水分脅迫對美國紅楓幼苗生長及葉色變化的影響
2016-05-03 15:31:15張鑫肖婷婷李艱王玉濤劉廣林
江蘇農業科學 2016年3期
張鑫+肖婷婷+李艱+王玉濤+劉廣林

摘要: 為研究不同水分脅迫處理對優良景觀觀賞樹種美國紅楓幼苗生長及葉色變化的影響,采用1年生改良美國紅楓Acer×freem anii ‘Autumn Blaze扦插苗為試驗材料,采用盆栽試驗的方法,設4個水分梯度處理,土壤水分含量分別為土壤田間持水量的35%(重度干旱組)、50%(中度干旱組)、65%(輕度干旱組)和80%(對照組)。研究結果表明:本試驗中,在幼苗生長方面,土壤水分含量為土壤田間持水量的80%時,最有利于美國紅楓幼苗生長,株高、地徑分別達到63.43、0.91 cm。其次是土壤水分含量為土壤田間持水量的65%時,株高、地徑分別為62.18、0.89 cm。在幼苗葉色變化方面,土壤水分含量為土壤田間持水量的50%時,最有利于葉片中花青素的積累。而土壤水分含量為土壤田間持水量的80%時,葉片最早變紅且鮮艷,但變化較為緩慢。因此,土壤水分含量在本研究的范圍內時,土壤田間持水量80%的水分含量最有利于美國紅楓幼苗的生長及變色。
關鍵詞: 美國紅楓;水分脅迫;幼苗生長;葉色變化
中圖分類號: S687.101 文獻標志碼: A 文章編號:1002-1302(2016)03-0224-04
美國紅楓(Acer rubrum)屬槭樹科槭樹屬多年生落葉喬木,又名紅花槭、北美紅楓等,原產于美國北部以及加拿大大部分地區。樹高可達30 m,葉掌狀3~5裂,耐寒、耐旱、耐濕且抗污染,適應性強,生長迅速。入秋葉紅色,為歐美典型的秋色葉樹種,又因樹冠整潔,被廣泛用于園林造景以及行道樹栽種。目前我國華北、東北、西北及華東等地都有栽培。
隨著水資源危機和干旱化危害的不斷加劇,植物如何適應干旱已成為全球研究的重要問題之一[1]。園林綠化的節水灌溉已成為城市節水的重要途徑之一。現在好多關于水分的研究都是針對于農作物的產量,而研究彩葉樹種的水分生理特別是干旱脅迫下的生長及葉色變化,對于園林建設的節水灌溉及提高園林植物的觀賞特性具有重要意義。侯舒婷[2]在研究黃金香柳對水分脅迫的響應時發現,隨著干旱程度的加劇,黃金香柳各指標的生長均受到顯著抑制;楊躍文[3]的研究表明,干旱脅迫使北沙柳的生長受到抑制,株高與基徑的生長減緩,總生物量的積累受到抑制。Sherwin等[4]在研究花青苷對復蘇植物的光保護作用時,發現干旱脅迫能夠增加復蘇植物和黃櫨葉片中花青苷含量;孔艷菊[5]的研究表明,黃櫨葉片中的葉綠素含量隨干旱脅迫程度的加劇呈上升趨勢。曹晶[6]的研究表明,夏、秋干旱和水淹脅迫均使紅葉石楠葉片中葉綠素含量降低,花青苷含量升高。彩葉樹木葉片呈色是各種色素綜合作用的結果,以上研究表明水分脅迫下能夠引起彩葉樹木葉片中的色素含量發生變化,從而引起葉片呈色的變化。而目前還沒有有關水分脅迫對美國紅楓幼苗的生長及葉色變化影響的研究。本試驗通過不同的水分處理,研究其對植株幼苗生長及葉色變化的影響,掌握其生長期水分的需求及變色期水分對它的影響,從而為苗圃紅楓的栽培與管理提供理論依據和技術指導。
