橡膠樹熱研7—33—97不同雜交組合子代早期鑒定研究
2015-05-30 10:48:04張源源高新生張曉飛黃肖李維國
熱帶作物學報 2015年8期
張源源 高新生 張曉飛 黃肖 李維國
摘 ?要 ?橡膠樹(Hevea brasiliensis)優良品種熱研7-33-97具有生長速度快、抗性強、產膠潛力大和干膠理化性能好等優點。為研究熱研7-33-97作為雜交親本選育優良新品種的潛力,分別對熱研2-73×熱研7-33-97、熱研5-11×熱研7-33-97、熱研7-33-97×海墾1的人工雜交授粉子代進行生長量和產量早期鑒定。結果表明:不同雜交組合子代的莖圍生長和產量均有顯著差異,熱研7-33-97作為父本或者母本均能較好地遺傳其速生、高產性狀。子代性狀的正態分析表明,從熱研2-73×熱研7-33-97雜交組合子代中選育出膠木兼優品種的可能性較大,從熱研5-11×熱研7-33-97雜交組合子代中選育出高產品種的可能性較大,從熱研7-33-97×海墾1雜交組合子代中有選育出抗風高產品種的可能性。
關鍵詞 ?橡膠樹;雜交組合;早期鑒定;莖圍生長;干膠產量
中圖分類號 ?S722.34 ? ? ? ? ?文獻標識碼 ?A
Early Identification of Different Hybrid of
CATAS 7-33-97 Rubber Tree
ZHANG Yuanyuan, GAO Xinsheng, ZHANG Xiaofei,
HUANG Xiao, LI Weiguo*
Rubber Research Institute,Chinese Academy of Tropical Agricultural Sciences/
State Centre for Rubber Breeding,Danzhou, Hainan 571737,China
Abstract ?CATAS 7-33-97 is a rubber tree(Hevea brasiliensis Müll. Arg)clone with characteristics of fast growth, strong resistance, high-yielding potential, good adhesive physical and chemical properties. The girth growth and dry rubber yield of different hybrid of CATAS 7-33-97 was identified in early period. Results showed that there was significant differences in girth growth and dry rubber yield among hybrid; The fast growth and high yield characters of CATAS 7-33-97 can pass to progeny as male and female parent well; The hybrid combination of CATAS 2-73×CATAS 7-33-97 has the potential to select out Latex/Timber clones; The hybrid of CATAS 5-11×CATAS 7-33-97 has the potential to select out high yield clones; The hybrid of CATAS 7-33-97×Haiken 1 has the potential to select out wind resistance clones.
