山羊繁殖季節(jié)性相關(guān)基因KiSS-1與RFRP的表達(dá)研究
2015-03-23 02:57:35黃冬維儲(chǔ)明星劉秋月胡文萍王翔宇潘章源郭曉飛
畜牧獸醫(yī)學(xué)報(bào) 2015年6期
黃冬維,儲(chǔ)明星,狄 冉,劉秋月,胡文萍,王翔宇,潘章源,郭曉飛
(1.中國(guó)農(nóng)業(yè)科學(xué)院北京畜牧獸醫(yī)研究所,農(nóng)業(yè)部畜禽遺傳資源與種質(zhì)創(chuàng)新重點(diǎn)實(shí)驗(yàn)室,北京 100193;2.安徽省農(nóng)業(yè)科學(xué)院畜牧獸醫(yī)研究所,合肥 230031)
山羊繁殖季節(jié)性相關(guān)基因KiSS-1與RFRP的表達(dá)研究
黃冬維1,2,儲(chǔ)明星1*,狄 冉1,劉秋月1,胡文萍1,王翔宇1,潘章源1,郭曉飛1
(1.中國(guó)農(nóng)業(yè)科學(xué)院北京畜牧獸醫(yī)研究所,農(nóng)業(yè)部畜禽遺傳資源與種質(zhì)創(chuàng)新重點(diǎn)實(shí)驗(yàn)室,北京 100193;2.安徽省農(nóng)業(yè)科學(xué)院畜牧獸醫(yī)研究所,合肥 230031)
本研究為初步闡明KiSS-1和RFRP基因?qū)ι窖蚍敝臣竟?jié)性的作用,選擇常年發(fā)情濟(jì)寧青山羊和季節(jié)性發(fā)情遼寧絨山羊?yàn)閷?duì)比品種,應(yīng)用RT-PCR和qRT-PCR技術(shù)分別研究KiSS-1和RFRP基因組織表達(dá)譜以及在下丘腦-垂體-卵巢(子宮)繁殖軸表達(dá)差異。結(jié)果發(fā)現(xiàn):(1)KiSS-1基因主要表達(dá)于山羊下丘腦、垂體、松果體、腎、卵巢、脂肪和甲狀腺等組織,RFRP基因主要表達(dá)于山羊下丘腦、大腦皮層、小腦、卵巢和垂體等組織,兩基因組織表達(dá)譜均存在一定的品種特異性;(2)KiSS-1基因在濟(jì)寧青山羊下丘腦表達(dá)量顯著高于遼寧絨山羊(P<0.05),而在垂體和卵巢中的表達(dá)量?jī)善贩N間差異不明顯(P>0.05);(3)濟(jì)寧青山羊下丘腦RFRP基因表達(dá)顯著高于遼寧絨山羊(P<0.05),而濟(jì)寧青山羊卵巢和垂體中RFRP基因表達(dá)明顯低于遼寧絨山羊(P<0.05)。研究結(jié)果提示,KiSS-1基因可能參與調(diào)控山羊季節(jié)性繁殖,而RFRP基因是否調(diào)控季節(jié)性繁殖或以何種方式作用還需要進(jìn)一步研究。本研究可為山羊KiSS-1和RFRP基因功能研究打下基礎(chǔ),并為揭示山羊繁殖季節(jié)性的遺傳基礎(chǔ)提供參考。
山羊;繁殖季節(jié)性;KiSS-1;RFRP;表達(dá)
自然界多數(shù)動(dòng)物表現(xiàn)為季節(jié)性繁殖,繁殖季節(jié)性受下丘腦-垂體-性腺軸系統(tǒng)的調(diào)控[1]。雖經(jīng)多年研究,但動(dòng)物繁殖季節(jié)性的分子調(diào)控機(jī)制仍不清楚。近年來(lái),研究者發(fā)現(xiàn),RFamide家族神經(jīng)肽類編碼基因KiSS-1與RFRP可能是動(dòng)物季節(jié)性繁殖活動(dòng)的重要調(diào)控因子。KiSS-1基因最初從人黑色素瘤細(xì)胞中分離出來(lái),它編碼一個(gè)含164個(gè)氨基酸殘基的蛋白,編碼產(chǎn)物經(jīng)過加工后產(chǎn)生13~54個(gè)氨基酸殘基長(zhǎng)度不等的生物活性多肽,這些多肽通稱為kisspeptins,它們具有相同的C端結(jié)構(gòu)和共同受體GPR54[2-3]。Kisspeptin神經(jīng)纖維末梢連接下丘腦促性腺激素釋放激素(GnRH)神經(jīng)元[4],而GPR54表達(dá)于GnRH神經(jīng)元,表明kisspeptin直接調(diào)控GnRH的脈沖釋放[5-6]。綿羊腦室內(nèi)注射kisspeptin試驗(yàn)表明,kisspeptin直接作用于下丘腦-垂體-性腺軸,促進(jìn)GnRH釋放和LH水平上升,調(diào)控季節(jié)性繁殖活動(dòng)[7-8]。RFRP基因編碼蛋白最早于2000年從日本鵪鶉大腦中提取出來(lái),是定位于下丘腦室旁核的十二肽,因其具有抑制促性腺激素的作用又命名為促性腺激素抑制激素(Gonadotropin-inhibitory hormone,GnIH)[9]。RFRP神經(jīng)元與GnRH神經(jīng)元緊密毗鄰并投射到正中隆起[10],RFRP成熟肽可以抑制嚙齒動(dòng)物及綿羊促性腺激素的合成釋放[11]。KiSS-1與RFRP基因編碼蛋白同屬RFamide神經(jīng)肽類,但作用正好相反,兩者均受到動(dòng)物季節(jié)性光照信號(hào)內(nèi)源分子褪黑激素調(diào)控[12-14]和雌激素負(fù)反饋?zhàn)饔肹15-16],因而相互協(xié)調(diào)作用共同控制動(dòng)物季節(jié)性繁殖活動(dòng)[17]。