1 材料與方法
1.1 供試材料
本試驗選用株高、地徑相近的1年生美國紅楓改良品種扦插苗120株作為試驗苗,每盆1棵幼苗,盆的規格為高 250 mm、直徑330 mm并帶有底部托盤,每盆盛土量為7 kg。試驗用土為農田表土,自然曬干過2 mm篩,與蚯蚓糞以 36 ∶ 1 配比。試驗前肥水管理一致,試驗進行時不再施肥。試驗在沈陽農業大學后山進行,于2014年5月10日移入普通塑料大棚。進行適應性生長,8月初進行水分脅迫處理。
1.2 試驗設計
采用環刀法[7]測定所用土壤的土壤田間持水量(結果為26.17%),然后將120盆幼苗進行隨機分配,分成4組進行不同的水分梯度處理,每個處理3次重復。
處理A:重度干旱組,土壤水分含量為土壤田間持水量的35%,共30盆。
處理B:中度干旱組,土壤水分含量為土壤田間持水量的50%,共30盆。
處理C:輕度干旱組,土壤水分含量為土壤田間持水量的65%,共30盆。
處理D:對照組,土壤水分含量為土壤田間持水量的80%,共30盆。
土壤的干旱程度是通過連續的自然耗水以及人為的澆水方式得到的。試驗過程嚴格控制各處理土壤水分含量,當土壤水分達到設計標準時用透明塑料袋將盆套起來,以防止水分蒸發。采取稱質量法每隔 5 d,每日早晨07:00用感量 0.5 g 的電子稱稱質量測定1次土壤含水量,計算出需澆水量,低于處理供水下限時用大量筒準確均勻加入相應水量,補水到設定水分范圍,維持水分脅迫梯度。每個盆栽間保持一定距離,防止相互影響。用移動透明塑料棚防止自然降水淋入盆栽容器中。
1.3 測定指標及方法
植株生長狀況:緩苗期后,每15 d對植株的株高和地徑進行測量,進入快速生長期后,每7 d測1次,當3次測量數值不再變化時停止測量。苗高的測定是用鋼尺量取盆中土壤表面到最高葉片葉面的距離(精確到0.1 cm);地徑則是用游標卡尺選取植株距離土壤表面1 cm處進行測量(精確到 0.2 mm)。
當植株進入冬眠期停止生長時,將植株帶回實驗室進行鮮、干質量的測定。將取回的植株樣本清洗干凈,將水分徹底擦干,將根、莖分開,使用分析天平分別稱取鮮質量。把稱過鮮質量的植物材料裝入袋中,使用DHG-9240型干燥箱 105 ℃ 殺青20 min,然后在70 ℃狀態下烘4~6 h至恒質量,取出材料,在干燥器中冷卻到室溫,稱干質量。
葉片采集:秋季葉片變色期時,從有變色葉片開始,每隔5~7 d摘取葉片1次,直至葉片凋落,共摘5次。每次均在植株上、中、下3個部位各摘取1張健康的葉片,相同處理的樣本放在透氣性好的取樣袋中,帶回實驗室,進行色素含量的測定。
葉片色素含量測定:參照張憲政的方法[8],稍加改進。
花青素含量測定:參照何奕昆等的方法[9],稍加改進。
1.4 數據分析
應用Excel 2003和SPSS 19.0統計分析軟件進行數據處理與分析。
2 結果與分析
2.1 不同水分脅迫條件下美國紅楓苗木生長狀況
