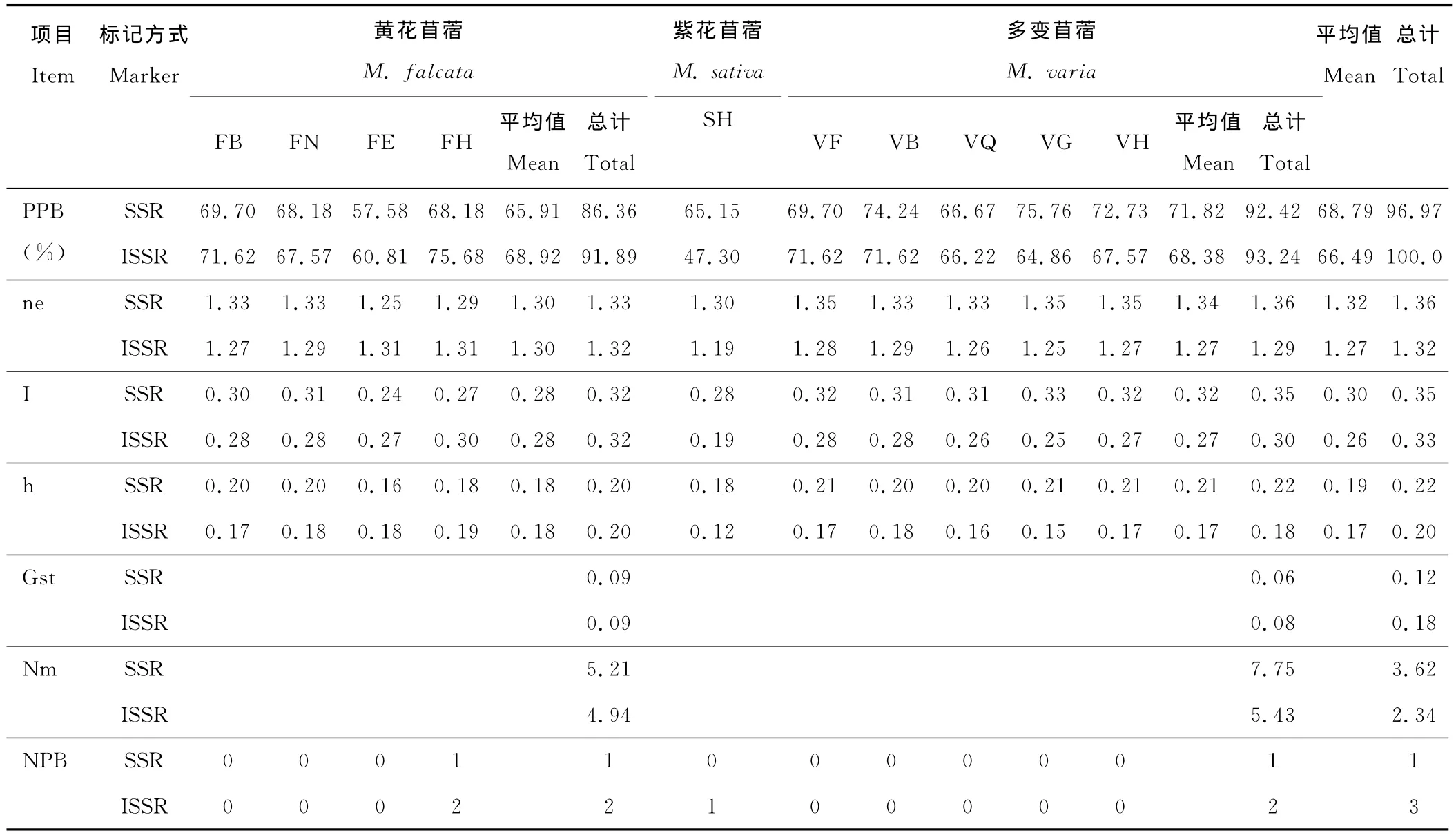
表3 紫花苜蓿復合體10個居群遺傳變異比較Table 3 Genetic variation of 10populations of M.sativacomplex
2.3 紫花苜蓿復合體親緣關系分析
在2種分子標記下總群體Shannon指數 (0.35,0.33)以及Nei’s基因多樣性指數 (0.22,0.20)均高于10個居群的平均值(I=0.30,h=0.19;I=0.26、h=0.17),可反映出遺傳多樣性主要來源于居群內而不是居群間,群體的AMOVA分子方差分析得出SSR標記下遺傳變異的7%來源于種間,7%來源于種內居群間,86%來源于居群內,ISSR標記下遺傳變異的12%來源于種間,遺傳變異的10%來源于種內居群間,79%來源于居群內,2種分子標記均證明遺傳變異主要分布在居群內。但2種標記對種間、居群間、居群內差異的劃分并不十分明顯(表4)。

表4 紫花苜蓿復合體10個居群SSR變異的分子方差分析Table 4 Analysis of molecular variance(AMOVA)for 190individual plants using 66SSR alleles and 74ISSR bands respectively
SSR標記中Nei’s遺傳距離平均值為0.07,ISSR標記中Nei’s遺傳距離平均值為0.19。利用Mantel檢測,2種分子標記下10個居群的遺傳距離相關系數(r)為0.50,P<0.001,呈顯著相關。SSR與ISSR兩種分子標記的UPGMA聚類分析表明(圖1):3個種中,黃花苜蓿與多變苜蓿關系較近,紫花苜蓿與其他2種遺傳距離較遠,單獨形成一支,自展支持率為100%;紫花苜蓿的所有居群在2種標記下均聚為一類;而黃花苜蓿的FE居群在SSR標記下單獨成為一支,且自展支持率為65%。

圖1 基于SSR(A)及ISSR(B)標記下紫花苜蓿復合體3個種10個居群Nei’s遺傳距離構建的UPGMA樹狀圖Fig.1 UPGMA dendrograms of the genetic relationships among 10populations of M.sativacomplex constructed from estimated simple matching genetic distance based on SSR markers(A)and ISSR markers(B)
Sun等[28]認為不同標記體系的數據綜合起來進行整體分析,更能客觀的反映種間關系。因此,本研究將ISSR和SSR數據合并起來,計算紫花苜蓿復合體10個居群的遺傳距離,并構建UPGMA樹。結果顯示遺傳距離范圍為0.05(VF和 VB)~0.23(VQ和SH),平均遺傳距離為0.12,大于SSR標記下的平均遺傳距離,小于ISSR標記下的平均遺傳距離。基于2種分子標記構建的UPGMA樹(圖2)顯示,多變苜蓿的所有居群聚為一支,自展支持率為86%。在多變苜蓿的5個居群中進一步劃分為2個亞分支,一支包含VQ、VG和VH居群,另一支包含VF和VB居群。黃花苜蓿的3個居群聚為一支,自展支持率為79%,FE居群從其他居群中分離出來。Metal相關性分析證明,多變苜蓿5個居群和黃花苜蓿4個居群的遺傳距離與其相應的地理距離(表5)不相關,相關系數(r)分別為0.49,P>0.05;-0.32,P>0.1。

