菠蘿蜜種質資源遺傳多樣性的SRAP分析
2015-05-30 10:48:04吳剛胡麗松黃麗芳郝朝運譚樂和
熱帶作物學報 2015年8期
吳剛 胡麗松 黃麗芳 郝朝運 譚樂和

中國熱帶農業科學院香料飲料研究所農業部香辛飲料作物遺傳資源利用重點實驗室 ?海南萬寧 ?571533
摘 ?要 ?利用SRAP標記技術,對46份菠蘿蜜種質資源進行遺傳多樣性研究。結果表明:20對SRAP引物組合共擴增出257條帶,其中多態性條帶194條,多態位點比率為75.5%,平均每對引物擴增條帶數和多態性條帶數分別為12.9條和9.7條。Shannon 遺傳多樣性指數I為0.365 7,Neis基因多樣性指數H為0.241 4,表明供試樣品遺傳多樣性不豐富。46份菠蘿蜜種質間的遺傳相似系數在0.674 8~0.975 5之間,平均為0.775 8,說明種質資源的親緣關系較近。通過UPGMA構建樹狀圖,當相似系數為0.771 0時,菠蘿蜜種質資源可分為6個類群,我國的種質資源和東南亞來源的種質相對分開聚類,來源地相同或較近的種質表現出了較為親密的親緣關系。
關鍵詞 ?菠蘿蜜;種質資源;遺傳多樣性;SRAP
中圖分類號 ?S667.8;Q943 ? ? ? ? ?文獻標識碼 ?A
Analysis of Genetic Diversity in
Artocarpus heterophyllus by SRAP
WU Gang, HU Lisong, HUANG Lifang, HAO Chaoyun, TAN Lehe*
Spice and Beverage Research Institute, CATAS/Key Laboratory of Genetic Resources Utilization of
Spice and Beverage Crops, Ministry of Agriculture,Wanning, Hainan 571533, China
Abstract ?In this study, Sequence Related Amplified Polymorphism(SRAP)technique was applied to analyze the genetic diversity of 46 Jackfruit accessions. A total of 257 bands were amplified by 20 selected primer combinations, in which 194 bands were polymorphic, the polymorphic percentage was 75.5%, with an average of 12.9 bands and 9.7 polymorphic bands for each primer combination. The Shannons genetic diversity information index(I)was 0.365 7 and the the Neis gene diversity(H)was 0.241 4, which revealed that the jackfruit accessions had low genetic diversity. The genetic similarity coefficients ranged from 0.674 8 to 0.975 5, with an average 0.775 8. Cluster analysis showed that 46 Jackfruit germplasm were divided into six groups