生物量是植物獲取能量的主要體現,水分脅迫將影響生物量在不同器官之間的分配[10]。從表1可以看出,土壤不同的水分含量對植物的生長產生了一定的影響。在4種水分處理的條件下,A、B、C、D組的株高與地徑的大小隨著土壤水分含量的增加而增大,其中D組的植株長勢最好,株高達到 63.43 cm,地徑為0.91 cm,平均葉片數為31張,且除了莖鮮質量、干質量以及根莖比3方面外,D組與A組的各項指標都有非常顯著的差異,總干質量比A組增加38.04%。B、C這2組只有在株高方面表現出了非常顯著的差異,地徑與根干質量方面表現出了顯著差異,而其他方面無顯著差異。在根莖比方面,C、D這2組雖然差異不顯著,但是可以看出,C組的平均根莖比要比D組稍大。由表2可以看出,土壤水分含量與各項生長指標都有相關關系,且為正相關。在株高、葉片數、根鮮質量、干質量以及總干質量方面表現與水分因子之間極顯著相關,表明在植株在這些方面受到土壤水分含量的影響大。
2.2 水分脅迫對美國紅楓幼苗葉片色素含量的影響
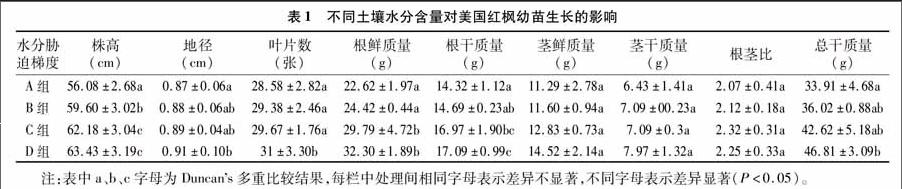
2.2.1 葉綠素含量的變化 (1)總葉綠素含量的變化。植物葉片呈色是相當復雜的,它與葉片細胞內色素的種類、含量以及在葉片中的分布有關[11]。而光合色素中的主要成分葉綠素參與光合作用光能的吸收、傳遞和轉換等過程。在本試驗中,紅楓幼苗變色期內,不同水分狀態下,苗木葉片葉綠素含量變化趨勢基本一致(圖1)。由圖1可以看出,在10月21日之前,葉綠素的含量變化整體呈下降趨勢,A組相對于其他組葉綠素下降趨勢較為平緩,在初期,也就是9月28日至10月6日之間,各組葉片中葉綠素含量迅速下降且D組下降速率最大,可能是由于10月1、2日連雨,氣溫驟降到0 ℃,導致葉片pH值下降,導致葉綠素脫鎂而不穩定[12]。到10月21日,A、B、C、D 4組葉片中葉綠素含量均達到最小值,為 0.024 8 mg/g。隨后,10月21—26日,除了土壤水分含量相對較少的A組外,其他組葉片中葉綠素含量又有了小幅度升高,而且B、C、D組的土壤水分比A組高。
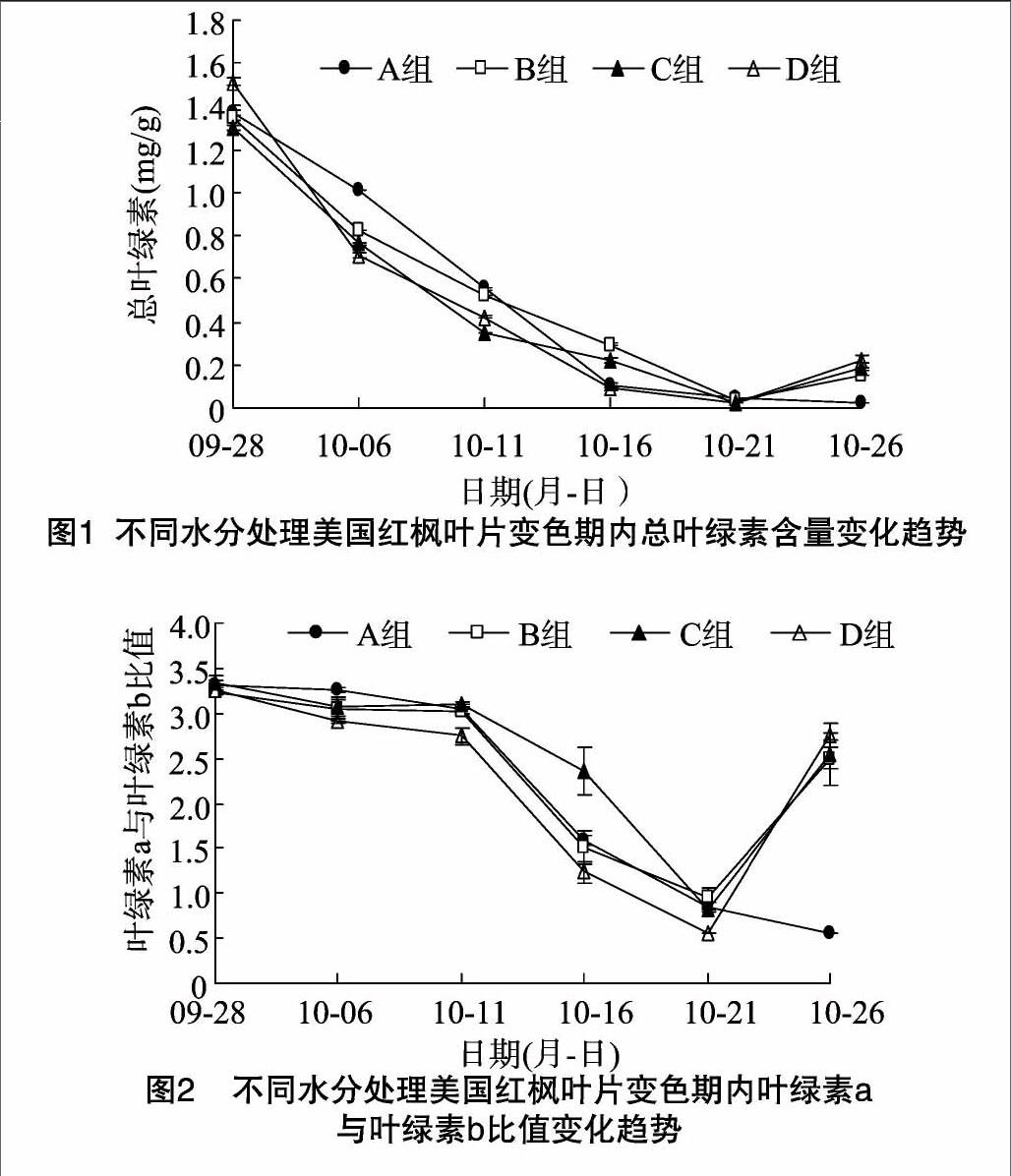
(2)葉綠素a與葉綠素b的比值變化。由圖2可以看出,B、C、D組葉片中葉綠素a與葉綠素b的比值變化趨勢相同,A組則不同于其他3組。9月28日至10月11日,4個組中葉綠素a與葉綠素b的比值緩慢減小,10月11—21日,B、C、D這3組中的比值減小速率加大,而此時A組的減小速率相對較緩慢。到在10月21日,B、C、D這3組葉片中葉綠素a與葉綠素b的比值達到最小,范圍在0.557~0.946。A組在10月21日后表現出來差異,呈現繼續下降趨勢。并且從數據可以看出,此時B、C、D組葉綠素a的含量明顯升高。
