2種藥用植物在喀斯特生境下的光合特征及適生性
2016-05-03 15:26:21王瑞吳沿友邢德科杭紅濤
江蘇農業科學 2016年3期
王瑞+吳沿友+邢德科+杭紅濤

摘要: 為篩選喀斯特生態修復植物,探討植物配置方案,利用Li-6400便攜式光合測定系統分別測定了莽草、長刺楤木2種藥用植物葉片的光合作用日變化特征,并結合植物葉片各生理生態因子對凈光合速率(Pn)影響的通徑分析、葉綠素含量、碳酸酐酶活力和穩定碳δ13C同位素組成對比了2種植物在喀斯特生境下的適生能力。結果表明:長刺楤木的大多數光合作用指標(Pn、LUE、Cond、Ci、Tr)均高于莽草,且具有較高的抗脅迫能力和碳酸酐酶活力;2種藥用植物在相同喀斯特生境下表現出不同的適應機制,長刺楤木的喀斯特適生性高于莽草,但莽草的抗干旱能力強于長刺楤木。
關鍵詞: 喀斯特;莽草;長刺楤木;適生性;光合生理特征
中圖分類號: S567.901 文獻標志碼: A 文章編號:1002-1302(2016)03-0216-04
生態修復是治理喀斯特石漠化地區的有效途徑[1-2],而合理的植物種群、群落構建是該領域的共性關鍵技術[3],因此喀斯特地區適生植物的選擇就顯得至關重要。喀斯特地貌因存在石漠化、干旱溶巖、土地瘠薄、高pH值、高鈣、高重碳酸鹽及高鈣鎂濃度等限制因素而被認定為一種特殊的脆弱生境,嚴重影響植物的生長發育[4]。但有研究表明,部分植物可借助體內的碳酸酐酶調節控制每一個與氣孔開閉有關的保衛細胞,且可在碳酸酐酶作用下利用無機碳源進行光合作用[5]。近年來研究表明,喀斯特適生植物可交替使用大氣中的CO2和土壤中的重碳酸鹽,經碳酸酐酶的催化作用進行光合作用而正常生長,且碳酸酐酶活力可作為植物喀斯特適生性的有力判據[6-9]。高適生性喀斯特先鋒植物的合理配置不僅可有效發揮喀斯特生態修復效用,還在低成本固碳增匯方面具有相當優勢[10]。在選擇、配置喀斯特生態修復作物的同時,若能兼顧經濟價值和碳匯效應,可有效激發種植者的積極性和環保效益,對修復和改善喀斯特生態系統具有促進意義。
長刺楤木(Aralia spinifolia Merr)、莽草(Illicium lanceolatum A.C.Smith)在貴州喀斯特山區廣泛分布,且資源豐富[11-12],具有一定藥用價值。長刺楤木根、根皮主要含皂苷類化合物,可供藥用,具有駁骨、拔毒之功效,具有治頭昏、頭痛、風濕痹痛、跌打損傷、蛇傷,以及強壯、免疫促進、抗腫瘤、抗潰瘍和保肝等作用[13]。莽草因含有seco-prezizaane類型的倍半菇類化合物而具有殺蟲、抗氧化、抗菌、鎮痛、抗腫瘤等多種藥理活性,有研究者正嘗試從莽草中尋找高效、安全、依賴性小的抗炎、鎮痛新藥先導化合物[12]。