on genetic similarity of 0.771 0. The phylogenetic dendrogram with UPGMA revealed that Chinese accessions and Southeast Asia from relative independent clusters, the germplasm resources coming from the same geographic area presented closer relationship in phylogeny.
Key words ?Artocarpus heterophyllus;Germplasm resources;Genetic diversity;SRAP
doi ?10.3969/j.issn.1000-2561.2015.08.004
菠蘿蜜(Artocarpus heterophyllus Lam.)又稱木菠蘿、樹菠蘿,為桑科(Moraceae)木菠蘿屬(Artocarpus)典型的熱帶果樹,原產于印度。在世界上菠蘿蜜多分布于東南亞國家,主產國為印度、孟加拉國及馬來西亞等。目前在我國海南、廣東、臺灣、福建、廣西和云南西雙版納以及四川南部的熱帶、南亞熱帶地區均有種植,其中海南省有近千年栽培歷史,為我國菠蘿蜜的主產區[1]。菠蘿蜜果實甜如蜜、香氣獨特、營養豐富,含有豐富的糖類、蛋白質、維生素和礦物質等;果實除生食外,還可制作脆片、糕點、飲料等,未成熟果可作各種菜肴的配料;菠蘿蜜樹體木質細密、色澤鮮黃、紋理美觀,是優良的家具和建筑用材[1-2]。自1999年,海南西聯農場從馬來西亞、泰國引進菠蘿蜜種質資源試種,并從中篩選出適合本地生產、綜合性狀表現較好的馬來西亞品種(系)后,近10余年來,菠蘿蜜生產發展迅速,種植面積以每年15%左右速度增長,目前種植面積約1.2萬hm2,年產量約15萬t[3]。隨著種植面積的快速增長,生產上因偽劣品種或品系混雜、同名異物、同物異名等造成損失的現象時有發生,菠蘿蜜品種(系)在苗期,形態上較相似,傳統的形態學鑒定不是很有效。
利用分子標記技術對菠蘿蜜種質資源進行準確、科學的鑒定和評價,可為資源保存和育種等工作提供可靠依據。國外,2001年Schnell等[4]利用AFLP標記技術對美國佛羅里達26份菠蘿蜜資源進行遺傳多樣性研究;2008年,印度學者[5]對來自印度不同區域資源進行AFLP多樣性分析。國內,葉春海等[6-8]采用RAPD、AFLP等分子標記技術對我國雷州半島的菠蘿蜜種質資源進行了遺傳多樣性分析,研究結果顯示供試種質的遺傳多樣性處于較低至中等水平,但涉及到菠蘿蜜主產區海南的資源量較少。以上研究采用的分子標記,如RAPD標記方法簡單、成本低,但重復性低;AFLP標記譜帶多、但分析程序復雜、成本高[9]。與其他標記相比, SRAP(Sequence related amplified polymorphism)標記具有操作簡單、重復性好、產率高、在基因組中分布均勻,便于克隆目標片段的特點,已經廣泛應用于枇杷、柑橘、黑芝麻、木薯、余甘子等[10-15]物種的遺傳多樣性分析,均表明SRAP 是一種能較好分析種質資源遺傳多樣性的分子標記技術之一。目前,利用SRAP分子標記技術對菠蘿蜜種質資源的研究尚未見報道。本研究利用SRAP 標記對中國熱帶農業科學院香料飲料研究所(以下簡稱香飲所)收集保存的46份菠蘿蜜種質資源進行遺傳多樣性分析,探討其親緣關系,為我國菠蘿蜜種質資源的鑒定、評價以及育種利用提供相關理論依據。
1 ?材料與方法
1.1 ?材料
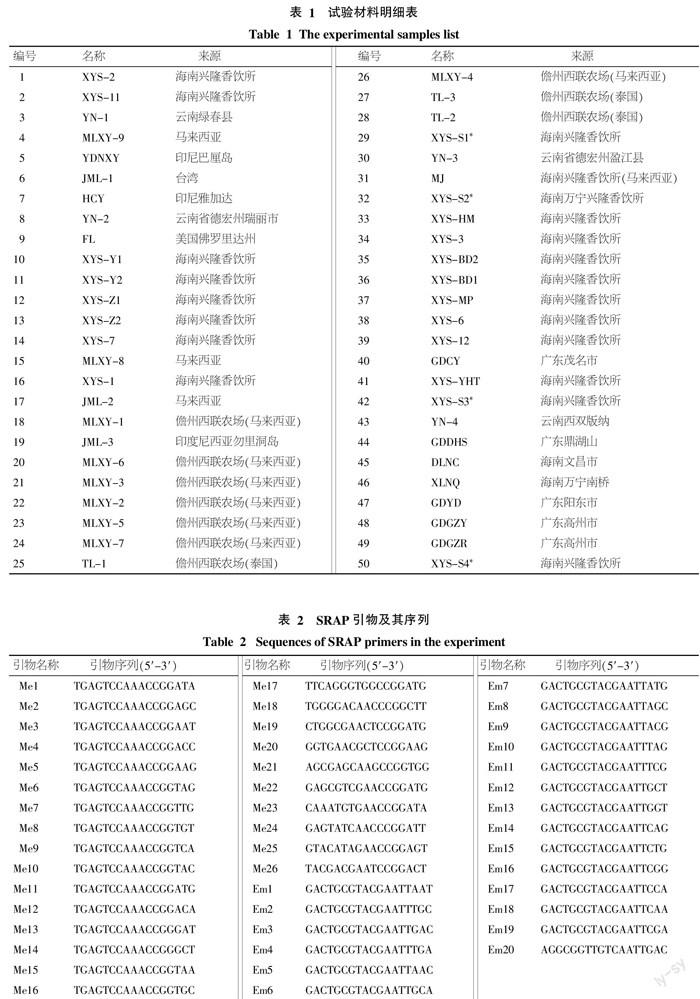
