高滲應(yīng)激環(huán)境下傷寒沙門菌t4606基因功能研究*
2014-10-11 01:57:24繆敏慧孟紅委倪紅元戴如順蘇州大學附屬第二醫(yī)院檢驗科江蘇蘇州215004
檢驗醫(yī)學與臨床 2014年18期
繆敏慧,孟紅委,倪紅元,戴如順,王 敏,杜 鴻(蘇州大學附屬第二醫(yī)院檢驗科,江蘇蘇州 215004)
傷寒沙門菌是沙門菌屬中經(jīng)消化道感染人類的致病菌之一,可導(dǎo)致感染患者發(fā)生全身系統(tǒng)性感染,并能引發(fā)腸穿孔等嚴重的并發(fā)癥,進而危及感染患者的生命[1-2]。傷寒沙門菌作為食源性致病菌,必須克服人體腸道內(nèi)的高滲應(yīng)激環(huán)境才能最終致病。筆者在前期的研究工作中發(fā)現(xiàn),在高滲應(yīng)激環(huán)境下,傷寒沙門菌t4606基因的表達受到兩個重要的sigma因子,即RpoE和RpoS的雙重調(diào)控[3],提示t4606基因?qū)抽T菌克服高滲應(yīng)激環(huán)境極為重要。本研究利用原核生物基因敲除技術(shù)制備了傷寒沙門菌t4606基因缺陷株,并分析了其在高滲應(yīng)激環(huán)境下的生物學特性,旨在通過分析t4606基因的功能,及其在沙門菌致病機制中的作用,為臨床治療和預(yù)防傷寒沙門菌感染提供可能的靶向基因。
1 材料與方法
1.1 菌株和質(zhì)粒 本試驗所采用的傷寒沙門菌、自殺質(zhì)粒及大腸埃希菌(E.coli)DH5α菌株由本實驗室保存。
1.2 試劑 核酸限制性內(nèi)切酶BglⅡ、BamHⅠ及用于聚合酶鏈反應(yīng)(PCR)的EXTaq酶購自寶生物工程(大連)有限公司;質(zhì)粒提取試劑、膠回收試劑及T4DNA連接酶購自美國Promega公司。
1.3 方法
1.3.1 t4606基因缺陷株的制備 利用原核生物基因同源重組技術(shù)制備t4606缺陷變異株,篩選連續(xù)3次傳代均獲得穩(wěn)定重組的菌株作為t4606基因缺陷變異株,并采用序列分析進行驗證。具體方法參考文獻[4]。相關(guān)引物見表1。
1.3.2 t4606基因缺陷株回補株的制備 設(shè)計特異性引物擴增t4606基因全長,上、下游引物的5′端分別加入NcoⅠ、SalⅠ酶切位點(見表1),擴增獲得的全長片段經(jīng)酶切后與表達載體pBAD/gⅢ(阿拉伯糖操縱元)連接,克隆至E.coli DH5α感受態(tài)細胞,利用質(zhì)粒的氨芐西林抗性初步篩選陽性重組質(zhì)粒,篩選獲得的陽性重組質(zhì)粒采用PCR及序列分析進行驗證(由上海生工生物技術(shù)公司完成)。培養(yǎng)經(jīng)驗證確定的陽性重組菌株,提取重組質(zhì)粒pBADt4606,電擊導(dǎo)入t4606變異株,經(jīng)酶切及PCR驗證后命名為回補菌Δt4606(pBADt4606)。采用質(zhì)量體積比(W/V)為0.2%的阿拉伯糖誘導(dǎo)質(zhì)粒表達,具體方法參考文獻[5]。
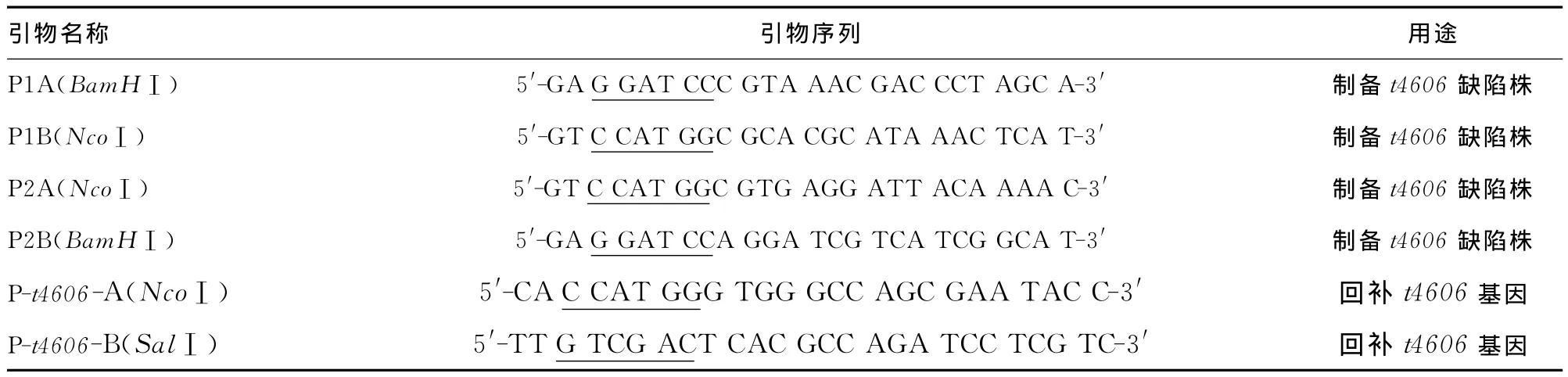
表1 PCR引物及序列
1.3.3 生長曲線分析 基因缺陷株、缺陷回補株和野生株分別在37℃條件下,于LB培養(yǎng)液中振搖培養(yǎng)過夜;過夜培養(yǎng)后以1∶100的比例轉(zhuǎn)入新鮮、預(yù)熱的LB培養(yǎng)液,每隔1h檢測培養(yǎng)液在600nm處的吸光度值(A600值),以A600值為縱坐標,培養(yǎng)時間為橫坐標繪制生長曲線。用相同方法比較t4606缺陷株和野生株在高滲應(yīng)激(300mmol/L NaCl)環(huán)境下的生存能力。每組實驗重復(fù)3次。于對數(shù)生長早期(培養(yǎng)4h)和對數(shù)生長晚期(12h)比較不同菌株培養(yǎng)液的A600值。