2.2.2 花青素相對含量的變化 花青素是一種水溶性色素,可以隨著細胞液的酸堿改變顏色,它在各色素中所占比重的大小是影響葉色變化的重要原因[13]。不同水分處理下葉片花青素含量的變化表現出一定的差異性。由圖3可以看出,在10月份以前至10月11日期間,4組表現出相同的變化趨勢,葉片中花青素含量都平穩上升。而在10月中旬,各組的花青素含量也有所上升,但是速度十分緩慢。在變色期的后期,也就是10月下旬,A、B、C、D組表現出了不同的變化趨勢,A、B這2組葉片中花青素含量繼續上升,最終B組的花青素含量高于A組。而C、D這2組葉片中花青素含量呈現降低的趨勢。最終葉片中花青素含量由高到低分別為B、A、C、D,且B組與D組的差值達到41.84 mg/g。
2.2.3 花青素含量與總葉綠素含量的比值變化 和葉綠素a和葉綠素b比值的情況相同,在花青素與葉綠素的比值中,A組同樣表現出了與其他3組不同的變化趨勢。由圖4可以看出,9月28日至10月21日期間,A、B、C、D組的花青素與葉綠素的比值都呈現上升的趨勢,開始上升的速率非常緩慢。直到10月16日時,上升速率明顯增大。在此期間,B、C、D組的上升速率表現出一致性,且在10月21日時,B、C、D組的比值都達到了最大。而A組在此時表現出了與其他組不同的情況,A組花青素與葉綠素比值上升的速率相對于其他3組較為緩慢。在此之后,A組與其他組的差異更加明顯,它呈現出繼續上升的趨勢,而B、C、D組的花青素與葉綠素的比值都表現出了下降趨勢。
3 結論與討論
水分條件是植物生長過程中的重要因素,植物受到水分脅迫時,在外部形態方面發生一系列的變化以適應水分脅迫。有研究表明[14],宏觀上表現為葉片面積縮小,植株矮化,葉厚度增加,葉生物量減少,微觀上體現為細胞增大,細胞增殖加快。本試驗中,土壤水分含量越多的處理,植株的長勢越好。喻曉麗等在對火炬樹幼苗的研究中也發現,水分脅迫顯著降低了火炬樹幼苗的株高生長和基徑生長[15]。但是輕度干旱組C組、中度干旱組B組以及重度干旱組A組的植株長勢并沒有非常明顯的差別,可能是因為水分脅迫的梯度差距不夠大的原因。輕度干旱組C組的根莖比略大于對照組D組,可能是由于適當的干旱脅迫會促進根的生長。有研究表明,隨著干旱脅迫的加劇,生物量更多的流向根中,來適應干旱的生存環境[16]。Zang等在研究水分脅迫對山毛櫸樹苗的影響時也發現,適當的干旱脅迫會促進細根的產生[17]。
彩葉植物葉色的表達是內在因素與外在因素共同作用的結果,而水分是植物生長發育的重要因素之一,土壤含水量的變化會影響植物葉片的內在結構和物質含量,從而影響葉片的色素合成及葉色表達[18]。本研究中,美國紅楓幼苗葉色變化期間內,除了返青時期,葉綠素不斷下降,而花青素含量逐漸增高,這與孫明霞等的研究結果[19]一致,葉片變色是由于葉片大量合成花青素的結果。同時Saure認為,在秋季隨著葉綠素的降解,花青素開始合成[20]。試驗中,總葉綠素含量在后期出現了增大的現象,這可能是因為此時的氣溫相對于月初有所升高,使葉片產生了返青現象[21]。而且此時B、C、D組的葉綠素含量由大到小分別為D(對照組)、C(輕度干旱組)、B(中度干旱組),可能由于土壤的濕度加高溫會使植物的高溫返青現象更加明顯,所以表現出后期葉綠素含量不同的變化趨勢,且土壤水分含量越多,返青現象越明顯。