圖2 基于全部分子數據的紫花苜蓿復合體3個種10個居群的Nei’s遺傳距離UPGMA樹狀圖Fig.2 UPGMA dendrogram of the genetic relationships among 10 populations of 3species constructed from estimated simple matching genetic distance based on SSR and ISSR combined data

表5 黃花苜蓿居群間及多變苜蓿居群間的地理距離Table 5 Geographical distances among populations of M.falcataand M.varia km
3 討論
3.1 紫花苜蓿復合體3個種及種下居群遺傳多樣性分析
本研究首次以SSR和ISSR標記對紫花苜蓿復合體居群內及居群間的遺傳分化模式進行了探討,結果發現,2種分子標記的平均多態信息含量都達到了高度多態性,且SSR及ISSR位點在居群內及居群間均表現為豐富的遺傳變異,表明居群內及居群間均存在豐富的遺傳多樣性。此外SSR等位變異與ISSR位點在居群間和居群內的變異程度極不平衡,不同位點間的變異相差較大。結果還發現SSR標記中僅FH具有特有變異位點,ISSR標記中僅居群FH和VH具有特有變異位點,這2個居群的采集地都位于新疆烏魯木齊后峽距白楊溝10km處,因此推測這個采集地的黃花苜蓿與多變苜蓿具有較豐富的遺傳多樣性,保存價值大。
在SSR分子標記下的群體遺傳多樣性分析表明,黃花苜蓿居群FE的多態位點百分率、Shannon信息指數、Nei’s基因多樣性指數、PIC指數均最低,并具有最多純合位點,與其他居群差別較大,分析主要有以下2個原因,1)黃花苜蓿居群FE的樣品形態特征為葉片小、黃色花、莢果鐮形,且生境偏旱,為黃花苜蓿的變種草原苜蓿(M.falcatavar.romanica)。2)采自額敏喇嘛昭山前荒漠草原,地理分布上處于相對封閉的狀態,人口牲畜來往不頻繁,不會帶來太多的雜交機會。
10個居群相比較的結果表明,SSR標記分析中居群VG遺傳多樣性最高,FE遺傳多樣性最低,ISSR標記分析中居群FH遺傳多樣性最高,紫花苜蓿遺傳多樣性最低。因此不同的分子標記在分析遺傳多樣性時所得到的結果有所不同,進一步分析發現VG位于新疆烏魯木齊甘溝草原站的保護區內,分布區內的植被得到了當地很好的保護,而FH具有特有變異位點,因此這2個居群在不同分子標記中表現出最豐富的遺傳多樣性具有其合理性。而紫花苜蓿在ISSR標記中表現出較低的遺傳多樣性,推測與其為栽培品種有關。多變苜蓿與黃花苜蓿總體遺傳多樣性相差不大,在2種分子標記中高低各有不同,但多變苜蓿多態性比率在2種分子標記中均高于黃花苜蓿,說明多變苜蓿遺傳變異略大于黃花苜蓿。
通過種間-種內、居群間-居群內各層次結構的AMOVA分析發現,2種標記均顯示群體遺傳變異主要來源于居群內。Hilde和Lgor[29]認為,一年生或短命多年生、自交和演替階段早期的類群在居群間保持有高的遺傳變異,而壽命長、異交、演替階段晚期的類群在居群內保持有高的遺傳變異。而紫花苜蓿復合體3個種均為多年生、異交、演替階段晚期植物,因此在居群內具有高的遺傳變異與其本身生物學特性有關。