Key words ?Hevea brasiliensis; Hybrid; Early identification; Girth growth; Dry rubber yield
doi ?10.3969/j.issn.1000-2561.2015.08.001
人工雜交授粉并對子代進行鑒定,選擇出優良單株進入無性系比較試驗程序是橡膠樹(Hevea brasiliensis)常規育種的基本方法。在此過程中,親本的選配及早期鑒定決定著從授粉子代中選擇出優異個體的效率。
熱研7-33-97是目前在海南大面積種植的優良橡膠樹品種,其親本組合為PR107×RRIM600[1],PR107為印度尼西亞選育的初生代無性系,具有后期產量高、耐乙烯利刺激、抗風能力強等優點,RRIM600為馬來西亞橡膠研究院選育,具有開割前后莖圍生長快、單位面積產量高等優點,以此雜交組合已經選育出了熱研7-20-59[2]、熱研7-18-55[3]、熱研6-881[4]、文昌11等高產品種。熱研7-33-97集合了2個親本的優良性狀,具有生長速度快、抗性強、產膠潛力大、干膠理化性能好等優點[5],同時雄花開花量大,可以充分滿足雜交授粉的需要,是橡膠樹品種改良和新品種選育的理想親本材料。目前,以熱研7-33-97為親本已得到部分優異育種材料。
橡膠樹的苗期產量預測方法前人研究較多,現已總結出刺檢法、試割法、形態預測法、葉脈膠法、小葉柄膠法、乳管計數法和葉脈檢定角法等多種早期預測方法,其中以3 a以后的幼樹試割法較為準確[6-8],這也是本試驗進行產量鑒定的主要方法。方家林等[9]、李維國等[10]在以野生種質為親本的F1群體中發現豐富的遺傳變異,并利用試割法等預測方法初篩了一批潛力育種中間材料,為橡膠樹親本組配和雜交子代早期鑒定提供了有益的參考。
本文對以熱研7-33-97分別為親本與部分優良品種雜交授粉的子代進行早期系統鑒定,研究其作為父本或母本對子代主要遺傳性狀的影響,評估不同雜交組合選育優良新品種的潛力,從而為熱研7-33-97作為雜交育種利用提供理論和實踐依據。
1 ?材料與方法
1.1 ?材料
1.1.1 ?供試材料及預處理 ? 以定植于中國熱帶農業科學院試驗場和國家橡膠樹種質資源圃的熱研7-33-97、熱研2-73、熱研5-11和海墾1為親本,于2003~2005年春花期進行人工授粉,采果播種育苗,其中熱研2-73×熱研7-33-97獲得子代授粉苗167株,熱研5-11×熱研7-33-97獲得授粉子代苗112株,熱研7-33-97×海墾1獲得子代授粉苗45株。
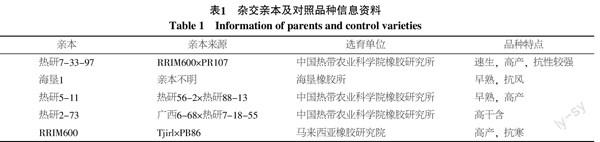
為獲得年齡相對一致的試驗材料,2006年10月,將獲得的授粉子代苗在距離地面30 cm處截干,以裸根苗形式定植于熱科院橡膠研究所五方潭育種基地。試驗區采用1.5 m×2 m株行距定植,參照《橡膠樹育種技術規程》隨機配置對照品種RRIM600。試驗地按照試驗要求撫管。雜交親本及對照品種信息見表1[1,11-13]。