我國(guó)多數(shù)山羊品種屬于季節(jié)性繁殖,而目前對(duì)山羊季節(jié)性繁殖遺傳基礎(chǔ)不清楚。我國(guó)山羊品種眾多,繁殖季節(jié)性各異,遼寧絨山羊?qū)偌竟?jié)性繁殖,一般集中于秋冬發(fā)情繁殖[18],而濟(jì)寧青山羊具有常年發(fā)情的特點(diǎn),一年四季均可繁殖[19]。本研究以季節(jié)性發(fā)情遼寧絨山羊和常年發(fā)情濟(jì)寧青山羊作為對(duì)比品種,選擇KiSS-1和RFRP作為山羊季節(jié)性發(fā)情調(diào)控候選基因,通過反轉(zhuǎn)錄PCR和熒光定量PCR技術(shù)分別研究?jī)蓚€(gè)基因在兩個(gè)品種中的組織表達(dá)譜和在下丘腦-垂體-性腺軸的表達(dá)差異,從轉(zhuǎn)錄水平初步揭示兩基因在山羊季節(jié)性繁殖中的作用,為獲得繁殖季節(jié)性關(guān)鍵基因提供參考。
1 材料與方法
1.1 試驗(yàn)動(dòng)物和組織樣品采集
于2011年4月中下旬在農(nóng)業(yè)部濟(jì)寧青山羊保種基地(山東省嘉祥縣)和遼寧省遼陽(yáng)市遼寧絨山羊育種中心選擇健康、空懷濟(jì)寧青山羊與遼寧絨山羊成年母羊各3只,屠宰后迅速采集新鮮組織并放置于預(yù)先裝有RNA固定液的無(wú)RNase離心管中,樣品采集在1 h內(nèi)完成。所采組織冷藏(4 ℃)過夜后轉(zhuǎn)入-20 ℃保存,干冰運(yùn)輸帶回實(shí)驗(yàn)室轉(zhuǎn)入-80 ℃超低溫冷凍保存?zhèn)溆谩?/p>
1.2 RNA提取和反轉(zhuǎn)錄
利用高純總RNA快速提取試劑盒(百泰克,北京)提取組織總RNA,其中裂解液用Trizol(Invitrogen,美國(guó))替代。用DNaseⅠ(天根,北京)消化基因組DNA。將紫外分光光度計(jì)和瓊脂糖凝膠電泳檢測(cè)無(wú)降解,且A260 nm/A280 nm在1.8~2.1 的RNA置于-80 ℃冷凍備用。
使用cDNA快速合成試劑盒(天根,北京)反轉(zhuǎn)錄合成cDNA,反轉(zhuǎn)錄體系為20 μL:10×RT mix 2 μL,dNTPs (2.5 mmol·L-1) 2 μL,Oligo-dT15 2 μL,Quant Reverse Transcriptase 1 μL,1 μg總RNA,無(wú)RNase H2O補(bǔ)足20 μL。反應(yīng)程序:37 ℃溫育 60 min。反轉(zhuǎn)錄產(chǎn)物稀釋后,用GAPDH持家基因RT-PCR檢測(cè)后,-20 ℃保存,以備表達(dá)檢測(cè)所用。
1.3 引物設(shè)計(jì)
根據(jù)山羊KiSS-1基因序列(GenBank登錄號(hào):GU142847和AB433789)和RFRP基因序列(GenBank登錄號(hào):JF327669)各設(shè)計(jì)1對(duì)表達(dá)檢測(cè)引物,內(nèi)參基因選用甘油醛-3-磷酸脫氫酶(GAPDH),其RT-PCR引物采用課題組成熟引物[20],引物均由Invitrogen合成(引物序列見表1)。
表1 擴(kuò)增目的基因與內(nèi)參基因的引物信息
Table 1 Informations of primers amplifying target and internal control genes

基因引物Primer引物序列(5'-3')Sequence片段大小/bpSize退火溫度/℃TmKiSS-1F:ATGAACGTGCTGCTTTCCTR:TCCGAGCTGCGAGCCTGTG11760RFRPF:GCCTTTCCAGACGGGTCCCAR:GGCAGGTCATGGAGTAAAGT14460GAPDHF:GAGAAACCTGCCAAGTATGAR:CGAAGGTAGAAGAGTGAGTG13960GAPDH(RT-PCR用)F:AGGCTGGGGCTCACTTGAAGR:ATGGCGTGGACAGTGGTCAT22359
1.4 RT-PCR擴(kuò)增與測(cè)序
PCR體系為20 μL:含2×PCR Master Mix(博邁德,北京)10 μL,上、下游引物(10 mmol·L-1)各0.3 μL,cDNA 1.5 μL,ddH2O 7.9 μL。PCR程序:95 ℃預(yù)變性5 min;95 ℃變性30 s,60 ℃退火30 s,72 ℃延伸30 s,34次循環(huán);72 ℃延伸5 min。擴(kuò)增產(chǎn)物片段經(jīng)1.5%瓊脂糖凝膠電泳檢測(cè)后進(jìn)行回收,連接pMD19-T載體(TaKaRa,大連),轉(zhuǎn)化大腸桿菌TOP10感受態(tài)細(xì)胞,每個(gè)基因片段挑選3個(gè)陽(yáng)性克隆進(jìn)行擴(kuò)大培養(yǎng),并送北京六合華大基因股份有限公司進(jìn)行雙向測(cè)序。