多變苜蓿5個居群之間的基因流(SSR Nm=7.752,ISSR Nm=5.431)大于黃花苜蓿4個居群之間的基因流(SSR Nm=5.214,ISSR Nm=4.939);SSR標記中總群體基因流為3.619,ISSR標記分析中總群體基因流為2.336。本研究結果中的基因流均大于一般廣布種植物的基因流(Nm=1.881)[30],且 Wright[31]認為,當 Nm值大于1時基因流就可以防止由遺傳漂變引起的居群之間的分化,因此紫花苜蓿復合體種群中的基因流防止了3個種居群之間的分化,并防止了同源四倍體在進化過程上變得呆板。
雖然基因流對防止物種分化具有很重要的作用,但另一方面,在人類活動干擾或自然環境變化過程中,種間基因流和雜交可能導致原生境隔離的近緣物種間產生雜交沒化(hybridization merging),使得物種間的區別不再像以前明顯進而導致物種合并[32,33],形成不同分類群間遺傳同化(genetic assimilation)現象。因此介于紫花苜蓿復合體3個種之間具有的雜交現象,其種群的基因流會隨時間而改變,需定時對其進行測定,以了解當前這3個種的遺傳分化。
3.2 紫花苜蓿復合體3個種的親緣關系分析
從聚類圖上分析,SSR與ISSR數據合并構成的聚類圖不僅符合2個標記共有的特征,且所反映三者之間的關系更為客觀。UPGMA樹上形成3個主要的分支,紫花苜蓿獨立成一支,來源于同一采集地的黃花苜蓿居群FH與多變苜蓿居群VH都已分開,充分說明多變苜蓿、黃花苜蓿和紫花苜蓿雖然遺傳關系較近,但三者之間仍可以得到有效地劃分,因此盡管這3個種之間的遺傳關系非常近,但本研究仍支持將多變苜蓿、黃花苜蓿和紫花苜蓿劃分為3個種。很長一段時間,黃花苜蓿是否應當劃分為一個單獨的種都存在爭議。Brummer等[34]采用RFLP分析二倍體和四倍體苜蓿發現黃花苜蓿與M.caerulea不同,能有效的區分。Havananda等[35]基于葉綠體和線粒體基因同樣也討論了黃花苜蓿與M.caerulea的關系,葉綠體的結果顯示黃花苜蓿明顯的與較原始的種M.caerulea分別開來,而線粒體的結果卻不明顯。因四倍體的紫花苜蓿是由M.caerulea直接進化而來,故本研究中基于SSR和ISSR兩種分子標記分析的結果與Havananda等[35]的研究結果相似。而孫毅等[36]利用核基因ITS的研究結果顯示黃花苜蓿與紫花苜蓿應該合并為一個種。因此,這些研究結果并不能決定這3個分類群為種或亞種,但可以確定的是他們的DNA序列很不相同,利用不同的基因片段分析得出的結果會不一樣。從地理分布和物種特性上來看,完全野生的紫花苜蓿沒有記載,而黃花苜蓿多為野生分布。因此本研究支持將這3個分類群作為3個種的劃分。遺傳關系顯示多變苜蓿與黃花苜蓿關系較近,這與李擁軍和蘇加楷[10]利用RAPD分子標記所得到的結果有所不同,他認為多變苜蓿與紫花苜蓿遺傳距離更小。
多變苜蓿、黃花苜蓿各居群的遺傳距離與其地理距離相關性分析發現沒有顯著相關性存在于多變苜蓿及黃花苜蓿的遺傳距離與地理距離之間。這主要由于紫花苜蓿各居群的生長環境不一致導致,而黃花苜蓿居群FE主要為草原苜蓿與其他居群遺傳分化較大,明顯區分于其他居群分支而單獨存在。因此本研究的分子標記所得到的結果支持Lesins和Lesins[7]的結論,即草原苜蓿為黃花苜蓿一個變種的分類學地位。
通過以上研究,得出以下3個結論:1)多變苜蓿居群VG和黃花苜蓿居群FH在10個居群中遺傳多樣性最高,并且居群FH及居群SH含有特有多態性位點,因此需要加強保護。2)本研究結果支持仍將紫花苜蓿復合體3個分類群劃分為3個種。3)SSR和ISSR標記作為研究紫花苜蓿復合體的遺傳多樣性及遺傳關系是非常有效的分子標記。