供試的50份材料均種植于香飲所保育基地,包括46份菠蘿蜜種質,其中海南種質22份、廣東5份、云南4份、馬來西亞種質9份、泰國3份、印尼2份以及美國1份;3份尖蜜拉種質(Artocarpus champeden Spreng.)和1份雜交種質(Artocarpus heterophyllus×Artocarpus champeden)。種質資源名稱中有“XYS-”種質為20世紀60~80年代,香飲所從海南省各地收集保存的優異資源,樹齡30 a以上,其余種質資源為近年收集保存的資源;種質名稱之后加“*”為濕胞種質;種質名稱JML-1(6)、 JML-2(17)JML-3(19)為尖蜜拉;種質名稱MJ(31)為雜交種。具體種質及其來源見表1。
1.2 ?方法
1.2.1 ?CTAB法提取DNA ? 取冷凍新鮮葉片0.3 g放入滅菌研缽中,加入液氮,充分研磨后,倒入離心管中,加入CTAB,65 ℃水中保溫1 h,不時搖勻。冷卻后離心,上清加入等體積氯仿:異戊醇溶液[24 ∶ 1(V/V)]抽提2次,取上清加1/100 RNase H,37 ℃靜置30 min,接著加2倍體積的無水乙醇沉淀,-20 ℃靜置1 h,離心后沉淀用70%乙醇洗2次。沉淀晾干后,加入50 mL ddH2O溶解,-20 ℃保存。電泳檢測并校定DNA濃度至50 ng/μL,備用。
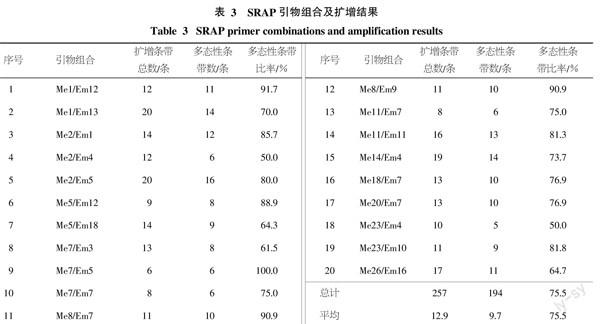
1.2.2 ?PCR擴增 ? SRAP引物由上海生工生物工程技術服務有限公司合成,引物序列參照齊蘭等[14],引物序列具體見表2。PCR擴增體系和程序參照鄭翠芳等[16]的方法并略加修改,PCR擴增體系為20 μL體系,含模板DNA 1μL、10×Buffer 2 μL、2.5 mmol/L dNTP Mixture 1.6 μL、正反向引物各1.2 μL、5 U Taq酶 0.2 μL、ddH2O 12.8 μL。PCR擴增程序為:94 ℃預變性5 min;94 ℃變性l min,35 ℃復性1 min,72 ℃延伸1 min,5個循環;94 ℃變性1 min,53 ℃復性1 min,72 ℃延伸1 min,35個循環;72 ℃延伸10 min。PCR擴增產物于4 ℃冰箱中保存。SRAP引物及其序列見表2。
1.2.3 ?電泳檢測 ? PCR擴增產物采用8%變性聚丙烯酰胺凝膠(210 g尿素、1 g甲叉、39 g丙烯酰胺、50 mL 10×TBE,定容至500 mL)進行電泳檢測;采用1×TBE作為電泳緩沖液;在20μL SRAP-PCR產物體系中加入6 μL溴酚藍上樣緩沖液(0.125 g溴酚藍、20 g蔗糖,ddH2O定容至50 mL),上樣1.5 μL至點樣孔中,120 V恒壓電泳1 h。
1.2.4 ?銀染顯影 ? 電泳結束后迅速進行銀染顯影。試驗步驟如下:①固定:將電泳完的凝膠放入固定液中(100 mL無水乙醇、5 mL乙酸,ddH2O定容至1 000 mL),固定5 min后取出,蒸餾水清洗1次。② 銀染:將固定后的凝膠放入銀染液中(0.8 g硝酸銀,ddH2O定容至1 000 mL),銀染10 min后取出,蒸餾水清洗2次。③顯影:將銀染后的凝膠放入顯影液中(15 g氫氧化鈉、10 mL甲醛,ddH2O定容至1 000 mL),直至條帶清晰顯出,取出凝膠于膠片燈箱上觀察拍照。
1.3 ?數據分析
條帶的記錄,根據SRAP指紋圖譜中Mark的分子量標準估計擴增條帶的分子量,每個供試樣品的擴增帶按有或無記錄,“有”賦值為1,“無”賦值為0,得到原始數據表征矩陣,弱帶及重復性不好的條帶不予統計。將圖形資料轉換成數據資料,計算單位引物擴增的條帶、多態性條帶及多態性條帶百分率。應用PopGen32軟件包計算46份菠蘿蜜種質資源的Shannon遺傳多樣性指數I,Neis基因多樣性指數H。用NTSYS pc2.10e分析軟件,采用UPGMA聚類方法,對菠蘿蜜種質資源進行分析,得到種質間的遺傳相似性,并建立樣品間的親緣關系樹狀圖。