1.5 熒光定量
1.5.1 熒光定量PCR體系和程序 熒光定量PCR體系為15 μL:UltraSYBR Mixture (with ROX)(CWBIO,北京) 10 μL,上、下游引物各0.4 μL,cDNA 2.0 μL,無(wú)RNase H2O 7.2 μL。PCR程序:95 ℃ 預(yù)變性10 min;95 ℃ 變性15 s,60 ℃延伸60 s,40個(gè)循環(huán);熒光定量PCR程序后做熔解曲線分析。
1.5.2 標(biāo)準(zhǔn)曲線的建立 取適合的cDNA樣本,進(jìn)行5倍等比稀釋制備5個(gè)濃度梯度(1、1/5、(1/5)2、(1/5)3、(1/5)4)的cDNA樣品。以這些cDNA模板對(duì)目的基因及內(nèi)參基因進(jìn)行熒光定量PCR,以獲得的Ct值為縱坐標(biāo),以濃度梯度的對(duì)數(shù)值(10為底)為橫坐標(biāo),繪制目的基因及內(nèi)參GAPDH基因的標(biāo)準(zhǔn)曲線。
1.5.3 熒光定量與數(shù)據(jù)分析 熒光定量PCR全部在LC480熒光定量PCR機(jī)器上進(jìn)行,每個(gè)樣品重復(fù)檢測(cè)3次,設(shè)置陰性對(duì)照(模板為H2O),內(nèi)參基因GAPDH。采用2-ΔΔCt法[21-22]計(jì)算相對(duì)表達(dá)量,運(yùn)用SAS 8.1(ANOVA)分析基因表達(dá)差異。
2 結(jié) 果
2.1 RNA提取與cDNA合成
瓊脂糖凝膠電泳檢測(cè)發(fā)現(xiàn),28S亮度大于18S,5S很淡(圖1a),表明RNA完整性好,無(wú)明顯降解。以反轉(zhuǎn)錄產(chǎn)物cDNA為模板,進(jìn)行GAPDH基因擴(kuò)增,結(jié)果發(fā)現(xiàn)內(nèi)參基因擴(kuò)增良好(圖1b)。
2.2KiSS-1與RFRP基因組織表達(dá)譜
KiSS-1、RFRP和GAPDH基因RT-PCR擴(kuò)增結(jié)果如圖2所示。濟(jì)寧青山羊和遼寧絨山羊24個(gè)組織GAPDH內(nèi)參基因均擴(kuò)增良好,亮度基本一致,差異不明顯。濟(jì)寧青山羊KiSS-1基因主要表達(dá)于下丘腦、垂體、松果體、小腦、丘腦和腦橋等腦部組織及腎、卵巢、心、脂肪和甲狀腺組織也有一定表達(dá)。遼寧絨山羊KiSS-1基因主要表達(dá)組織與濟(jì)寧青山羊相似,但其子宮體有一定表達(dá)而卵巢表達(dá)很微弱。濟(jì)寧青山羊RFRP基因主要表達(dá)于下丘腦、大腦皮層、小腦、卵巢和肺。遼寧絨山羊RFRP基因主要表達(dá)于下丘腦、垂體、大腦皮層、小腦、丘腦以及卵巢、心、腎、皮下脂肪和甲狀腺等組織。
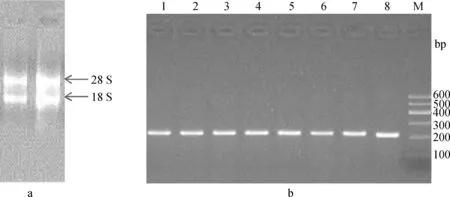
a.總RNA提取結(jié)果;b.部分組織GAPDH持家基因RT-PCR擴(kuò)增結(jié)果:1~8.下丘腦、垂體、松果體、卵巢、子宮體、子宮角、甲狀腺和脾;M.DNA marker Ⅰa.Gel electrophoresis of total RNA;b.RT-PCR amplification of housekeeping gene GAPDH in some tissues:1-8.hypothalamus,pituitary,pineal,ovary,corpus uteri,cornua uteri,thyroid and spleen,respectively;M.DNA marker Ⅰ圖1 組織總RNA提取和反轉(zhuǎn)錄合成cDNAFig.1 The quality of total RNA and cDNA

a.濟(jì)寧青山羊;b.遼寧絨山羊。1~24.