1.1.2 ?試驗地概況 ? 試驗地位于海南省儋州市境內,19°30'N,109°30'E,年均溫度23.6 ℃,雨量充沛,年均降雨量1 704.7 mm,試驗地為﹤5°緩坡,土壤為磚紅壤。試驗地靠近小河流,四周有緩坡和農田,屬半開闊土地。
1.2 ?方法
2009和2011年,每年12月份逐株調查距地面1 m處莖圍。2010和2012年,每年9月采用試割法s/2d/3割制,50 cm高度,割10刀,前5刀產量不計,后5刀每次杯凝后取膠塊烘干稱重,計算每株試割干膠總產量對現存的264株授粉苗(其中熱研2-73×熱研7-33-97子代139株、熱研7-33-97×海墾1子代40株、熱研5-11×7-33-97子代85株)及13株對照苗進行產量早期鑒定。2010年測產同時測量割線長度。
1.3 ?數據處理
數據整理在Microsoft Excel 2013中進行,使用IBM SPSS Statistics22中文版進行多重比較和正態分布檢驗。
2 ?結果與分析
2.1 ?雜交子代莖圍生長
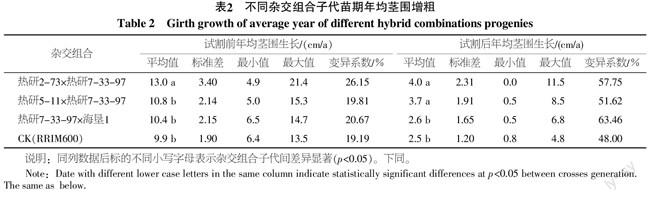
從橡膠樹不同組合雜交子代試割前和試割后的年均莖圍增粗可以看出(表2),不同雜交組合子代的生長速度不同,雜交組合內不同子代生長速度差異較大,橡膠樹子代試割前后生長速度差異較大。試割前,3個雜交組合子代的平均年均莖圍生長均超過10 cm,其中雜交組合熱研2-73×熱研7-33-97的子代平均年均莖圍生長達到13 cm,顯著高于對照品種RRIM600,其它組合及對照內均無顯著差異。試割后2 a,3個雜交組合子代的年均莖圍增粗均不超過4 cm,生長速度較試割前明顯降低,其中雜交組合熱研2-73×熱研7-33-97和熱研5-11×熱研7-33-97子代的平均年均增粗分別為4.0和3.7 cm,均顯著高于對照RRIM600的2.5 cm,熱研7-33-97×海墾1子代與對照之間無顯著差異。3個雜交組合子代在試割后年均莖圍生長變異系數分別達到57.75%、51.62%和63.46%,均較試割前明顯增大。
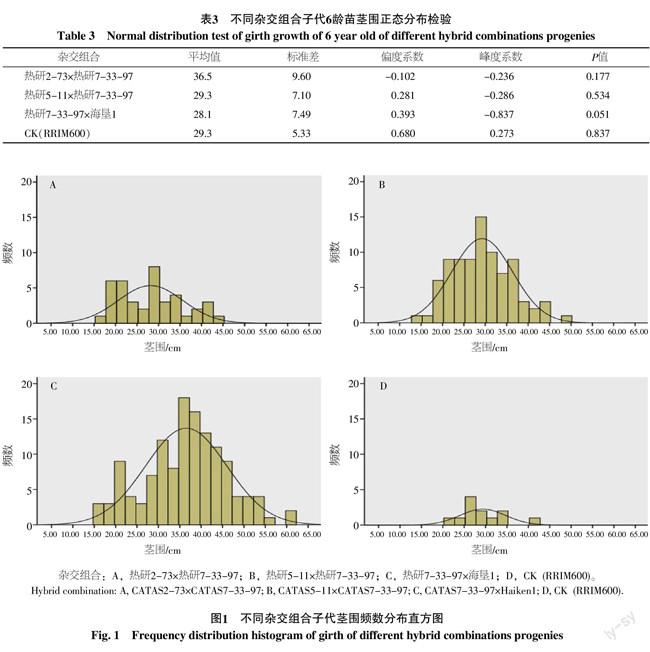
對3個雜交組合子代及對照在2012年的莖圍進行Shapiro-Wilk正態分布檢驗(表3),結果表明4個群體的莖圍分布均符合正態分布(圖1)。其中熱研2-73×熱研7-33-97子代莖圍分布的偏度系數和峰度系數分別為-0.102和-0.236,表明該雜交組合的子代傾向于生長較快,但性狀分離程度較大;熱研5-11×熱研7-33-97和熱研7-33-97×海墾1的子代莖圍分布的偏度系數和峰度系數分別為0.281和-0.286、0.393和-0.837,表明這2個雜交組合的子代傾向于生長較慢,且性狀分離程度較大。對照RRIM600無性系群體莖圍分布的偏度系數和峰度系數分別為0.680和0.273,表明其莖圍生長傾向于生長較慢,性狀分離程度較小,符合無性系群體的生長特點。
2.2 ?雜交子代試割產量