2 ?結果與分析
2.1 ?SRAP標記多態性分析
用4份種質篩選520對SRAP引物組合,選用20對擴增多態性較好的引物組合對50份種質進行擴增分析。由于菠蘿蜜與尖蜜拉是同屬不同種的果樹,表3結果是基于46份菠蘿蜜種質資源擴增結果所統計,尖蜜拉種質以及兩種果樹的雜交種在遺傳相似性和聚類分析中與菠蘿蜜種質加以比較。
從表3可得出,20對引物組合對46份菠蘿蜜種質資源擴增,共產生257條帶,其中多態性DNA條帶194條,總多態性條帶比率為75.5%,每對引物平均擴增條帶為12.9條,引物Me7/Em5組合擴增的總條帶數(總位點數)最少6條,引物Me1/Em13和Me2/Em5等2個組合擴增的總條帶數最多,均為20條;平均每對引物組合擴增多態性條帶9.7條,范圍為6~16條,多態性條帶比率50.0%~100.0%。Shannon多樣性指數評價46份菠蘿蜜種質資源的遺傳多樣性,平均多樣性指數I為0.3657,Neis多樣性指數H為0.241 4,表明供試樣品遺傳多樣性不豐富。部分試驗材料的擴增結果見圖1。
2.2 ?遺傳相似性分析
通過對46份菠蘿蜜、3份尖蜜拉和1份雜交種,共50份種質的SRAP擴增結果(共擴增出286條DNA帶)進行計算其SM相似系數,46份樣品中有來自國內海南、云南和廣東以及國外馬來西亞、泰國、印尼和美國的種質資源。結果表明,46份菠蘿蜜種質之間的遺傳相似系數變化范圍在0.674 8~0.972 8之間,平均相似系數為0.775 8,種質資源的親緣關系較近,其中來自香飲所的2份種質XYS-Z1(12)和XYS-Z2(13)遺傳相似系數最高為0.975 5,其親緣關系最近;香飲所濕胞種質XYS-13(32)與馬來西亞的MLXY-6(20)、MLXY-3(21)資源遺傳相似系數都最低為0.674 8,親緣關系最遠。雜交種與46份菠蘿蜜種質的平均遺傳相似系數為0.747 1,雜交種與3份尖蜜拉種質的平均遺傳相似系數為0.698 1,表明雜交種與菠蘿蜜種質親緣關系較近。來自印尼、馬來西亞和臺灣的3份尖蜜拉種質平均相似系數為0.701 6,表明3份種質間存在一定的遺傳變異。
2.3 ?聚類分析
UPGMA法構建的聚類分析樹狀圖,在遺傳相似系數為0.745 0處時,46份菠蘿蜜種質資源和3份尖蜜拉及1份雜交種明顯區分開,其中菠蘿蜜種質歸為一大類,來自馬來西亞的JML-2(17)和雜交種(31)歸為一起,來自印尼JML-3(19)和臺灣JML-1(6)的尖蜜拉歸為一起;在遺傳相似系數為0.771 0處時菠蘿蜜種質又可分為6個亞類,見圖2。
圖2中,一亞類共19份資源:包括海南香飲所收集保存資源11份、海南萬寧南橋1份、廣東資源5份、云南資源2份;二亞類主要是來自香飲所收集保存的8份資源,分別是XYS-HM(33)、XYS-BD1(36)、XYS-MP(37)、XYS-BD2(35)、XYS-12(39)、XYS-YHT(41)、XYS-S3*(42)和XYS-3(34);三亞類包括4份資源,來自印尼的YDNXY(5)與HCY(7)、馬來西亞的MLXY-9(4)以及美國佛羅里達州FL(9); 四亞類包括11份資源,有8份種質資源來自馬來西亞,3份來自泰國,其中泰國資源先明顯聚在一起,再和其他馬來西亞資源聚為同一亞類;五亞類包括3份資源,來自云南德宏州瑞麗的YN-2(8)、德宏州盈江縣的YN-3(30)和來自海南文昌的DLNC(45);六亞類僅包含一份材料,香飲所收集的XYS-Y1(10)。