同時也有可能是因為長期的重度干旱使植物的調節能力降低了,而沒有出現返青現象。在葉綠素a與葉綠素b比值的變化中,土壤水分含量最多的D組,在返青現象前比值一直比其他組低,雖然中度干旱組B組與輕度干旱組C組表現差異不明顯,但是同樣可以認為這與許麗穎等研究的結果[22]一致,葉綠素a與葉綠素b的比值會隨著水分脅迫的加重而上升。而重度干旱組A組葉綠素a與葉綠素b的比值一直下降,可能是由于此組沒有出現返青現象造成的。在返青期間,從數據看出B、C、D組的葉綠素a含量明顯增加,這表明植物出現返青現象時,體內色素的變化主要表現為葉綠素a含量的增加。但是從葉綠素a和葉綠素b的數據中,并沒有看出賈利強等研究的結果,即水分脅迫會部分特異性地破壞葉綠素a,致使葉綠素a與葉綠素b比值降低[23]。
試驗中,隨著葉綠素含量的降低,植物體內花青素含量上升,與周肖紅等的研究結果[24]一致,黃櫨葉色變紅時,葉綠素下降,花青素含量上升。變色初期花青素含量上升速率較快,可能是由于10月初溫度的驟降引起的,有研究表明,低溫能誘導花青素苷的積累,高溫則會加速花青素苷的降解[25]。最終,土壤水分含量相對較多的輕度干旱組C組與對照組D組葉片中花青素含量較少,可能是由于出現了返青現象,導致葉片中葉綠素含量升高的緣故。而中度干旱組B組葉片中花青素含量最多,證明適度的干旱脅迫可以增加葉片中花青素的含量。王澤瑞認為,干燥的土壤環境使植物體內出現相對干旱,細胞內輕度缺水[26]。為了從環境中吸收水分,細胞需降低水勢,液泡中淀粉等不溶性大分子糖降解為可溶性的低分子糖,可導致細胞水勢降低,而可溶性糖含量的增加有利于花青素的形成。有研究表明,葉片的呈色是由花青素與葉綠素比值的大小決定的,比值越大,葉片顏色越紅[27]。本試驗中,在返青現象出現之前,雖然各組花青素與葉綠素比值的差別不大,但還是可以看出,對照組D組的比值要大于其他組。而且觀察結果也顯示D組葉片最早變紅,且顏色比其他組鮮艷,但是變化較為緩慢。
綜上所述,本試驗中,土壤水分含量為土壤田間持水量的80%時,最有利于美國紅楓幼苗生長。但是更多的水分含量是否會更有利于其幼苗的生長還有待進一步研究。并且適當的干旱脅迫會促進根的生長。土壤水分含量為土壤田間持水量的50%時,最有利于葉片中花青素的積累,雖然適當的干旱脅迫會促進花青素的合成,但是葉片呈色是多種色素綜合作用的結果。而土壤水分含量為土壤田間持水量的80%時,葉片最早變紅且鮮艷,但變化較為緩慢。因此,土壤水分含量在本研究的范圍內時,土壤田間持水量80%的水分含量最有利于美國紅楓幼苗的生長及變色。
參考文獻:
[1]羅永忠,成自勇. 水分脅迫對紫花苜蓿葉水勢、蒸騰速率和氣孔導度的影響[J]. 草地學報,2011,19(2):215-221.
[2]侯舒婷,張 倩,劉思岑,等. 黃金香柳對水分脅迫的生長與生理響應[J]. 西北植物學報,2014,34(12):2491-2499.