筆者所在課題組在喀斯特生態修復植物篩選、配置的野外試驗中發現,長刺楤木、莽草在貴州喀斯特山區長勢良好,資源豐富。本研究考察野生長刺楤木、莽草在喀斯特生境下的光合作用特征參數,并通過穩定性碳同位素技術、碳酸酐酶活力對這2種藥用植物的喀斯特適生性進行比較,以期為喀斯特生態修復的適生植物篩選和配置提供科學依據。
1 材料與方法
1.1 研究區域概況、材料與處理
研究區域為貴州省六盤水市玉舍國家森林公園(喀斯特原始森林),地理位置為104°47′813″~104°50′385″E、26°27′508″~26°27′881″N,總面積為3 342.27 hm2,海拔 1 700~2 503 m,屬于典型的亞熱帶季風氣候,立體氣候較為明顯。土壤類型主要為由玄武巖、石灰巖等發育的山地黃壤土,相關指標為銨態氮含量(11.01±2.2) mg/L,有效鉀含量(29.67±5.64) mg/L,速效鉀含量(31.53±5.64) mg/L,pH值5.24±0.19,含鹽量(0.03±0.26)%,有機質含量(6.43±2.41)%。年平均溫度為12 ℃左右,最低溫度為1月份的 3 ℃,最高溫度為7月份的22 ℃,年平均降水量為 1 380 mm;雨熱同季,濕度大,降水主要集中在4—10月;植被豐富,具有近700種藥用種子植物,是目前六盤水市保護較好的原始喀斯特天然森林植被[14-15]。本試驗測定該區域內具有代表性的植株葉片光合作用日變化特征參數;采集相應葉片封存于液氮中,帶回實驗室測定相應植物葉片的碳酸酐酶活力、葉綠素含量和穩定碳同位素δ13C值。
1.2 光合作用日變化特征參數測定
光合作用日變化特征參數于2012年5月13日(晴朗)測定,分別從試驗植株的第4張葉(從展開心葉向下數)開始,選取健康、完全展開葉進行測定(測定時用緩沖瓶使CO2濃度相對穩定,每張葉測3次,取平均值)。每隔2 h用Li-6400光合儀連續測定光合作用日變化特征參數,具體包括凈光合速率[Pn,μmol/(m2·s)]、蒸騰速率[Tr,mmol /(m2·s)]、氣孔導度[Cond,mmol/(m2·s)]、胞間CO2濃度(Ci,μmol/mol)、大氣CO2濃度(Ca,μmol/mol)、光合有效輻射[Par,μmol/(m2·s)]、大氣溫度(Ta, ℃)、大氣相對濕度(RH,%)等特征參數,并計算出葉片光能利用率(LUE,即Pn/Par,%)、瞬時水分利用效率(WUE,即Pn/Tr,以CO2計,μmol/mmol)。
1.3 葉綠素熒光參數測定