1.4 統(tǒng)計學處理 采用SPSS13.0軟件進行數(shù)據(jù)處理和統(tǒng)計學分析。計量資料組間比較采用t檢驗;P<0.05為比較差異有統(tǒng)計學意義。
2 結(jié) 果
2.1 成功制備t4606基因缺陷株 首先利用自殺質(zhì)粒所具有的氨芐西林抗性,在含有氨芐西林的LB平板上篩選具有抗性的菌落,再進一步篩選耐蔗糖的菌落。以篩選獲得的陽性菌落DNA為模板,用t4606基因上、下游引物P1A和P2B進行PCR擴增,分析傷寒沙門菌t4606基因的重組變異情況。PCR擴增檢測相應(yīng)菌落,直至篩選獲得連續(xù)傳代超過3次均僅有缺損片段的菌株。所獲得的菌株即為穩(wěn)定的t4606基因缺陷株。缺陷株及野生株P(guān)CR產(chǎn)物電泳結(jié)果見圖1。經(jīng)DNA序列分析,進一步證實702bp處被BglⅡ核酸限制性內(nèi)切酶的6個堿基酶切位點所替代。
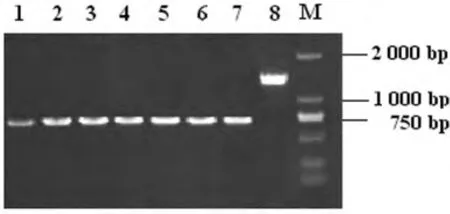
圖1 t4606缺陷株及野生株P(guān)CR產(chǎn)物電泳圖
2.2 成功構(gòu)建回補菌株Δt4606(pBADt4606) 為進一步證實t4606基因的功能,利用表達載體pBAD/gⅢ將t4606基因回補引入t4606基因缺陷株,構(gòu)建回補菌株Δt4606(pBADt4606)。利用質(zhì)粒的氨芐西林抗性,初步篩選攜帶回補t4606基因質(zhì)粒的細菌后,提取質(zhì)粒,進行酶切及PCR驗證,酶切產(chǎn)物電泳結(jié)果見圖2。DNA序列分析證實堿基序列與實驗設(shè)計一致。
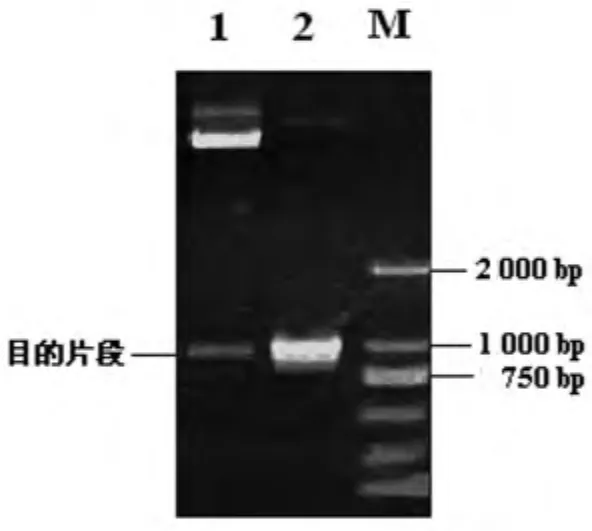
圖2 回補t4606基因的鑒定
2.3 生長曲線分析 于對數(shù)生長早期(培養(yǎng)4h)和對數(shù)生長晚期(12h)檢測并比較不同菌株培養(yǎng)液的A600值,結(jié)果顯示,高滲應(yīng)激環(huán)境下t4606缺陷株(Δt4606)的生存能力明顯弱于野生株(4h:t=22.725,P<0.05;12h:t=5.008,P<0.05),回補t4606基因至t4606缺陷株后,細菌生存能力恢復(fù)至野生株水平(4h:t=3.559,P>0.05;12h:t=0.404,P>0.05),說明t4606基因在細菌適應(yīng)高滲應(yīng)激環(huán)境時發(fā)揮了重要作用。生長曲線如圖3所示。
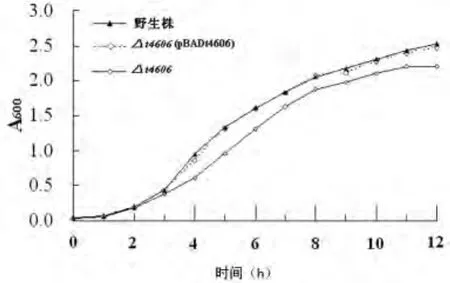
圖3 不同菌株在高滲應(yīng)激環(huán)境下的生存曲線
3 討 論
傷寒沙門菌作為食源性致病菌,常通過污染食物而經(jīng)消化道入人體內(nèi),進而造成全身性感染,嚴重時可危及感染患者的生命。全球每年約有2 100 萬人感染傷寒沙門菌,其中死亡人數(shù)超過21萬,即使在發(fā)達國家,傷寒沙門菌的暴發(fā)流行也時有發(fā)生[6]。傷寒沙門菌在致病過程中,必須克服人體腸道內(nèi)的高滲應(yīng)激環(huán)境才能最終致病[7-8]。在應(yīng)激環(huán)境下,細菌需要及時有效地調(diào)節(jié)某些基因的表達和蛋白的活性,從而適應(yīng)新的環(huán)境。盡管傷寒沙門菌已成為重要的原核生物研究模式菌,其全基因組結(jié)構(gòu)也已闡明,然而仍然有近三分之一基因的功能尚不明確,而這些基因在細菌生存過程中可能具有極為重要的生物學作用。隨著一些未知基因功能的逐步明確,對傷寒沙門菌致病機制的認識也將進一步深入。然而,如何選擇在傷寒沙門菌適應(yīng)應(yīng)激環(huán)境時發(fā)揮重要作用的未知功能基因,是后續(xù)研究工作中的難點。