從圖2還可得出,一亞類19份資源二亞類8份資源先聚為一起,這27份資源都來自中國;三亞類4份資源四亞類11份資源也先聚為一起,這15份資源除了1份來自美國佛羅里達外,其余14份都來自泰國、馬來西亞和印尼等東南亞國家。此外從樹狀圖還可得出,4份濕胞種質XYS-S1*(29)、XYS-S2*(32)、XYS-S3*(42)、XYS-S4*(50)與其他干胞種質混雜聚類在一起。
3 ?討論與結論
本研究采用SRAP分子標記方法對46份菠蘿蜜種質資源DNA的遺傳多樣性進行檢測,20對引物組合共產生257條重復性好、穩定性高的條帶,且能把46份種質分開,說明了SRAP分子標記技術在菠蘿蜜品種資源鑒定中的可行性。利用分子標記技術可克服基于形態學、品種鑒定過程中出現的種種不足。
Shannon多樣性指數評價46份菠蘿蜜種質資源的遺傳多樣性,平均多樣性指數I為0.365 7,Neis多樣性指數H為0.241 4,表明供試樣品在DNA水平的遺傳多樣性低。遺傳相似系數變化范圍在0.674 8~0.975 5之間,平均遺傳相似系數為0.775 8,表明本研究中來自我國海南、廣東和云南的菠蘿蜜種質資源和來自泰國、馬來西亞、印尼等東南亞地區的種質資源在DNA水平上的遺傳相似度高,遺傳背景較狹窄。本研究采用的種質,以海南種質為主(22份),但研究結果與葉春海[6-7]對以雷州半島菠蘿蜜種質資源為主的遺傳多樣性研究結果類似,菠蘿蜜平均遺傳相似系數分別為0.734 1(RAPD)、0.775 0(ISSR),遺傳多樣性低、遺傳相似度高。表明目前我國收集、保存的菠蘿蜜資源遺傳多樣性不高,建議從菠蘿蜜原產地引進資源以豐富遺傳多樣性。此外,香飲所濕胞種質XYS-13(32)與馬來西亞來源的MLXY-6(20)、MLXY-3(21)資源遺傳相似系數都最低為0.674 8,親緣關系最遠,因而在選擇親本進行雜交組合育種時,選擇親緣關系遠的種質資源,更有利于雜交后代的遺傳性狀改良。
聚類分析時,選取尖蜜拉和雜交種作為外類群驗證,在遺傳相似系數為0.745 0處時,46份菠蘿蜜種質資源和3份尖蜜拉及1份雜交種明顯分開。從聚類圖中可看出,在遺傳相似系數為0.771 0處時,46份菠蘿蜜種質又可分為6個亞類,6個亞類種質資源聚類雖不能按地區來源單獨聚類,但大多數來源地相同或較近的種質表現了較為親密的親緣關系,呈現一定的地區性差異,如一亞類和二亞類的菠蘿蜜資源先聚類,主要包含我國海南、廣東和云南的資源;三亞類和四亞類等東南亞地區的資源也優先聚為一起(其中1份資源來自美國)。印度學者[5]對本國菠蘿蜜資源基于AFLP分子標記的聚類分析時也發現,資源聚類和來源地、形態性狀高度一致。本研究也發現一些較為特殊的個類,如六亞類僅包含一份材料,香飲所收集的XYS-Y1(10),說明該份資源有著較為特殊的遺傳背景。濕胞種質與其他干胞種質不能獨立聚類,SRAP分析的結果與前人利用ISSR和RAPD標記得到結果類似,均不能區分濕胞、干胞種質[6-7]。這可能與所用SRAP引物有限,沒有擴增出濕胞、干胞的特異位點有關。
豐富的遺傳多樣性是物種適應外界環境變化的物質基礎,農業生產上應用的作物品種類型愈豐富愈能抵御變化莫測的自然災害。綜合Shannon多樣性指數、Neis多樣性指數和遺傳一致性結果得出,供試菠蘿蜜種質資源遺傳多樣性較低,在資源保存和遺傳育種中需要進一步拓寬資源多樣性和遺傳背景,如引進來自印度的資源或者菠蘿蜜屬的野生近緣種,并進一步加強資源研究的廣度和深度,將利于促進我國菠蘿蜜種質資源的創新和新品種選育。
參考文獻
[1] 譚樂和, 劉愛勤, 林民富. 菠蘿蜜種植與加工技術[M]. 北京: 中國農業出版社, 2007.