[3]楊躍文,季 蒙,包哈森高娃,等. 干旱脅迫對不同種源北沙柳幼苗生長特性的影響[J]. 林業科技,2013,38(6):6-9.
[4]Sherwin H W,Farrant J M. Protection mechanisms against excess light in resurrection lants Cruterastigna wilmsii and Xeroph taviscose[J]. Plant Growth Reg,1998,24:203-210.
[5]孔艷菊,孫明高,胡學儉,等. 干旱脅迫對黃櫨幼苗幾個生理指標的影響[J]. 中南林學院學報:自然科學版,2006,26(4):42-46.
[6]曹 晶,姜衛兵,翁忙玲,等. 夏秋季旱澇脅迫對紅葉石楠光合特性的影響[J]. 園藝學報,2007,34(1):163-172.
[7]袁娜娜. 室內環刀法測定土壤田間持水量[J]. 中國新技術新產品,2014(9):184-184.
[8]張憲政. 作物生理研究法[M]. 北京:農業出版社,1992:148-149.
[9]何奕昆,代慶陽,蘇學輝. 雁來紅葉色轉變與超微結構及色素含量的關系[J]. 四川師范學院學報:自然科學版,1995,16(3):195-198.
[10]胡繼超,姜 東,曹衛星,等. 短期干旱對水稻葉水勢、光合作用及干物質分配的影響[J]. 應用生態學報,2004,15(1):63-67.
[11]胡永紅,秦 俊,蔣昌華,等. 上海地區秋色葉成因的調查與分析[J]. 東北林業大學學報,2004,32(5):84-86.
[12]胡敬志,田 旗,魯心安. 楓香葉片色素含量變化及其與葉色變化的關系[J]. 西北農林科技大學學報:自然科學版,2007,35(10):219-223.
[13]張 平. 幾種常見彩葉植物的色素組成與葉色關系的研究[J]. 山東林業科技,2008,1(3):14-16.
[14]尉秋實,趙 明,李昌龍,等. 不同土壤水分脅迫下沙漠葳的生長及生物量的分配特征[J]. 生態學雜志,2006,25(1):7-12.
[15]喻曉麗,邸雪穎,宋麗萍. 水分脅迫對火炬樹幼苗生長和生理特性的影響[J]. 林業科學,2007,43(11):57-61.
[16]劉長利,王 文,崔俊茹,等. 干旱脅迫對甘草光合特性與生物量分配的影響[J]. 中國沙漠,2006,26(1):142-145.
[17]Zang U,Goisser M,Haeberle K H,et al. Effects of drought stress on photosynthesis,rhizosphere respiration,and fine-root characteristics of beech saplings:a rhizotron field study[J]. Journal of Plant Nutrition and soil Science,2014,177(2):168-177.
[18]李云飛,李彥慧,王中華,等. 土壤干旱脅迫對紫葉矮櫻葉片呈色的影響[J]. 生態學報,2009,29(7):3678-3684.
[19]孫明霞,王寶增,范 海,等. 葉片中的花色素苷及其對植物適應環境的意義[J]. 植物生理學通訊,2003,39(6):688-694.
[20]Saure M C. External control of anthocyanin formation in apple[J]. Scientia Horticulturae,1990,42(3):181-218.
[21]Graham T L. Flavonoid and flavonol glycoside metabolism in Arabidopsis[J]. Plant Physiology and Biochemistry,1998,36(1/2):135-144.
[22]許麗穎,赫玉蘋,王 剛,等. 水分脅迫對紫葉李葉片色素含量與PAL活性的影響[J]. 吉林農業大學學報,2007,29(2):168-172.
[23]賈利強,李吉躍,郎南軍,等. 水分脅迫對黃連木、清香木幼苗的影響[J]. 北京林業大學學報,2003,25(3):55-59.
[24]周肖紅,葛雨萱,王亮生,等. 黃櫨葉片變色期生理變化及植物生長調節劑對葉色的影響[J]. 林業科學,2009,45(7):59-62.
[25]胡 可,韓科廳,戴思蘭. 環境因子調控植物花青素苷合成及呈色的機理[J]. 植物學報,2010,45(3):307-317.
[26]王澤瑞. 幾種紅色系彩葉樹種光合特性及呈色機理的研究[D]. 合肥:安徽農業大學,2011:1-39.
[27]史寶勝,孟建朝,劉冬云,等. 秋季不同色澤五葉地錦葉片生理生化特性的研究[J]. 華北農學報,2009,24(增刊2):172-175.