下丘腦、垂體、松果體、大腦皮層、小腦、丘腦、海馬、腦橋、延髓、卵巢、子宮體、子宮角、輸卵管、腎上腺、肝、心、脊髓、腎、肺、胰、脾、骨骼肌、皮下脂肪以及甲狀腺;M.DNA 相對(duì)分子質(zhì)量標(biāo)準(zhǔn)a.Jining Grey goat;b.Liaoning Cashmere goat.1.Hypothalamus;2.Pituitary gland;3.Pineal;4.Cerebral cortex;5.Cerebellum;6.Thalamus;7.Hippocampus;8.Pons;9.Medulla oblongata;10.Ovary;11.Uterus;12.Uterine horn;13.Oviduct;14.Adrenal gland;15.Liver;16.Heart;17.Spinal cord;18.Kidney;19.Lung;20.Pancreas;21.Spleen;22.Skeletal;23.Subcutaneous fat;24.Thyroid;M.DNA marker Ⅰ圖2 濟(jì)寧青山羊與遼寧絨山羊KiSS-1、RFRP和GAPDH基因在各組織中的RT-PCR結(jié)果Fig.2 RT-PCR analysis of KiSS-1,RFRP and GAPDH gene expression in different tissues in Jining Grey goat and Liaoning Cashmere goat
2.3 山羊KiSS-1和RFRP基因在繁殖軸的表達(dá)水平
通過RT-PCR技術(shù)定性檢測(cè)了KiSS-1和RFRP基因在兩個(gè)山羊品種的表達(dá)情況,但結(jié)果很難精確定量,因而運(yùn)用了熒光定量PCR技術(shù)對(duì)兩個(gè)基因在下丘腦-垂體-卵巢和子宮等繁殖調(diào)控通路組織的表達(dá)水平進(jìn)行了研究,結(jié)果見圖3和圖4。
KiSS-1基因除在濟(jì)寧青山羊子宮中沒有檢測(cè)到表達(dá)外,兩個(gè)山羊品種其他組織中均有表達(dá)。濟(jì)寧青山羊KiSS-1基因在下丘腦中表達(dá)量最高,但與垂體表達(dá)量差異不顯著(P>0.05),而前兩者表達(dá)量顯著高于卵巢(P<0.05)。遼寧絨山羊垂體組織表達(dá)量顯著高于其他組織(P<0.05)。兩個(gè)品種相比較發(fā)現(xiàn),濟(jì)寧青山羊下丘腦表達(dá)量顯著高于遼寧絨山羊(P<0.05),而品種間其他組織KiSS-1基因表達(dá)無(wú)明顯差異。
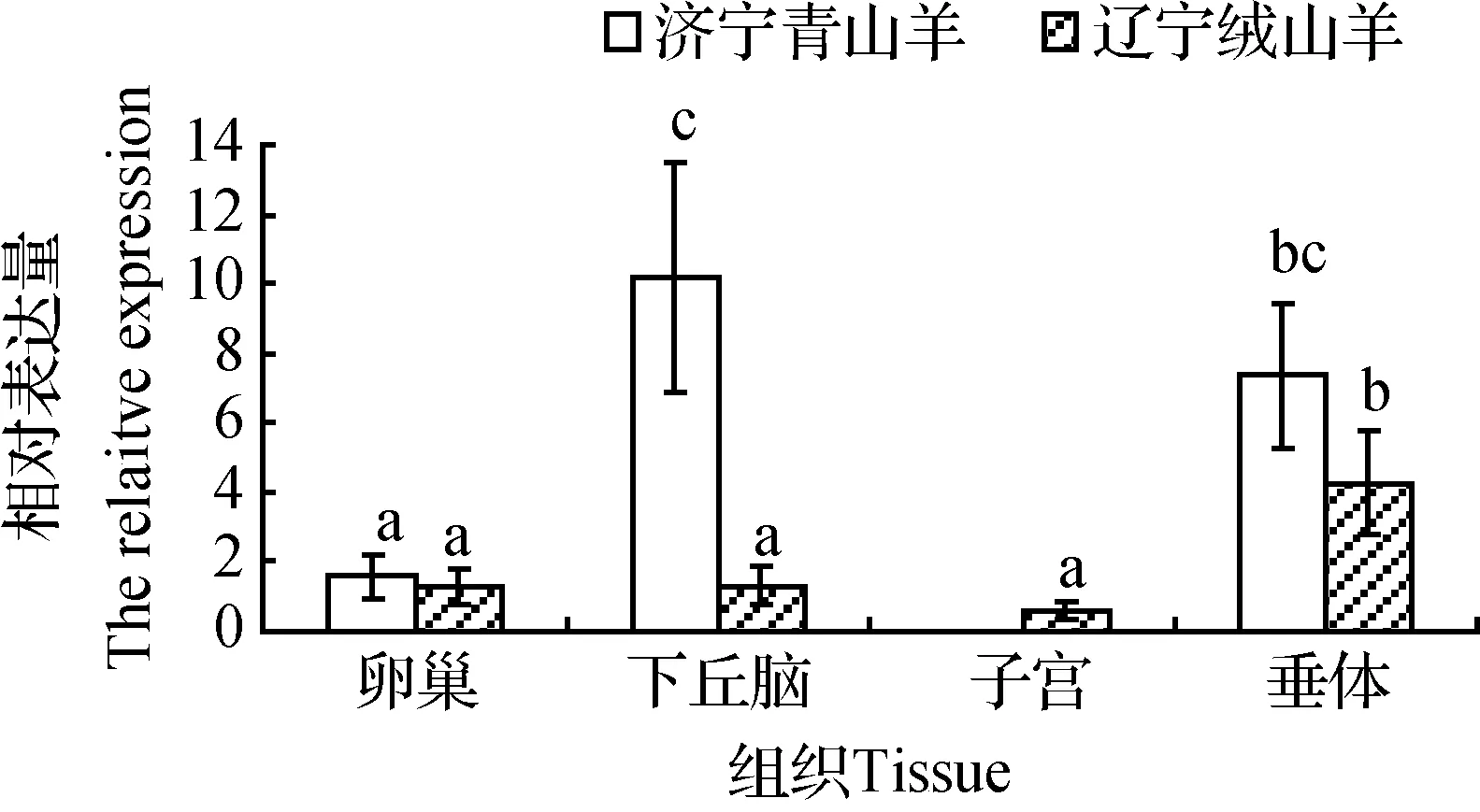
圖中表達(dá)量標(biāo)注相同字母表示差異不顯著(P>0.05),標(biāo)注不同字母表示差異顯著(P<0.05)。圖4同The expression levels with the same superscript have no significant difference (P>0.05).The expression levels with the different superscripts differ significantly (P<0.05).The same as Figure 4圖3 KiSS-1基因在卵巢、下丘腦、子宮、垂體組織中的表達(dá)水平Fig.3 Expression levels of KiSS-1 gene in ovary,hypothalamus,uterus and pituitary
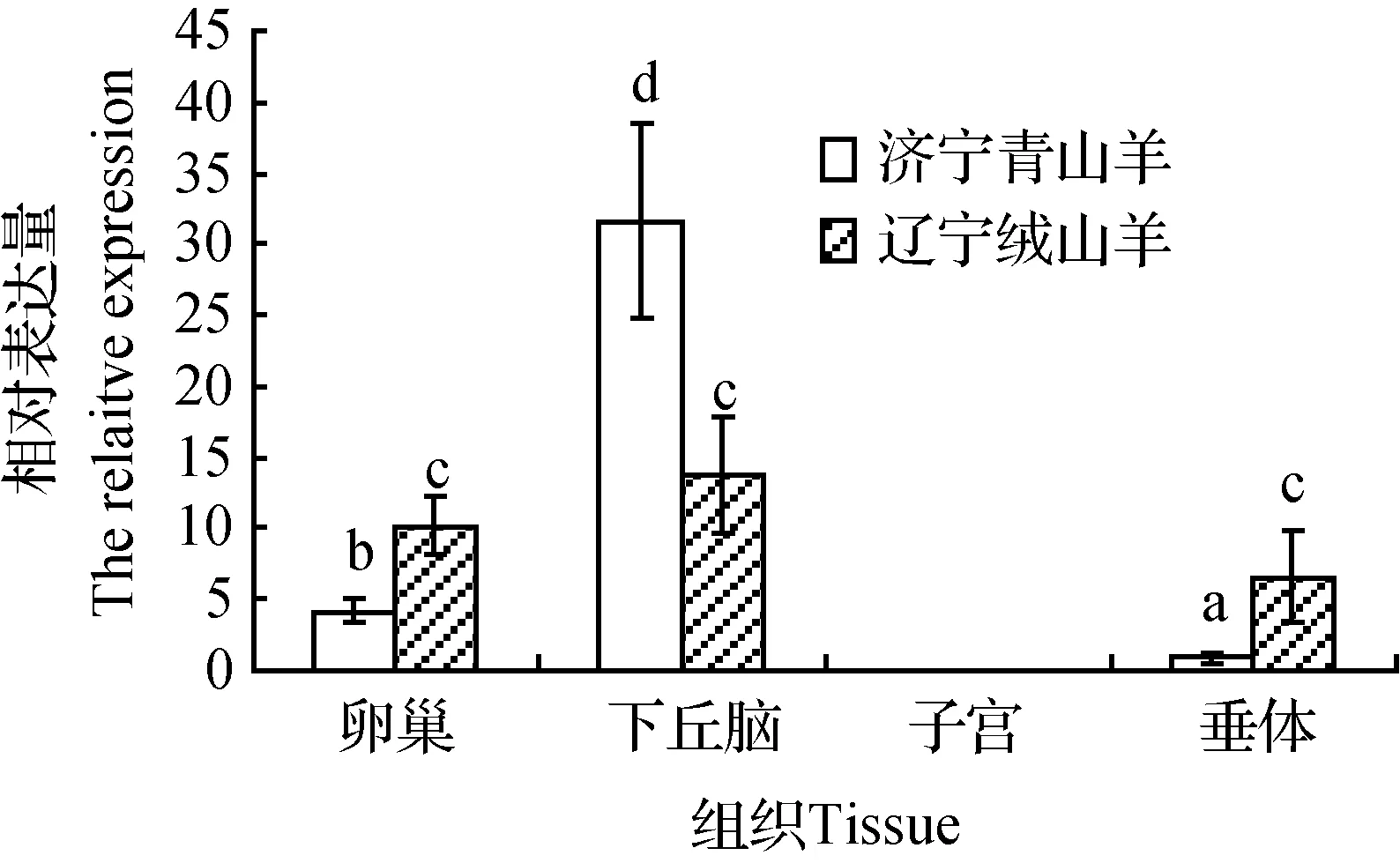
圖4 RFRP基因在卵巢、下丘腦、子宮、垂體組織的表達(dá)水平Fig.4 Expression levels of RFRP gene in ovary,hypothalamus,uterus and pituitary
RFRP基因在兩個(gè)山羊品種下丘腦、垂體和卵巢中均表達(dá),但在子宮組織沒有檢測(cè)到。濟(jì)寧青山羊下丘腦RFRP基因表達(dá)量明顯高于卵巢(P<0.05),而卵巢高于垂體(P<0.05)。遼寧絨山羊下丘腦、卵巢和垂體RFRP基因表達(dá)無(wú)明顯差異。兩個(gè)品種相比較發(fā)現(xiàn),濟(jì)寧青山羊下丘腦RFRP基因表達(dá)量高于遼寧絨山羊(P<0.05),而遼寧絨山羊垂體和卵巢表達(dá)量均顯著高于濟(jì)寧青山羊(P<0.05)。
3 討 論
3.1KiSS-1基因表達(dá)與季節(jié)性繁殖
J.H.Lee 等[2]應(yīng)用Northern雜交方法檢測(cè)發(fā)現(xiàn),人KiSS-1基因在胎盤中高表達(dá),在腎和胰腺中有微弱表達(dá),在心、大腦、肝、肺和骨骼肌中沒有檢測(cè)到表達(dá)。A.I.Muir等[23]用定量RT-PCR方法研究人20個(gè)組織及中樞神經(jīng)18個(gè)部位KiSS-1基因的表達(dá)定位,同樣發(fā)現(xiàn)胎盤KiSS-1基因表達(dá)量最高,并廣泛表達(dá)于下丘腦、海馬、丘腦、小腦和脊髓等中樞神經(jīng)組織,不過表達(dá)量較低。臧猛等[24]克隆了長(zhǎng)白和大白豬KiSS-1基因,并檢測(cè)其在18種組織的表達(dá)情況,結(jié)果發(fā)現(xiàn),KiSS-1基因在大腦、下丘腦、垂體、子宮、十二指腸、空腸、回腸、盲腸、結(jié)腸、直腸、心、肝、肺、腎、肌肉和脂肪均有表達(dá),僅卵巢和脾中沒有檢測(cè)到KiSS-1基因表達(dá)。本研究發(fā)現(xiàn),山羊KiSS-1基因在下丘腦、垂體、松果體、小腦、丘腦和腦橋等多個(gè)腦部組織表達(dá),同時(shí)腎、卵巢、心、脂肪和甲狀腺也有一定表達(dá),這與人類研究結(jié)果相似,但表達(dá)KiSS-1基因的內(nèi)臟組織明顯少于豬。