當日光合有效輻射達到并保持0 μmol/(m2·s) 1 h后,用美國Li-6400-40便攜式熒光儀及配套的Li-6400-40熒光葉室進行葉綠素熒光測定。按van Kooten等方法[16]測定相應熒光參數指標:初始熒光(Fo)、最大熒光(Fm),并計算出PSⅡ最大光化學效率(Fv/Fm)=(Fm-Fo)/Fm及PSⅡ潛在活性(Fm-Fo)/Fo。
1.4 碳酸酐酶活力測定
碳酸酐酶活力采用銻微電極法進行測定[17-18],選擇長勢一致用于光合作用日變化特征參數測定的葉片,立即封存于液氮中帶回試驗室。取0.3~1.0 g植物葉片,放入預冷的研缽中,迅速加入液氮,再加入3mL巴比妥緩沖液(10 mmol/L,含巰基乙醇50 mmol/L,pH值8.3)進行研磨,取研磨液倒入5 mL離心管中,將離心管冰浴20 min后,在 13 000 r/min 下離心5 min,取上清液,冷藏待測。樣品檢測過程中保持反應系統溫度在0~2 ℃,取待測上清液50~1 000 μL,加入15 mL 巴比妥緩沖液中,然后迅速加入10 mL預冷的(0~2 ℃)的飽和CO2蒸餾水,用pH電極監測反應體系pH值變化,記下pH值下降1個單位(如pH值從8.2~7.2)所需的時間,記為t;同時記錄在酶失活條件下pH值下降1個單位所需的時間,記為t0;酶的活力用WA-unit表示(WA=t/t0-1)。
1.5 葉綠素含量的測定
植物葉片(液氮保存)葉綠素含量采用分光光度法測定[19]。
1.6 穩定碳同位素比值(δ13C)的測定
將采回的植物葉片經108 ℃殺青,再于80 ℃下烘48 h,粉碎均勻并過篩,稱取3~5 mg樣品裝入石英管中,再裝入經高溫處理的氧化銅絲、鉑絲(催化劑),將石英管在高溫下煅燒(850 ℃保溫3~4 h),確保樣品中的碳全部轉化成氣態的CO2。稱取一定量轉化成供質譜儀分析的二氧化碳氣體,在MAT-252質譜儀上進行測定(中國科學院地球化學研究所),測量精度范圍為±0.1%,分析結果以PDB(Pee Dee Belemnite)表示,即δ13CPDB=[(13C/12C)樣本-(13C/12C)標準]/(13C/12C)標準×1 000‰。
1.7 數據處理
試驗數據采用Microsoft Excel 2007和SPSS 13.0統計軟件進行統計分析。
2 結果與分析
2.1 2種植物的光合參數日變化