從雜交組合子代2次試割產量可以看出(表4),不同雜交組合子代的試割產量不同,雜交組合內子代的試割產量差異較大,因樹齡差異2次試割產量也不同。2010年試割時,3個雜交組合的子代和對照的平均單株產量均在25 g以下,其中雜交組合熱研7-33-97×海墾1子代的平均單株產量僅為14.9 g,顯著低于對照RRIM600,其余群體之間差異均不顯著。單位割線長度產量與平均單株產量表現不同,對照RRIM600顯著高于3個雜交組合子代,3個雜交組合之間差異不顯著。2012年試割時,3個雜交組合的子代及對照群體的平均單株產量均在35 g以上,3個子代群體的平均單株產量均與對照差異不顯著,熱研5-11×熱研7-33-97子代平均單株產量顯著高于其它雜交組合子代。各雜交組合子代群體產量的變異系數較大,均在70%以上,原因在于試割中部分單株沒有產膠,產量按照0計算。不同雜交組合的2次試割產量相關系數均在0.6左右,相關性極顯著,表明進行1次試割就可以滿足早期產量預測的要求。
對3個雜交組合子代和對照群體的2次試割總產量進行正態分布檢驗。2次試割總產量頻數分布直方圖(圖2)表明4個群體的產量分布均不符合正態分布,其中熱研2-73×熱研7-33-97子代2次試割總產量符合指數分布。3個雜交組合子代的產量大部分較低,高產單株較少,其中熱研2-73×熱研7-33-97組合子代此種趨勢最為明顯,熱研5-11×熱研7-33-97組合最不明顯,對照RRIM600也同樣不明顯。
2.3 ?雜交子代第一次選擇
橡膠樹雜交組合子代早期鑒定的目的是為了快速選擇出優良的單株進入無性系比較試驗程序。本試驗以膠木兼優為育種目標,因此子代優良單株的選擇標準設計為產量性狀與對照相當,即2次試割的平均產量不低于對照,同時綜合考慮開割前后莖圍生長速度表現,并參照單位割線長度產量。以此為標準,從3個雜交組合子代中選擇出優良單株進入無性系比較試驗程序,每一株均有2項或者以上指標表現超過對照平均表現(表5)。其中,從熱研2-73×熱研7-33-97子代中選擇出8株,選出率為5.76%;從熱研5-11×熱研7-33-97子代中選擇出11株,選出率為12.94%;從熱研7-33-97×海墾1子代中選擇出3株,選出率為7.50%。
3 ?討論與結論
熱研7-33-97無論作為父本或者母本均能較好地遺傳其優良性狀給子代。用熱研7-33-97分別作為父母本與不同品種進行雜交授粉,其作為母本的子代在早期生長和試割產量上均比其作為父本的子代較差,但是差異不顯著,這種微弱的差異可以歸結為與之雜交的親本不同,而熱研7-33-97無論是作為父本或者母本,均不影響其優良性狀遺傳到子代。方家林等[9]用熱研88-13做母本與野生種質59號雜交,子代優株選出率為1.66%,李維國等[10]用熱研88-13做母本與3份野生種質雜交,子代優株選出率分別為6.90%、1.12%和0%,均明顯低于熱研7-33-97的子代優株選出率。
橡膠樹熱研7-33-93的不同雜交組合之間的莖圍生長和產量表現差異較大。雜交組合熱研2-73×熱研7-33-97子代的平均莖圍增粗較快,快速生長單株所占比例較高,但是平均試割產量較低,高產單株所占比例較低,可以考慮從該雜交組合子代中選育膠木兼優品種;雜交組合熱研5-11×熱研7-33-97子代的平均莖圍增粗次之,快速生長單株所占比例一般,但其子代平均試割產量較高,高產單株所占比例較高,可以考慮從該雜交組合子代中選育高產品種。雜交組合熱研7-33-97×海墾1子代平均莖圍生長和平均試割產量均一般,但是考慮到海墾1具有較強的抗風能力,因此從該雜交組合中選擇在生長和產量表現較好的單株,進一步鑒定抗風能力,具有從中選育出抗風高產品種的可能性。