基于嚙齒動(dòng)物和綿羊KiSS-1基因在季節(jié)繁殖調(diào)控中的重要作用[25],本研究針對(duì)山羊下丘腦-垂體-卵巢-子宮繁殖軸組織KiSS-1基因表達(dá)進(jìn)行定量檢測(cè),與RT-PCR結(jié)果基本相符,同時(shí)在遼寧絨山羊卵巢檢測(cè)到KiSS-1基因表達(dá),這可能是定量PCR技術(shù)比RT-PCR更加靈敏所致;另外,濟(jì)寧青山羊子宮沒有檢測(cè)到KiSS-1基因表達(dá),與RT-PCR結(jié)果一致。品種對(duì)比研究發(fā)現(xiàn),濟(jì)寧青山羊下丘腦KiSS-1基因表達(dá)量顯著高于遼寧絨山羊,而本研究中遼寧絨山羊處于非繁殖季節(jié),而濟(jì)寧青山羊常年繁殖,推測(cè)下丘腦KiSS-1基因的高表達(dá)可能是導(dǎo)致濟(jì)寧青山羊與遼寧絨山羊繁殖季節(jié)性差異的原因之一。長(zhǎng)日照繁殖動(dòng)物敘利亞倉(cāng)鼠置于短日照條件下8~10周后其睪丸變小,血液睪酮水平降低,下丘腦ARC和AVPV內(nèi)KiSS-1基因mRNA表達(dá)均顯著下調(diào),KiSS-1神經(jīng)細(xì)胞數(shù)量顯著減少;而非季節(jié)性繁殖動(dòng)物Wistar試驗(yàn)大鼠在長(zhǎng)日照和短日照條件下丘腦ARC中KiSS-1基因mRNA表達(dá)水平相同,KiSS-1基因表達(dá)差異導(dǎo)致了嚙齒動(dòng)物繁殖季節(jié)性差異,這從側(cè)面支持了本研究結(jié)果[26]。短日照繁殖動(dòng)物綿羊在短日照時(shí)KiSS-1基因 mRNA表達(dá)量比長(zhǎng)日照時(shí)(休情)要高[27-28];而在休情季節(jié),注射kisspeptin可促使80%的綿羊發(fā)情并排卵,表明kisspeptin是啟動(dòng)母綿羊季節(jié)性繁殖的關(guān)鍵因子[8]。Abadeh母山羊繁殖季節(jié)卵泡期下丘腦kisspeptin神經(jīng)元明顯多于黃體期和休情季節(jié),而kisspeptin 10可以促進(jìn)山羊促黃體素和睪酮的釋放,這強(qiáng)有力地支持了本研究結(jié)果[29-31]。當(dāng)然,本研究缺乏蛋白表達(dá)和短日照繁殖季節(jié)對(duì)比數(shù)據(jù),需要進(jìn)一步深入研究。
3.2RFRP基因表達(dá)與季節(jié)性繁殖
目前RFRP基因表達(dá)譜研究不多,T.Ikemoto等[32]研究發(fā)現(xiàn),雞RFRP基因mRNA僅表達(dá)于中腦,而在垂體、卵巢、睪丸等其他17種組織均無(wú)表達(dá)。甘超等[33]應(yīng)用熒光定量PCR研究發(fā)現(xiàn),天府肉鵝母鵝GnIH前體基因主要表達(dá)于下丘腦,延髓和脊髓也有一定表達(dá),其他神經(jīng)組織幾乎無(wú)表達(dá),在腎上腺、輸卵管和卵巢也有一定表達(dá)。天府肉鵝4~5 mm直徑卵泡GnIH基因表達(dá)量均高于其他各級(jí)卵泡,因而GnIH基因可能參與卵泡選擇和閉鎖的調(diào)控[34]。用半定量RT-PCR方法檢測(cè)了新西蘭白兔33種組織GnIHmRNA的表達(dá),結(jié)果發(fā)現(xiàn),GnIH在眼球壁、下丘腦、肝、腎和腎上腺表達(dá)較高,在垂體、脊髓、腦橋、小腦、延髓、海馬、卵巢、睪丸、附睪、心、脾和胰也有表達(dá)[35]。原位雜交檢測(cè)發(fā)現(xiàn),綿羊RFRP基因在下丘腦內(nèi)側(cè)基底部、室旁核、背內(nèi)側(cè)核等廣泛表達(dá),僅腹中核沒有表達(dá)[36]。本研究檢測(cè)了兩個(gè)山羊品種24個(gè)組織RFRP基因表達(dá),結(jié)果發(fā)現(xiàn),RFRP基因主要表達(dá)于山羊下丘腦、大腦皮層、小腦、卵巢,這與兔的研究結(jié)果相似;另外品種間表達(dá)組織存在一定差異,遼寧絨山羊垂體、皮下脂肪和甲狀腺等組織均特異表達(dá)。