2.1.1 凈光合速率和光能利用率日變化特征 由圖1可見,莽草、長刺楤木葉片的凈光合速率變化趨勢大致相似,呈單峰趨勢。2種植物葉片的Pn值都是從08:00至10:00呈下降趨勢,而10:00后開始上升,14:00達到峰值[莽草:7.48 μmol/(m2·s)、長刺楤木:14.71 μmol/(m2·s)];2種植物的Pn值均在18:00達到最低值,分別為-1.65、0.03 μmol/(m2·s)。莽草、長刺楤木的日均Pn值表現為長刺楤木[(9.21±1.21) μmol/(m2·s)]>莽草[(4.31±0.69) μmol/(m2·s)],可見莽草的光合生產力高于長刺楤木。在光能利用效率方面,2種植物均表現出早晚高、中午低的變化趨勢,2種藥用植物光能利用效率峰值均在08:00達到最高值,10:00達到最低值;由2種植物葉片的日均LUE值[長刺楤木(6.15%)、莽草(3.19%)]可以推斷出長刺楤木對于光照的利用能力高于莽草。
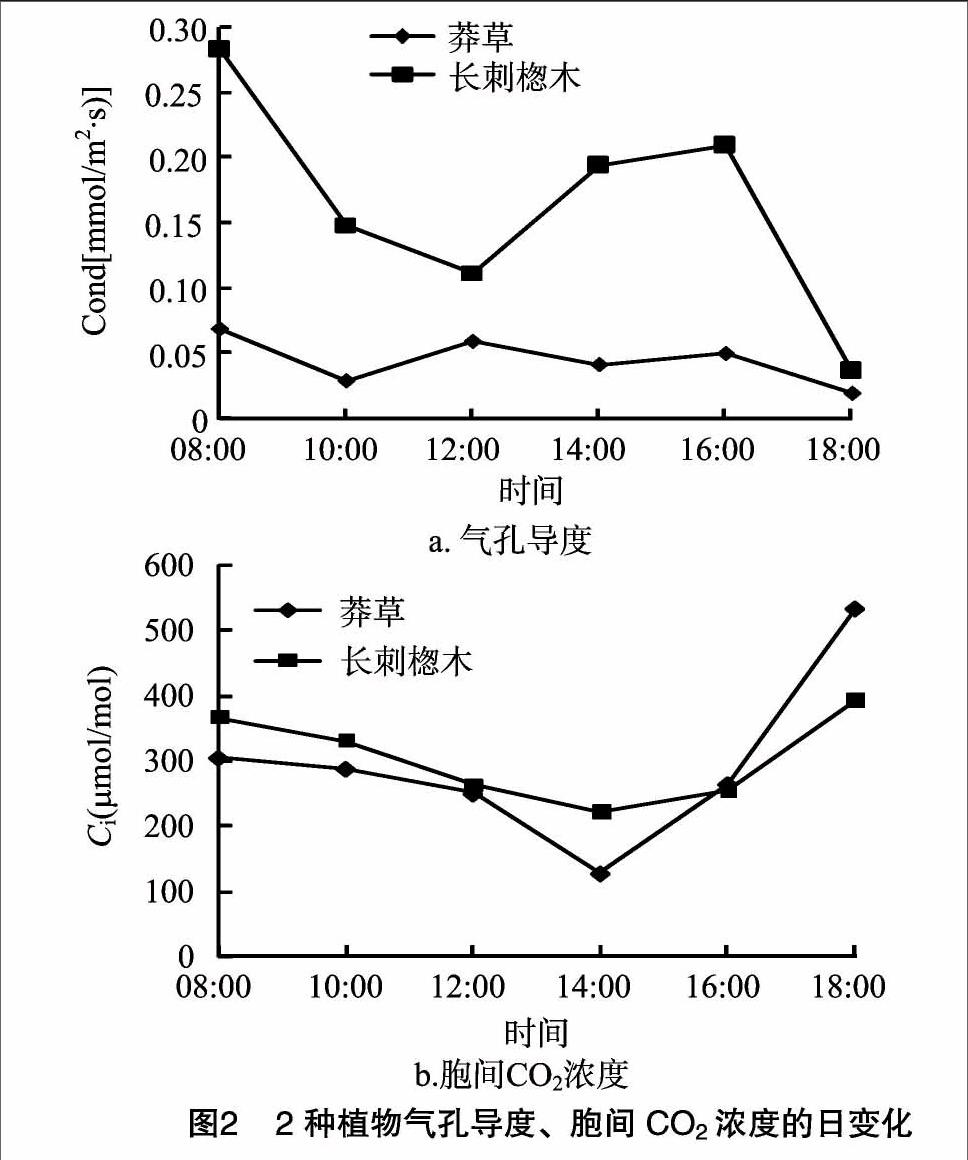
2.1.2 氣孔導度和胞間CO2濃度的日變化特征 葉面上的氣孔在控制CO2吸收和水分損失的平衡中起著關鍵作用[20]。由圖2可見,08:00時2種植物葉片Cond值最高[長刺楤木:0.28 mmol/(m2·s),莽草:0.07 mmol/(m2·s)];為適應光照度增強和溫度升高,2種植物的氣孔導度發生不同的變化:莽草呈現以4 h為周期的波動,且受外界影響較小,而長刺楤木沒有表現出明顯規律;另外,長刺楤木變化曲線的波動明顯強于莽草,說明長刺楤木的氣孔導度更易受環境因子影響。2種植物葉片Ci值均表現出早晚高、中午低的變化趨勢。08:00—14:00,Ci值呈現出與凈光合速率相反的變化趨勢,說明該時間段內隨著2種植物葉片光合作用的增強,CO2消耗量逐漸增大,Ci降低,14:00達到峰底(長刺楤木:220.19 μmol/mol,莽草:126.91 μmol/mol),而后隨著Pn的下降而回升。
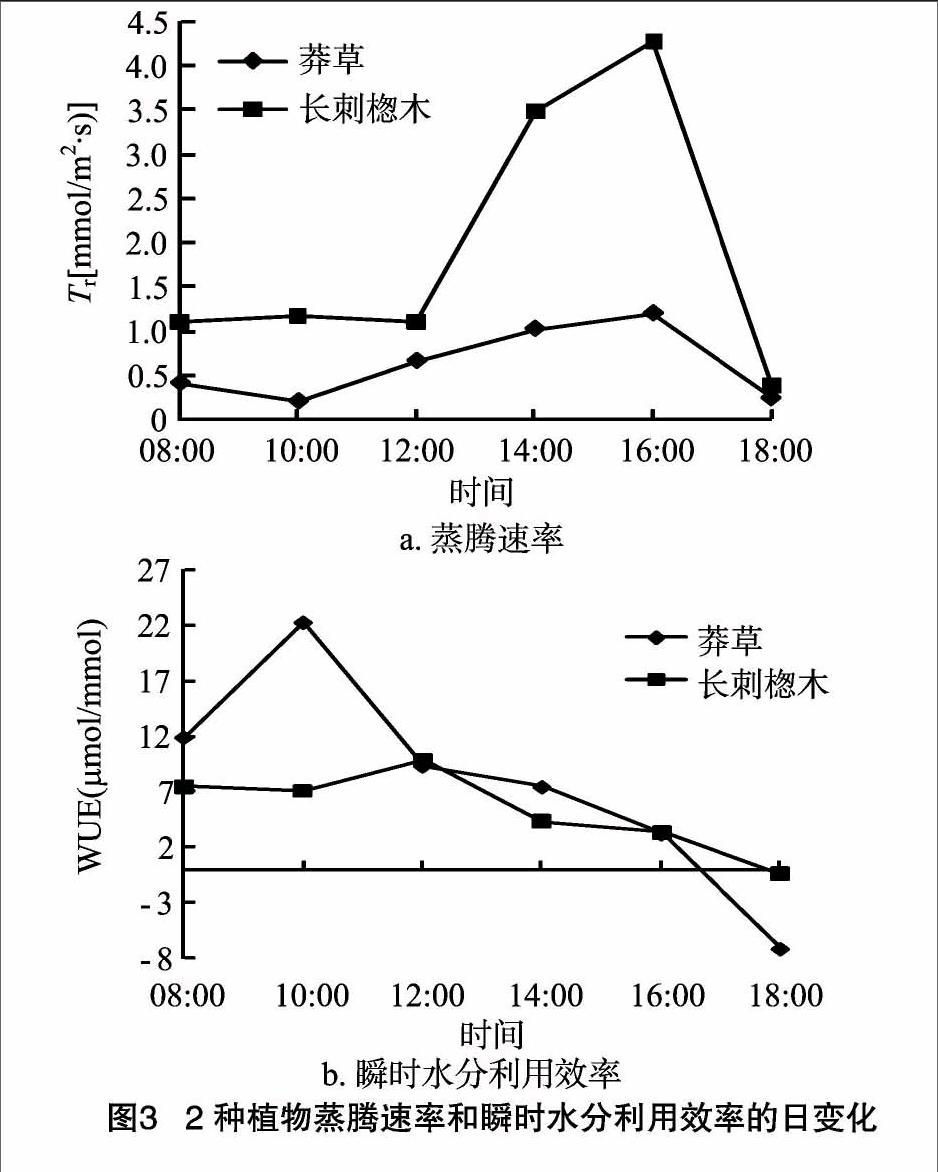
2.1.3 蒸騰速率和瞬時水分利用效率的日變化特征 由圖3可見,莽草與長刺楤木的Tr日變化大致呈先上升后下降的趨勢(與Pn變化趨勢一致), 測定時間內2種植物Tr峰值均出現在16:00[莽草:1.19 mmol/(m2·s),長刺楤木:4.27 mmol/(m2·s)];從圖3還可看出,莽草的Tr日變化較為平緩。2種植物的WUE平均值大小為莽草[(7.80±2.28) μmol/mmol]>長刺楤木[(5.21±0.86)μmol/mmol],說明莽草的瞬時水分利用效率高于長刺楤木。