不同雜交組合子代的平均單株產量與單位割線產量表現不一致,表明割線長度是影響橡膠樹產膠量的因素之一。現有研究表明,乳管列數與干膠產量有密切關系[14-16]。在乳管列數相同的情況下,橡膠樹的割線越長,其干膠產量應越高。在當前的s/2d/3的割膠制度下,割線的長度由莖圍大小決定,因此莖圍大小間接決定著橡膠樹產量的高低。因此,優選單株時莖圍生長的速度和乳管列數,可以作為橡膠樹早期試割鑒定的輔助指標。莖圍和產量均屬于數量性狀[17],而不同雜交組合的子代群體分別屬于一個孟德爾群體[18],因此不同雜交組合子代的莖圍和產量應該呈正態分布[19]。本研究中不同雜交組合子代群體的莖圍均屬于正態分布,但產量分布則均不屬于正態分布。分析其原因,一種可能是子代群體較小導致的,另一種可能是產量性狀受主效基因控制的作用更加明顯,與產量相關的主效基因的分離導致子代產量表現出低產的傾向性。此外,因為樹體和年齡均較小,相當多的植株尚未產膠,也是導致群體出現正態偏離的原因,這也可能是對照群體的產量變異系數較雜交組合子代大的原因。
不同雜交組合子代在試割前的年均莖圍增粗均明顯高于試割后。研究表明,機械損傷會誘導橡膠樹開割部位分化出大量次生乳管[20-21],因而改變了同化物分配的比例,大量的光合作用產物用來合成次生乳管,分配給樹木生長的同化物大量減少,從而使試割后莖圍生長速度明顯低于試割前。同時,試割時破壞了樹皮的篩管組織,影響了橡膠樹的光合作用產物的向下運輸,也會影響橡膠樹莖圍的生長。
子代測定可以估算親本無性系育種值,用于后向選擇可以作為原建種子園無性系留優去劣的依據[20]。橡膠樹極低的結實率[22-27]及橡膠樹遺傳基礎狹窄的現實[25]決定了橡膠樹無法像杜仲[28]、桉樹[29]、杉樹[30]、松樹[31]等種子量大的樹種一樣通過建立種子園進行良種選育。因此,選擇一個綜合性狀優良的親本同具有某個特定優異性狀的親本雜交,在子代中篩選更優單株是橡膠樹育種可行的途徑。
參考文獻
[1] 黃華孫, 梁茂寰, 吳云通, 等.中規模推廣級橡膠樹優良品種熱研7-33-97的選育[J]. 熱帶作物學報, 1994, 15(2): 1-6.
[2] 黃華孫, 方家林, 卓書蟬, 等. 橡膠樹優良品種熱研7-20-59的選育[J]. 熱帶作物學報, 2000, 21(2): 1-6.
[3] 伍思光, 劉作基, 馬 ?偉. 橡膠樹小規模推廣級無性系熱研7-18-55 的研究報告(I)[J]. 熱帶作物研究, 1995, 15(1): 8-10.
[4] 黃華孫, 吳云通, 方家林, 等. 橡膠樹優良品種熱研6-881的選育[J]. 熱帶作物研究, 1995, 15(3): 5-9.
[5] 李維國. 橡膠樹優良品種熱研7-33-97推廣應用前景[J]. 世界熱帶農業信息, 2008(11): 27-28.
[6] 周鐘毓. 我國巴西橡膠樹雜種后代早期選擇方法的研究概況[J]. 云南熱作科技, 1994, 17(3): 1-5
[7] 周鐘毓. 我國橡膠產量和抗性早期預測方法研究概況[J]. 熱帶作物研究, 1983(1): 8-15.
[8] 袁燮輝. 橡膠樹產量和抗寒、 抗風性能苗期葉片綜合測定[J]. 熱帶作物研究, 1984(4): 27-35.
[9] 方家林, 張曉飛, 李維國, 等. 橡膠樹野生種質59號雜交子代早期鑒定初報[J]. 熱帶農業科學, 2013, 33(8): 20-23.
[10] 李維國, 張曉飛, 黃 ?肖, 等. 3份橡膠樹野生種質雜交子代早期鑒定初報[J]. 熱帶作物學報, 2014, 35(10): 1 897-1 883.
[11] 黃華孫. 中國橡膠樹育種五十年[M]. 北京: 中國農業出版社, 2005.