致謝:感謝牧草植物分類學專家崔乃然教授的指導,感謝新疆畜牧科學院李捷老師提供的幫助。
[1]秦峰梅,張紅香,武祎,等.鹽脅迫對黃花苜蓿發芽及幼苗生長的影響[J].草業學報,2010,19(4):71-78.
[2]鄒亞麗,韓方虎,耿麗英,等.溫度和濕度對紫花苜蓿土壤氮礦化的影響[J].草業學報,2010,19(4):101-107.
[3]王平,周道瑋,姜世成.半干旱地區禾-豆混播草地生物固氮作用研究[J].草業學報,2010,19(6):276-280.
[4]Gunn C R,Skrdla W H,Spencer H C.Classification ofMedicagosativaL.using Legume Characters and Flower Colors[M].Washington D C:United States Department of Agriculture Press,1978:84.
[5]中國科學院中國植物志編輯委員會.中國植物志(第42卷第2分冊)[M].北京:科學出版社,1998:304-328.
[6]Small E,Jomphe M.A synopsis of the genusMedicago(Leguminosae)[J].Canadian Journal of Botany,1988,67:3260-3294.
[7]Lesins K A,Lesins I.GenusMedicago(Leguminosae):A Taxogenetic study[M].The Hague:W.Junk Press,1979:66-214.
[8]何詠松,吳仁潤.苜蓿自交不親和性的研究[J].草業科學,1987,4(4):6-11.
[9]李擁軍,蘇加楷.中國苜蓿地方品種親緣關系的研究Ⅰ種子貯藏蛋白標記[J].草業學報,1999,8(1):31-41.
[10]李擁軍,蘇加楷.中國苜蓿地方品種親緣關系的研究ⅡRAPD標記[J].草業學報,1999,8(3):46-53.
[11]王俊杰.中國黃花苜蓿野生種質資源研究[D].內蒙古:內蒙古農業大學,2008.
[12]Diwan N,Bhagwat A A,Bauchan G R,etal.Simple sequence repeat DNA markers in alfalfa and perennial and annualMedicagospecies[J].Genome,1997,40(6):887-895.
[13]Mengoni A,Ruggini C,Vendramin G G,etal.Chloroplast microsatellite variations in tetraploid alfalfa[J].Plant Breeding,2000,119(6):509-512.
[14]Falahati-Anbaran M,Habashi A A,Esfahany M,etal.Population genetic structure based on SSR markers in alfalfa(MedicagosativaL.)from various regions contiguous to the centres of origin of the species[J].Journal of Genetics,2007,86(1):59-63.