2.1.4 2種植物葉片Pn與各項生理生態因子的關系 由表1、表2莽草、長刺楤木2種植物葉片的凈光合速率與生理生態因子的通徑分析和相關分析可以看出,在生態因子中,莽草、長刺楤木的Ca、Ta與Pn均無顯著相關,其中Ca與Pn呈負相關;在2種植物的各生理因子中,莽草葉片的Ci與Pn呈極顯著負相關。通徑分析結果(總效應絕對值)表明,各生態因子對Pn直接作用大小為Par>RH>Ca>Ta。Par對莽草葉片Pn的最大直接效應除源于自身作用外,主要通過Ci的間接作用影響Pn,且對Pn呈最大總效應。在生理因子中,對莽草葉片Pn影響大小為Ci>Cond>Tr。對于長刺楤木而言,各生態因子對其Pn影響大小為Par>RH>Ca>Ta,而各生理因子對其Pn影響排序為Tr>Cond>Ci。
通過分析可見,對莽草Pn生態因子的決策系數(R2)大小為Par>Ta>RH>Ca,而對長刺楤木的大小也為Par>RH>Ca>Ta。不同的是對莽草Pn日變化規律的決定生態因子為Par,而長刺楤木為Par、RH,其余為限制因子。在生理因子方面,對莽草Pn生態因子的決策系數(R2)大小為Ci>Cond>Tr,而對長刺楤木大小為Tr>Ci>Cond。由表1可見,影響莽草Pn日變化規律的決定生理因子為Ci、Cond,限制因子為Tr;而對長刺楤木的決定生理因子為Tr,限制因子為Cond、Ci。
2.2 植物葉片葉綠素含量與葉綠素熒光參數
研究表明,葉綠素含量的高低能在一定程度上反映葉片的光合能力,直接影響植物的生長;而葉綠素熒光可與光合特性、水分代謝特性互補,用于揭示植物對環境生態適應性機制[21]。本研究根據相關文獻報道方法[19],測得莽草、長刺楤木2種植物葉片的葉綠素含量。由表3可見,2種植物葉片葉綠素含量大小為長刺楤木>莽草,結合上述Pn、LUE檢測結果可說明,長刺楤木對光能利用的能力高于莽草。
葉綠素熒光技術作為一種無損光合作用研究的探針,幾乎可以反映植物葉片所有的光合作用過程。近年來,該技術在光合作用、植物脅迫生理學、水生生物學、海洋學和遙感等方面得到了廣泛的應用[22]。由表3可見,長刺楤木的PSⅡ最大光化學效率(Fv/Fm)大于莽草,結合PSⅡ潛在活性(Fv/Fo)說明,長刺楤木相對于莽草表現出較高的潛在光合能力,由此可推測,在喀斯特生境下長刺楤木相對莽草具有較高的抗脅迫能力。
2.3 植物葉片的碳酸酐酶活力與δ13C 值
研究證明,具有高碳酸酐酶活力的喀斯特適生植物在遭受脅迫時,可利用碳酸酐酶的催化作用將細胞內的碳酸氫根離子轉化成CO2、H2O,以彌補水分、CO2 的不足;可交替使用大氣中的CO2和土壤中的無機碳進行光合作用而起到固碳增匯的目的[6,10];此外,植物根系分泌的高濃度碳酸酐酶可催化土壤中CO2、H2O轉換成HCO3-、H+,以加速石灰巖溶解、提高成土速率[23]。根據文獻[17]報道方法測得2種植物葉片的碳酸酐酶活力,兩者大小為長刺楤木[(11 161.51±1 159.2) WAU/g]>莽草[(876.58±62.87) WAU/g]。由此可推斷,在喀斯特生境下,長刺楤木的固碳增匯能力高于莽草,且具有更好的喀斯特生態修復效用。