[12] 李維國, 張偉算, 黃華孫. 橡膠樹優良無性系熱研5-11選育初報[J]. 熱帶作物學報, 2003, 24(3): 18-21.
[13] 張曉飛, 胡彥師, 黃華孫, 等. 橡膠樹優良親本RRIM600的研究與利用[J]. 熱帶農業科學, 2014, 34(6): 35-40.
[14] Paulo de Souza Goncalves. 巴西橡膠樹幾個苗期性狀的選擇及遺傳增益[J]. 田 ?郎, 譯. 世界熱帶農業信息, 2013(10): 1-3.
[15] 周鐘毓, 黃 ?香, 詹賽榮, 等. 橡膠樹乳管系與產量早期預測關系的研究[J]. 熱帶作物學報, 1984, 5(1): 29-36.
[16] 趙中奇. 橡膠樹乳管發育與產膠量關系研究[J]. 云南農業大學學報, 1998, 13(4): 58-61.
[17] 朱 ?軍. 遺傳學(第三版)[M]. 北京: 中國農業出版社, 2004.
[18] 徐晉麟, 陳 ?淳, 徐 ?沁. 現代遺傳學原理修訂本[M]. 北京: 科學出版社, 2004.
[19] 賈小明, 張廷楨, 張煥玲, 等.孟德爾群體數量性狀遺傳的概率論分析[J]. 西北林學院學報, 2014, 30(5): 106-110.
[20] Tian W M, Shi M j, Yu F Y, et al. Locaized effects of mechanical wounding and exogenous Jasmonic Acid on the induction of secondary laticifer differentiation in relation to the distribution of Jasmonic Acid in Hevea brasiliensis[J]. Acta Botanica Sinica, 2003, 45(11): 1 366-1 372.
[21]張世鑫, 劉世彪, 田維敏. 形成層活動對機械傷害誘導巴西橡膠樹次生乳管分化的影響[J]. 熱帶作物學報, 2011, 32(6): 1 037-1 041.
[22] 王明庥. 林木遺傳育種學[M]. 北京: 中國林業出版社, 2001.
[23] Rao B S. Pollination of hevea in Malaya[J]. J Rubb Res Inst Malaya, 1961(17): 14-18.
[24] Harihar G, Yeang H Y. The low fruit set that follows conventional hand pollination in Hevea brasiliensis: Insufficiency of pollen as a cause[J]. Rubb Res Inst Malaysia, 1984(32): 20-29.
[25] Omokhafe K O, Akpobome F A, Nasiru I. Diallel analysis of fruit set in Hevea brasiliensis Müll. Arg[J]. Genetics and Molecular Biology, 2007, 30(2): 428-430.
[26] Hamzah S, Chan J L, Yeang H Y. Pollen Tube growth and fruit-set success in Hevea brasiliensis hand-pollination influenced by the choice of clone and female flower[J]. Euphytica, 2002, 123: 1-8.
[27] 趙建文, 張曉飛, 翟琪麟, 等. 應用EST-SSR分析國外種質作為親本在中國橡膠樹雜交育種中的貢獻[J]. 熱帶作物學報, 2013, 34(2): 232-238.
[28] 魏永成. 杜仲雜交子代苗期表型性狀遺傳分析及選擇[D]. 楊凌: 西北農林科技大學, 2012.
[29] 陸釗華, 徐建民, 李光友, 等. 尾葉桉家系自由授粉子代性狀的分析[J]. 中南林業科技大學學報: 社會科學版, 2004, 24(5): 19-23.
[30] 鄭仁華, 蘇順德, 肖 ?暉, 等.杉木優樹多父本雜交子代測定及母本選擇[J]. 林業科學, 2014, 50(9): 44-50.
[31] 金國慶, 秦國峰, 劉偉宏, 等. 馬尾松生長性狀交配效應的遺傳分析及雜交組合選擇[J]. 林業科學, 2008, 44(6): 28-33.