筆者前期利用傷寒沙門菌全基因組芯片,研究了高滲應(yīng)激環(huán)境下的兩個重要調(diào)節(jié)因子(RpoE和RpoS)的雙重調(diào)控作用,結(jié)果發(fā)現(xiàn),在傷寒沙門菌處于高滲應(yīng)激環(huán)境時,t4606基因的表達同時受到RpoE和RpoS的雙重調(diào)控,說明t4606基因在傷寒沙門菌適應(yīng)高滲應(yīng)激環(huán)境方面具有重要的意義[3]。進一步利用生物信息學軟件對t4606基因序列進行分析后發(fā)現(xiàn),t4606基因具有70%的Patatin樣磷脂酶基序,可能編碼一種Patatin樣磷脂酶。Patatin樣磷脂酶和細菌的侵襲性和毒力密切相關(guān)。Banerji和Flieger[9]的研究表明,具有致病性的細菌內(nèi),含有Patatin樣編碼區(qū)域的蛋白比非致病性細菌更為常見。也有研究表明,銅綠假單胞菌肺炎患者體內(nèi)出現(xiàn)急性肺損傷,有可能是由銅綠假單胞菌Ⅲ型分泌系統(tǒng)分泌的ExoU毒素進入真核細胞的細胞質(zhì)所致[10]。ExoU毒素由exoU基因編碼,而exoU基因類似于傷寒沙門菌t4606基因,其所編碼蛋白質(zhì)的氨基酸序列中具有Patatin樣磷脂酶片段,且該片段與哺乳動物體內(nèi)的非鈣離子依賴性磷脂酶A2具有同源性,并且ExoU毒素的溶血磷酸酶A活性和細胞毒作用與該片段有關(guān)[10]。傷寒沙門菌中是否存在Patatin樣磷脂酶尚無文獻報道,值得進一步深入研究。
本研究采用分子生物學技術(shù),結(jié)合t4606基因的特性,通過優(yōu)化相關(guān)實驗條件,成功制備了傷寒沙門菌t4606基因缺陷株及其回補株,并觀察了不同菌株在模擬人體腸道高滲應(yīng)激環(huán)境下的生存能力。通常情況下,腸道致病菌在進入人體腸道后,必須適應(yīng)環(huán)境滲透壓的巨大改變,即從食物中的低滲環(huán)境到腸道內(nèi)的高滲環(huán)境。致病菌只有適應(yīng)了新的滲透壓環(huán)境,才能侵襲腸上皮細胞。因此,了解傷寒沙門菌適應(yīng)人體腸道內(nèi)高滲應(yīng)激環(huán)境的機制,對進一步分析傷寒沙門菌的整體致病機制具有重要的意義。本研究中的生長曲線分析證實,高滲應(yīng)激環(huán)境下,t4606缺陷株的生長能力明顯弱于野生株,但在回補t4606基因至t4606缺陷株后,其生長能力恢復(fù)至野生株水平。由此可見,t4606基因在傷寒沙門菌適應(yīng)高滲應(yīng)激環(huán)境方面發(fā)揮著重要的作用。
本文對高滲應(yīng)激環(huán)境下傷寒沙門菌t4606基因的生物學功能進行了初步研究,為進一步深入了解t4606基因在傷寒沙門菌中發(fā)揮作用的機制奠定了基礎(chǔ),也說明t4606有可能作為臨床預(yù)防和治療傷寒沙門菌感染的靶向基因,為傷寒沙門菌感染的臨床分子靶向治療提供了新的研究方向。
[1]Ukwenya AY,Ahmed A,Garba ES.Progress in management of typhoid perforation[J].Ann Afr Med,2011,10(4):259-265.
[2]Parry CM,Hien TT,Dougan G,et al.Medical progress:typhoid fever[J].NEJM,2002,347(22):1770-1782.