基于穩定碳同位素的強烈分餾特征,穩定碳同位素技術被作為一種衡量植物長期水分利用效率的有效工具,即植物的δ13C值與其長期水分利用效率呈正相關[24]。研究表明,2種植物葉片的δ13C大小為莽草的(-26.931±0.015)‰>長刺楤木的(-28.82±0.004)‰,即莽草的長期水分利用效率高于長刺楤木,與上述WUE結果一致。通過對上述各指標的綜合分析,可以將長刺楤木的低水分利用效率歸屬于該植物葉片的高Pn、Tr、Cond。因此,莽草較長刺楤木更能適應干旱環境。
3 討論與結論
本研究通過光合作用日變化特征參數、碳酸酐酶活力和穩定性碳同位素組成的分析對莽草、長刺楤木的喀斯特適生性進行比較和討論。在光合日變化特性方面,長刺楤木葉片的Pn、LUE、葉綠素含量均高于莽草,說明長刺楤木的作物生產力較高。經通徑分析和相關分析可以看出,對莽草Pn日變化規律的決定生態因子為Par,而長刺楤木為Par、RH;莽草Pn日變化規律的決定生理因子為Ci、Cond,而長刺楤木決定生理因子為Tr。這說明不同植物即使在相同的環境條件下,也會表現出不同的適應機制。通過對比發現,長刺楤木葉綠素含量、葉綠素熒光和PSⅡ最大光化學效率均高于莽草,結合碳酸酐酶活力比較,長刺楤木[(11 161.51±1 159.2) WAU/g]>莽草[(876.58±62.87) WAU/g]可以推測,長刺楤木的喀斯特適生性高于莽草,且固碳增匯能力高于莽草,具有更好的喀斯特生態修復效用。但是莽草具有高于長刺楤木的WUE,結合穩定碳同位素分析結果可知,莽草較長刺楤木更能適應干旱環境。
參考文獻:
[1]Wei Y,Yu L F,Zhang J C,et al. Relationship between vegetation restoration and soil microbial characteristics in degraded karst regions:a case study[J]. Pedosphere,2011,21(1):132-138.
[2]Zhu H H,He X Y,Wang K L,et al. Interactions of vegetation succession,soil bio-chemical properties and microbial communities in a Karst ecosystem[J]. European Journal of Soil Biology,2012,51(5):1-7.