[2] 譚樂和, 吳 ?剛, 劉愛勤,等. 菠蘿蜜高效生產技術[M]. 北京: 中國農業出版社, 2012.
[3] 吳 ?剛, 陳海平, 桑利偉, 等. 中國菠蘿蜜產業發展現狀及對策[J]. 熱帶農業科學, 2013, 33(2): 91-97.
[4] Schnell R J, Olano C T, Campbell R J, et al. AFLP analysis of genetic diversity within a jackfruit germplasm collection[J]. Scientia Horticulaturae, 2001, 91: 261-274.
[5] Shyamalamma S, Chandra S B C, Hegde M, et al. Evaluation of genetic diversity in Jackfiuit(Artocarpus heterophyllus Lam.)based on amplified fragment length polymorphism markers[J]. Genet Mol Res, 2008, 7(3): 645-656.
[6] 葉春海,李映志,豐 ?鋒. 雷州半島菠蘿蜜種質遺傳多樣性的RAPD分析[J]. 園藝學報, 2005, 32(6):1 088-1 091.
[7] 葉春海, 王耀輝, 李映志, 等. 菠蘿蜜遺傳多樣性的ISSR分析[J]. 果樹學報, 2009, 26(5): 659-665.
[8] LI Yingzhi, MAO Qi, FENG Feng, et al. Genetic diversity within a Jackfruit(Artocarpus heterophyllus Lam.) germplasm collection in China using AFLP markers[J]. Agricultural Sciences in China. 2010, 9(9): 1 263-1 270.
[9] 柳李旺, 龔義勤, 黃 ?浩, 等. 新型分子標記-SRAP與TRAP及其應用[J]. 遺傳, 2004, 26(5): 777-781.
[10] Li G, Quiros C F. Sequence-related amplified polymorphism(SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica[J]. Theoretical and Applied Genetics, 2001, 103(2-3): 455-461.
[11] 喬燕春, 林順權, 劉成明, 等. SRAP分析體系的優化及在枇杷種質資源研究上的應用[J]. 果樹學報, 2008, 25(3): 348-352.
[12] Uzun T, Yesiloglu Y, Aka-Kacar, et al. Gulsen Genetic diversity and relationships within Citrus and related genera based on sequence related amplified polymorphism markers(SRAPs)[J]. Sci Hort, 2009, 121(3): 306-312.
[13] 車 ?卓, 張艷欣, 孫 ?建, 等. 應用SRAP 標記分析黑芝麻核心種質遺傳多樣性[J]. 作物學報, 2009, 35(10): 1 ?936-1 941.
[14] 齊 ?蘭, 王文泉, 張振文, 等. 木薯種質資源的遺傳多樣性分析與評價[J]. 熱帶作物學報, 2010, 31(10): 1 661-1 668.
[15] 郭林榕, 周 ?平, 陳志峰. 22份余甘子核心種質資源遺傳多樣性的SRAP分析[J]. 熱帶作物學報, 2014, 35(7): 1 382-1 387.
[16] 鄭翠芳, 朱曉東, 方麗娜,等. 愛玉子性別與品系的SRAP分析[J]. 熱帶作物學報, 2009, 30(12): 1 740-1 745.