為闡明RFRP基因?qū)ι窖蚍敝臣竟?jié)性的影響,本研究通過定量PCR方法分析了常年發(fā)情濟(jì)寧青山羊和季節(jié)性發(fā)情遼寧絨山羊在春末時(shí)兩品種下丘腦-垂體-卵巢(子宮)繁殖調(diào)控軸組織RFRP基因的表達(dá)差異,結(jié)果發(fā)現(xiàn),RFRP基因均主要表達(dá)于下丘腦,這與家兔和綿羊等物種的研究結(jié)果相一致。然而,本研究還發(fā)現(xiàn),常年發(fā)情濟(jì)寧青山羊下丘腦RFRP表達(dá)量高于休情期遼寧絨山羊,這與其“GnRH抑制激素”之名并不符合。J.T.Smith等[37]研究也表明,RFRP基因表達(dá)升高很可能導(dǎo)致了Blackface母綿羊長(zhǎng)日照條件的季節(jié)性休情,這與本研究結(jié)果正好相反。不過,對(duì)西伯利亞倉(cāng)鼠和敘利亞倉(cāng)鼠的研究表明,RFRP基因?qū)Σ溉閯?dòng)物繁殖活動(dòng)具有激活作用[13,38],下丘腦RFRP基因表達(dá)上調(diào)促成其長(zhǎng)日照繁殖狀態(tài)[39]。另外還有研究發(fā)現(xiàn),綿羊RFRP基因表達(dá)變化可能并不與其繁殖活性有必然的關(guān)聯(lián)性[40]。可見,RFRP基因?qū)Σ溉閯?dòng)物繁殖的作用尚無(wú)定論。T.Ubuka等[39]研究發(fā)現(xiàn),給長(zhǎng)日照時(shí)西伯利亞倉(cāng)鼠注射RFRP1或RFRP3抑制其繁殖,而短日照時(shí)則激活繁殖,結(jié)果表明,RFRP基因只有適量表達(dá)才會(huì)激活繁殖,而表達(dá)過高或較低均抑制繁殖。因而,本研究結(jié)果較合理的解釋是濟(jì)寧青山羊常年發(fā)情特性可能與其下丘腦RFRP基因表達(dá)適量有關(guān),而遼寧絨山羊季節(jié)性休情因下丘腦RFRP基因表達(dá)較低所致,當(dāng)然這需要進(jìn)一步驗(yàn)證。本研究中,濟(jì)寧青山羊處于發(fā)情間期,其表達(dá)量較高還可能與發(fā)情周期所處階段有一定關(guān)系,這可以從對(duì)大鼠和豬的研究結(jié)果中獲得啟示[41-42]。此外,本研究還發(fā)現(xiàn),濟(jì)寧青山羊卵巢和垂體RFRP基因表達(dá)量均顯著低于遼寧絨山羊,RFRP基因是否通過外周組織而抑制山羊繁殖?S.Zhao等[43]研究發(fā)現(xiàn),RFRP及其受體基因mRNA均在敘利亞大鼠精細(xì)胞和精子細(xì)胞表達(dá),并受到日照長(zhǎng)短和繁殖條件的顯著影響,因而RFRP基因可能通過外周繁殖組織自分泌/旁分泌作用而參與繁殖調(diào)控。目前這方面研究很少,因而山羊卵巢和垂體組織RFRP基因表達(dá)與其繁殖季節(jié)性的關(guān)系值得深入研究。
4 結(jié) 論
山羊KiSS-1基因主要表達(dá)于下丘腦、垂體、松果體等腦部組織和腎、卵巢、脂肪和甲狀腺組織,山羊RFRP基因主要表達(dá)于下丘腦、大腦皮層、小腦、卵巢和垂體等組織,山羊兩基因組織表達(dá)譜存在一定的品種特異性。KiSS-1基因在常年發(fā)情濟(jì)寧青山羊下丘腦表達(dá)顯著高于季節(jié)性發(fā)情遼寧絨山羊,提示其可能對(duì)山羊季節(jié)性繁殖有重要的調(diào)控作用,山羊下丘腦KiSS-1基因高表達(dá)可能是非繁殖季節(jié)常年發(fā)情品種能夠繁殖的原因。而下丘腦RFRP基因表達(dá)量與繁殖抑制或激活的關(guān)系需要深入的研究,RFRP基因在卵巢和垂體表達(dá),并且濟(jì)寧青山羊明顯低于遼寧絨山羊,研究結(jié)果提示,RFRP基因可能通過下丘腦以外組織對(duì)山羊季節(jié)繁殖活動(dòng)進(jìn)行調(diào)控。
[1] GREIVES T J,MASON A O,SCOTTI M A L,et al.Environmental control of kisspeptin:implications for seasonal reproduction [J].Endocrinology,2007,148(3):1158-1166.
[2] LEE J H,MIELE M E,HICKS D J,et al.KiSS-1,a novel human malignant melanoma metastasis-suppressor gene [J].JNatlCancerInst,1996,88(23):1731-1737.[3] OHTAKI T,SHINTANI Y,HONDA S,et al.Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor [J].Nature,2001,411(6837):613-617.
[4] FRANCESCHINI I,LOMET D,CATEAU M,et al.Kisspeptin immunoreactive cells of the ovine preoptic area and arcuate nucleus co-express estrogen receptor alpha [J].NeurosciLett,2006,401(3):225-230.