[3]郭 柯,劉長成,董 鳴. 我國西南喀斯特植物生態適應性與石漠化治理[J]. 植物生態學報,2011,35(10):991-999.
[4]Yan J,Li J,Ye Q,et al. Concentrations and exports of solutes from surface runoff in Houzhai Karst Basin,southwest China[J]. Chemical Geology,2012,304/305(3):1-9.
[5]Hu H H,Boisson-Dernier A,Israelsson-Nordstroem M,et al. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells[J]. Nature Cell Biology,2010,12(1):87-U234.
[6]吳沿友,邢德科,劉 瑩. 植物利用碳酸氫根離子的特征分析[J]. 地球與環境,2011,39(2):273-277.
[7]吳沿友,劉從強,李萍萍,周文美,王寶利. 利用碳酸酐酶活力鑒定喀斯特適生植物的方法:中國,CN101067148[P]. 2011-05-20.
[8]Xing D,Wu Y. Photosynthetic response of three climber plant species to osmotic stress induced by polyethylene glycol(PEG)6000[J]. Acta Physiologiae Plantarum,2012,34(5):1659-1668.
[9]Wu Y Y,Liu C Q,Li P P,et al. Photosynthetic characteristics involved in adaptability to Karst soil and alien invasion of paper mulberry (Broussonetia papyrifera L. Vent) in comparison with mulberry (Morus alba L.)[J]. Photosynthetica,2009,47(1):155-160.
[10]吳沿友. 喀斯特適生植物固碳增匯策略[J]. 中國巖溶,2011,30(4):461-465.
[11]楊碧仙. 貴州省五加科藥用植物的生境和地理分布研究[J]. 時珍國醫國藥,2010,21(6):1505-1506.
[12]梁 婕. 莽草的化學成分和生物活性研究[D]. 福州:福建中醫藥大學,2011.
[13]劉軍民,丁 平,徐鴻華,等. 長刺楤木的生藥學研究[J]. 廣州中醫藥大學學報,2000,17(2):173-175,194.
[14]左經會,林長松,田應洲.貴州玉舍國家森林公園種子植物區系研究[J]. 廣西植物,2006,26(4):434-440.
[15]向 紅,左經會,林長松,等. 貴州玉舍國家森林公園藥用種子植物資源調查[J]. 北方園藝,2010(17):79-82.
[16]Van Kooten O,Snel J F. The use of chlorophyll fluorescence nomenclature in plant stress physiology[J]. Photosynthesis Research,1990,25(3):147-150.
[17]吳沿友 施,王 坤,李萍萍,等. 一種利用銻微電極測定植物碳酸酐酶活力的電化學方法:中國,CN101793860A[P]. 2010-08-15.
[18]Wu Y Y,Shi Q Q,Wang K,et al. An electrochemical approach coupled with Sb microelectrode to determine the activities of carbonic anhydrase in the plant leaves[J]. Lecture Notes in Electrical Engineering,2011,86(1):87-94.
[19]李合生,孫 群,趙世杰,等. 植物生理生化實驗技術原理[M]. 北京:高等教育出版社,2000:248-250.
[20]Sun C X,Qi H,Hao J J,et al. Single leaves photosynthetic characteristics of two insect-resistant transgenic cotton (Gossypium hirsutum L.) varieties in response to light[J]. Photosynthetica,2009,47(3):399-408.
[21]Li X G,Meng Q W,Zhao S J. Photoinhibition and photoprotection mechanisms in Ginkgo biloba leaves under strong light stress[J]. Scientia Silvae Sinicae,2004,40(3):56-59.
[22]Gorbe E,Calatayud A. Applications of chlorophyll fluorescence imaging technique in horticultural research:a review[J]. Scientia Horticulturae,2012,138(1):24-35.
[23]李 強,何媛媛,曹建華,等. 植物碳酸酐酶對巖溶作用的影響及其生態效應[J]. 生態環境學報,2011,20(12):1867-1871.
[24]Scartazza A,Lauteri M,Guido M C,et al. Carbon isotope discrimination in leaf and stem sugars,water-use efficiency and mesophyll conductance during different developmental stages in rice subjected to drought[J]. Australian Journal of Plant Physiology,1998,25(4):489-498.