[3]Du H,Wang M,Luo Z,et al.Coregulation of gene expression by sigma factors RpoE and RpoS in Salmonella enterica serovar Typhi during hyperosmotic stress[J].Cur Microbiol,2011,62(5):1483-1489.
[4]茅凌翔,朱超望,黃新祥,等.傷寒沙門菌phoP基因缺陷變異株的制備[J].江蘇大學學報:醫(yī)學版,2007,17(2):145-149.
[5]Du H,Sheng XM,Zhang HF,et al.RpoE may promote flagellar gene expression in Salmonella enterica serovar Typhi under hyperosmotic stress[J].Curr Microbiol,2011,62(2):492-500.
[6]Bhutta ZA,Threlfau J.Addressing the global disease burden of typhoid fever[J].JAMA,2009,302(8):898-899.
[7]Winter SE,Winter MG,Godinez I,et al.A rapid change in virulence gene expression during the transition from the intestinal lumen into tissue promotes systemic dissemination of Salmonella[J/OL].PLoS Pathogens,2010-08-19[2013-01-07].http://www.ncbi.nlm.nih.gov/pubmed/20808848 .
[8]Zhao L,Ezak T,Li ZY,et al.Vi-Suppressed wild strain Salmonella typhi cultured in high osmolarity is hyperinvasive toward epithelial cells and destructive of Peyer′s patches[J].Microbiol Immunol,2001,45(2):149-158.
[9]Banerji S,F(xiàn)lieger A.Patatin-like proteins:a new family of lipolytic enzymes present in bacteria[J].Microbiol,2004,150(3):522-525.
[10]Pankhaniya RR,Tamura M,Allmond LR,et al.Pseudomonas aeruginosa causes acute lung injury via the catalyticactivity of the patatin-like phospholipase domain of ExoU[J].Crit Care Med,2004,32(11):2293-2299.
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
小讀者(2021年2期)2021-03-29 05:03:48
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
中國生殖健康(2020年6期)2020-02-01 06:28:50
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年11期)2019-01-07 01:28:02
文苑(2018年22期)2018-11-19 02:54:14
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