[15]Crochemore M L,Huyghe C,Kerlan M C,etal.Partitioning and distribution of RAPD variation in a set of populations of theMedicagosativacomplex[J].Agronomie,1996,16:421-432.
[16]Ghérardi M,Mangin B,Goffinet B,etal.A method to measure genetic distance between allogamous populations of alfalfa(Medicagosativa)using RAPD molecular markers[J].Theoretical and Applied Genetics,1998,96:406-412.
[17]Kidwell K K,Bingham E T,Woodfield D R,etal.Relationships among genetic distance,forage yield and heterozygosity in isogenic diploid and tetraploid alfalfa populations[J].Theoretical and Applied Genetics,1994,89:323-328.
[18]Skinner D Z.Non random chloroplast DNA hyper variability inMedicagosativa[J].Theoretical and Applied Genetics,2000,101:1242-1249.
[19]Havananda T,Brummer E C,Maureira-Butler I J,etal.Relationships among diploid members of theMedicagosativa(Fabaceae)species complex based on chloroplast and mitochondrial DNA sequences[J].Systematic Botany,2010,35(1):140-150.
[20]Bernadette J,Sandrine F,Philippe B,etal.Construction of two genetic linkage maps in cultivated tetraploid alfalfa(Medicagosativa)using microsatellite and AFLP markers[J].Plant Biology,2003,3:9.
[21]Qurios C F,Bauchan G R.Alfalfa and Alfalfa Improvement:The GenusMedicagoand the Origin of theMedicagosativaComplex[M].Madison,Wisconsin:American Society of Agronomy,Crop Science Society of America,Soil Science Society of America Press,1988:93-124.
[22]劉志鵬,楊青川,呼天明,等.用SSR標記研究不同耐鹽特性四倍體紫花苜蓿的遺傳多樣性[J].作物學報,2006,32(4):630-632.
[23]Yeh F C,Yang R-C,Boyle T B J,etal.Popgene:Microsoft Windows?-Based Freeware for Population Genetic Analysis,Version 1.32[M].Canada:University of Alberta and Centre for International Forestry Research,1997.
[24]Peskall R,Smouse P E.GENEALEX 6:Genetic analysis in Excel.Population genetic software for teaching and research[J].Molecular Ecology Notes,2006,6:288-295.
[25]Botstein D,White R L,Skolnick M,etal.Construction of genetic linkage map in man using restriction fragment length polymorphisms[J].American Journal of human genetics,1980,32:314-331.
[26]Takezaki N,Nei M,Tamura K.POPTREE2:Software for constructing population trees from allele frequency data and computing other population statistics with windows interface[J].Molecular Biology and Evolution,2010,27(4):747-752.
[27]Felsenstein J.Confidence limits on phylogenies:an approach using the bootstrap[J].Evolution,1985,39(4):783-791.
[28]Sun G L,Díaz O,Salomon B,etal.Genetic diversity inElymuscaninusas revealed by isozyme,RAPD,and microsatellite markers[J].Genome,1999,42:420-431.
[29]Hilde N,Lgor V B.Effective of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants[J].Perspectives in Plant Ecology,Evolution and Systematic,2000,3(2):93-114.
[30]Hamrick J L.Gene Flow and Distribution of Genetic Variation in Plant Populations[M].London:New York Academes Press,1987:53-68.
[31]Wright S.Evolution in Mendelian population[J].Anatomical Record-Advances in Integrative Anatomy and Evolutionary Biology,1929,44:287.
[32]Rieseberg L H,Carney S E.Tansley review no.102plant hybridization[J].New Phytologist,1998,140:599-624.
[33]Gross B L,Kane N C,Lexer C,etal.Reconstructing the origin ofHelianthusdeserticola:survival and selection on the desert floor[J].The American Naturalist,2004,164:145-156.
[34]Brummer E C,Kochert G,Bouton J H.RFLP variation in diploid and tetraploid alfalfa[J].Theoretical and Applied Genetics,1991,83:89-96.
[35]Havananda T,Brummer E C,Maureira-Butler I J,etal.Relationships among diploid members of theMedicagosativa(Fabaceae)species complex based on chloroplast and mitochondrial DNA sequences[J].Systematic Botany,2010,11:140-150.
[36]孫毅,梁愛華,王景雪,等.根據核糖體DNA ITS序列分析苜蓿屬的系統分類[J].西北植物學報,2003,23(2):242-246.