[5] IRWIG M S,F(xiàn)RALEY G S,SMITH J T,et al.Kisspeptin activation of gonadotropin releasing hormone neurons and regulation of KiSS-1 mRNA in the male rat [J].Neuroendocrinology,2004,80(4):264-272.
[6] HAN S K,GOTTSCH M L,LEE K J,et al.Activation of gonadotropin-releasing hormone neurons by kisspeptin as a neuroendocrine switch for the onset of puberty [J].JNeurosci,2005,25(49):11349-11356.
[7] MESSAGER S,CHATZIDAKI E E,MA D,et al.Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54 [J].ProcNatlAcadSciUSA,2005,102(5):1761-1766.[8] CARATY A,SMITH J T,LOMET D,et al.Kisspeptin synchronizes preovulatory surges in cyclical ewes and causes ovulation in seasonally acyclic ewes [J].Endocrinology,2007,148(11):5258-5267.
[9] TSUTSUI K,SAIGOH E,UKENA K,et al.A novel avian hypothalamic peptide inhibiting gonadotropin release [J].BiochemBiophysResCommun,2000,275(2):661-667.
[10] UBUKA T,MCGUIRE N L,CALISI R M,et al.The control of reproductive physiology and behavior by gonadotropin-inhibitory hormone [J].IntegrCompBiol,2008,48(5):560-569.
[11] SARI I P,RAO A,SMITH J T,et al.Effect of RFamide-related peptide-3 on luteinizing hormone and follicle-stimulating hormone synthesis and secretion in ovine pituitary gonadotropes [J].Endocrinology,2009,50(12):5549-5556.
[12] MALPAUX B,DAVEAU A,MAURICE-MANDON F,et al.Evidence that melatonin acts in the premammillary hypothalamic area to control reproduction in the ewe:presence of binding sites and stimulation of luteinizing hormone secretion by in situ microimplant delivery [J].Endocrinology,1998,139(4):1508-1516.[13] REVEL F G,SABOUREAU M,PéVET P,et al.RFamide-related peptide gene is a melatonin-driven photoperiodic gene [J].Endocrinology,2008,149(3):902-912.
[14] GINGERICH S,WANG X,LEE P K,et al.The generation of an array of clonal,immortalized cell models from the rat hypothalamus:analysis of melatonin effects on kisspeptin and gonadotropin-inhibitory hormone neurons [J].Neuroscience,2009,162(4):1134-1140.[15] SMITH J T.Sex steroid control of hypothalamic Kiss1 expression in sheep and rodents:comparative aspects [J].Peptides,2009,30(1):94-102.
[17] SMITH J T.The role of kisspeptin and gonadotropin inhibitory hormone in the seasonal regulation of reproduction in sheep [J].DomestAnimEndocrinol,2012,43(2):75-84.
[18] 姜懷志,宋先忱,張世偉.遼寧絨山羊繁殖生物學(xué)特性[J].中國(guó)草食動(dòng)物,2010,(z1):429-432. JIANG H Z,SONG X C,ZHANG S W.Biological characteristics of reproduction in Liaoning Cashmere goat [J].ChinaHerbivoreScience,2010,(Supplement):429-432.(in Chinese)
[19] 涂友仁.中國(guó)羊品種志[M].上海:上海科學(xué)技術(shù)出版社,1989. TU Y R.The Sheep and Goat Breeds in China [M].Shanghai:Shanghai Science and Technology Press,1989.(in Chinese)
[20] 曹貴玲.SSH篩選濟(jì)寧青山羊性早熟相關(guān)基因及KiSS-1、GPR54、Lin28B基因的研究[D].北京:中國(guó)農(nóng)業(yè)科學(xué)院,2011. CAO G L.Screening of sexual precocity related genes in Jining Grey goat using SSH and study onKiSS-1,GPR54 andLin28B genes [D].Beijing:Chinese Academy of Agricultural Sciences,2011.(in Chinese)
[21] LIVAK K J,SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method [J].Methods,2001,25(4):402-408.
[22] SCHMITTGEN T D,LIVAK K J.Analyzing real-time PCR data by the comparative C(T) method [J].NatProtoc,2008,3(6):1101-1108.
[23] MUIR A I,CHAMBERLAIN L,ELSHOURBAGY N A,et al.AXOR12,a novel human G protein-coupled receptor,activated by the peptide KiSS-1 [J].JBiolChem,2001,276(31):28969-28975.
[24] 臧 猛,晁利剛,張志強(qiáng),等.豬KISS-1基因克隆、序列分析[J].江西農(nóng)業(yè)學(xué)報(bào),2008,20(11):5-7. ZANG M,CHAO L G,ZHANG Z Q,et al.Molecular cloning and sequence analysis of porcine KISS-1 gene[J].ActaAgriculturaeJiangxi,2008,20(11):5-7.(in Chinese)
[25] CLARKE I J,CARATY A.Kisspeptin and seasonality of reproduction [J].AdvExpMedBiol,2013,784:411-430.
[26] REVEL F G,SABOUREAU M,MASSON-PéVET M,et al.Kisspeptin mediates the photoperiodic control of reproduction in hamsters [J].CurrBiol,2006,16(17):1730-1735.
[27] SMITH J T,CLAY C M,CARATY A,et al.KiSS-1 messenger ribonucleic acid expression in the hypothalamus of the ewe is regulated by sex steroids and season [J].Endocrinology,2007,148(3):1150-1157.
[28] WAGNER G C,JOHNSTON J D,CLARKE I J,et al.Redefining the limits of day length responsiveness in a seasonal mammal [J].Endocrinology,2008,149(1):32-56.
[29] JAFARZADE H,SHIRAZI M R,ZAMIRI M J,et al.Differential expression of RFamide-related peptide,a mammalian gonadotrophin-inhibitory hormone orthologue,and kisspeptin in the hypothalamus of Abadeh ecotype does during breeding and anoestrous seasons [J].JNeuroendocrinol,2014,26(3):186-194.
[30] SAITO H,SAWADA T,YAEGASHI T,et al.Kisspeptin-10 stimulates the release of luteinizing hormone and testosterone in pre- and post-pubertal male goats [J].AnimSciJ,2012,83(6):487-492.
[31] HASHIZUME T,SAITO H,SAWADA T,et al.Characteristics of stimulation of gonadotropin secretion by kisspeptin-10 in female goats [J].AnimReprodSci,2010,118(1):37-41.
[32] IKEMOTO T,PARK M K.Chicken RFamide-related peptide (GnIH) and two distinct receptor subtypes:identification,molecular characterization,and evolutionary considerations [J].JReprodDev,2005,51(3):359-377.
[33] 甘 超,胡深強(qiáng),茍 華,等.鵝GnIH受體基因的克隆及組織表達(dá)特性研究[J].中國(guó)家禽,2014,36(1):7-11. GAN C,HU S Q,GOU H,et al.Cloning of gonadotropin-inhibiting hormone receptor genes and their expression patterns in tissues of geese [J].ChinaPoultry,2014,36(1):7-11.(in Chinese)
[34] 董 霞.鵝等級(jí)前卵泡發(fā)育研究及GnIH在等級(jí)前卵泡中的表達(dá)分析[D].雅安:四川農(nóng)業(yè)大學(xué),2013. DONG X.The developmental study of geese pre-hierarchical follicles and the analysis ofGnIHexpression in pre-hierarchical follicles [D].Ya’an:Sichuan Agricultural University,2013.(in Chinese)
[35] 劉 晶.兔GnIH及其受體基因的克隆和mRNA在組織器官的表達(dá)[D].南京:南京農(nóng)業(yè)大學(xué),2011. LIU J.The cloning and mRNA expression of GnIH and its receptor genes in rabbit[D].Nanjing:Nanjing Agricultural University,2011.(in Chinese)
[36] DARDENTE H,BIRNIE M,LINCOLN G A,et al.RFamide-related peptide and its cognate receptor in the sheep:cDNA cloning,mRNA distribution in the hypothalamus and the effect of photoperiod [J].JNeuroendocrinol,2008,20(11):1252-1259.
[37] SMITH J T,COOLEN L M,KRIEGSFELD L J,et al.Variation in kisspeptin and RFamide-related peptide (RFRP) expression and terminal connections to gonadotropin-releasing hormone neurons in the brain:a novel medium for seasonal breeding in the sheep [J].Endocrinology,2008,149(11):5770-5782.
[38] ANCEL C,BENTSEN A H,SéBERT M E,et al.Stimulatory effect of RFRP-3 on the gonadotrophic axis in the male Syrian hamster:the exception proves the rule [J].Endocrinology,2012,153(3):1352-1363.[39] UBUKA T,INOUE K,F(xiàn)UKUDA Y,et al.Identification,expression,and physiological functions of Siberian hamster gonadotropin-inhibitory hormone [J].Endocrinology,2012,153(1):373-385.
[40] PAPARGIRIS M M,RIVALLAND E T,CLARKE I J,et al.Evidence that RF-amide related peptide-3 is not a mediator of the inhibitory effects of psychosocial stress on gonadotrophin secretion in ovariectomised ewes[J].JNeuroendocrinol,2011,23(3):208-215.
[41] SALEHI M S,JAFARZADEH SHIRAZI M R,ZAMIRI M J,et al.Hypothalamic expression of KiSS1 and RFamide-related peptide-3 mRNAs during the estrous cycle of rats [J].IntJFertilSteril,2013,6(4):304-309.
[42] LI X,SU J,LEI Z,et al.Gonadotropin-inhibitory hormone (GnIH) and its receptor in the female pig:cDNA cloning,expression in tissues and expression pattern in the reproductive axis during the estrous cycle [J].Peptides,2012,36(2):176-185.
[43] ZHAO S,ZHU E,YANG C,et al.RFamide-related peptide and messenger ribonucleic acid expression in mammalian testis:association with the spermatogenic cycle [J].Endocrinology,2010,151(2):617-627.
(編輯 郭云雁)
Study on Expression ofKiSS-1 andRFRPGenes Related to Reproductive Seasonality in Goats
HUANG Dong-wei1,2,CHU Ming-xing1*,DI Ran1,LIU Qiu-yue1,HU Wen-ping1,WANG Xiang-yu1,PAN Zhang-yuan1,GUO Xiao-fei1
(1.KeyLaboratoryofFarmAnimalGeneticResourcesandGermplasmInnovationofMinistryofAgriculture,InstituteofAnimalScience,ChineseAcademyofAgriculturalSciences,Beijing100193,China;2.InstituteofAnimalHusbandryandVeterinaryMedicine,AnhuiAcademyofAgriculturalSciences,Hefei230031,China)
To elucidate the roles ofKiSS-1 andRFRPgenes in goat,their expression profilings in tissues and the expression levels in hypothalamus-pituitary-ovary (and uterus) were determined between Jining Grey goat (year-round estrus) and Liaoning Cashmere goat (seasonal estrus) by using RT-PCR and qRT-PCR methods,respectively.The results showed that:(i)KiSS-1 mRNA was detected in goat hypothalamus,pituitary,pineal,kidney,ovary,subcutaneous fat and thyroid,whileRFRPmRNA was detected in goat hypothalamus,cerebral cortex,cerebellum,ovary and pituitary,both of them had different tissue expression profilings between two goat breeds;(ii)KiSS-1 mRNA expression level was higher in hypothalamus for Jining Grey than that in Liaoning Cashmere goats (P<0.05),while the expression had no significant difference in ovary and pituitary between 2 breeds (P>0.05);(iii) The expression level ofRFRPmRNA was higher in hypothalamus of Jining Grey than that in Liaoning Cashmere goat (P<0.05),while its expression in ovary and pituitary was opposite between 2 breeds (P<0.05).The present study suggests thatKiSS-1 gene may involve in regulating seasonal breeding in goats,but the effect (or the acting manner) ofRFRPgene in goat seasonal breeding need be further researched.The results lay the foundation for elucidating the function of goatKiSS-1 andRFRPgenes and provide reference for revealing the genetic mechanism of reproductive seasonality in goat.
goat;reproductive seasonality;KiSS-1;RFRP;expression
10.11843/j.issn.0366-6964.2015.06.007
2014-08-27
中國(guó)農(nóng)業(yè)科學(xué)院科技創(chuàng)新工程(ASTIP-IAS13);國(guó)家自然科學(xué)基金項(xiàng)目(30871773);國(guó)家肉羊產(chǎn)業(yè)技術(shù)體系專項(xiàng)(CARS-39);轉(zhuǎn)基因生物新品種培育重大專項(xiàng)課題(2014ZX08010005-003)
黃冬維(1981-),男,湖南長(zhǎng)沙人,助理研究員,博士,主要從事草食動(dòng)物遺傳育種與健康養(yǎng)殖研究,E-mail:hdwwxf@163.com
*通信作者:儲(chǔ)明星,博士,研究員,博導(dǎo),E-mail:mxchu@263.net
S827;S813.3
A
0366-6964(2015)06-0924-08
