三葉木通的組織培養和多倍體誘導
2016-07-23 19:59:42石小兵楊航趙致
江蘇農業科學 2016年5期
石小兵++楊航++趙致
摘要:以三葉木通種子為外植體誘導產生叢生芽,再用不同濃度的秋水仙素溶液對叢生芽進行不同時間的誘導,并用染色體計數法檢測材料倍性。結果表明,叢生芽誘導的較好方式為消毒好的種子沿其背腹線縱切成半后接種于MS+1.0 mg/L 6-BA的培養基中誘導出無菌苗,然后將無菌苗接種于MS+2.0 mg/L 6-BA+0.3 mg/L NAA的繼代增殖培養基中培養。0.3%的秋水仙素處理24 h效果最好,多倍體誘導率達12%。
關鍵詞:三葉木通;組織培養;叢生芽;多倍體
中圖分類號: S567.904文獻標志碼: A文章編號:1002-1302(2016)05-0069-03
三葉木通[Akebia trifoliate (Thunb.) Koidz.]為木通科木通屬藤本植物,全株均可入藥,具有通經下乳、清熱利尿等功效,主要分布于甘肅、貴州、河北、河南、山東等地[1]。三葉木通果實具有發達的胎座組織,味甜可口,具有獨特的風味,含有許多人體必需營養成分[2-3],是一種具有開發潛力的保健水果,但存在果皮厚、種子多、可食率低的問題。多倍體植株具有生物產量增加、藥用活性成分增加、抗逆性增強、植株育性下降、少籽或無籽等特點,多倍體育種可成為三葉木通遺傳改良的途徑之一。植物離體誘導多倍體的方法已成功地在許多植物中取得成功,其中建立組培快繁體系是其必要環節。本試驗旨在以三葉木通幼嫩種子為外植體進行組織培養獲得叢生芽,再用秋水仙素對叢生芽進行處理,以獲得三葉木通多倍體材料,為無籽三葉木通的研究奠定基礎。
1材料與方法
1.1材料
三葉木通果實采自貴州省貴陽市花溪區棉花關,植株經貴州大學生命科學學院趙財副教授鑒定為木通科木通屬植物三葉木通。
1.2方法
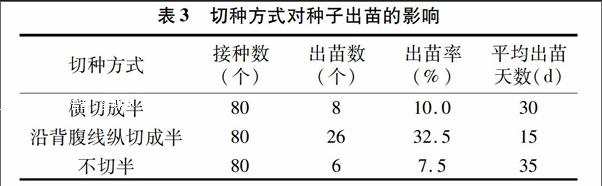
1.2.1外植體處理2014年8月中旬摘取三葉木通幼嫩果實,毛刷刷去表面塵土→洗潔精浸泡7 min→自來水沖洗20 min→超凈工作臺中取出種子→無菌水清洗種子數次→濾紙吸干表面水分→75%乙醇消毒15 s→0.1% HgCl2浸泡7 min→無菌水沖洗3次。將消毒好的種子橫切成半、沿背腹線縱切成半,以不切作對照處理,然后分別接種到MS培養基(添加蔗糖30 g/L、瓊脂7 g/L,pH值5.8~6.0,下同)中。每個處理接種20個培養瓶,每瓶4粒種子。(25±2) ℃黑暗培養40 d,統計出苗情況。
1.2.2無菌苗誘導培養采用合適的切種方式,將消毒好的種子接種在含6-BA不同濃度(0、0.5、1.0、1.5、2.0、2.5 mg/L)的MS培養基中進行無菌苗誘導,每個處理接種10個培養瓶,每瓶放5粒種子。(25±2) ℃黑暗培養30 d后,統計6-BA不同濃度處理下出苗率(出苗率=萌發外植體數/接種外植體數×100%)。
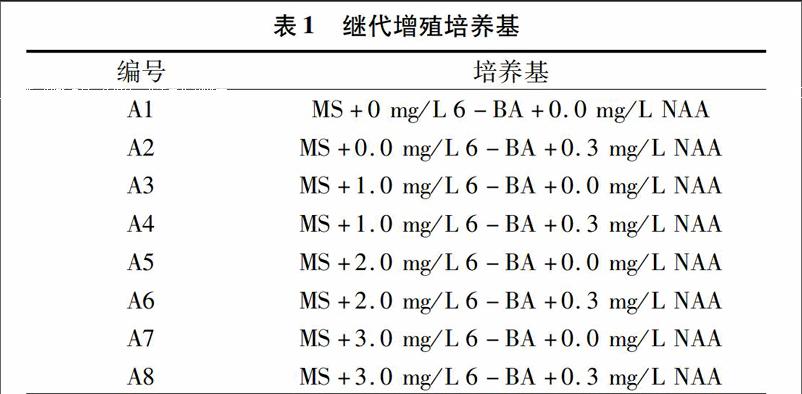
1.2.3繼代增殖培養以MS培養基為基本培養基,設置不同激素組合的處理,共8種培養基(見表1中A1~A8號培養基)。將無菌苗接種于這些培養基中,每種培養基接種10瓶,每瓶3個外植體。(25±2) ℃光照培養(光照時間 12 h/d,光照度1 000 lx)45 d統計增殖系數。
表1繼代增殖培養基
編號培養基A1MS+0 mg/L 6-BA+0.0 mg/L NAA A2MS+0.0 mg/L 6-BA+0.3 mg/L NAA A3MS+1.0 mg/L 6-BA+0.0 mg/L NAA A4MS+1.0 mg/L 6-BA+0.3 mg/L NAA A5MS+2.0 mg/L 6-BA+0.0 mg/L NAA A6MS+2.0 mg/L 6-BA+0.3 mg/L NAA A7MS+3.0 mg/L 6-BA+0.0 mg/L NAA A8MS+3.0 mg/L 6-BA+0.3 mg/L NAA
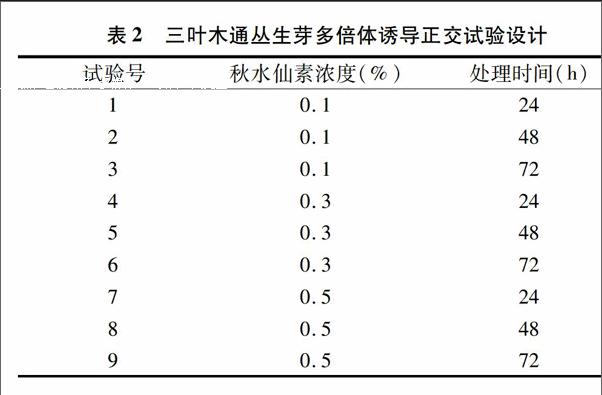
1.2.4多倍體誘導將生長良好的幼嫩叢生芽接入抽濾滅菌的秋水仙素溶液中,搖床振蕩培養。試驗采用2因素3水平L9(32)正交試驗試驗設計,試驗設計見表2。搖床轉速設定為50 r/min,培養溫度設定為24 ℃,光照24 h/d,光照強度為1 000 lx。處理完畢后轉接于MS+2.0 mg/L 6-BA+0.3 mg/L NAA的培養基中培養。培養條件同上。
1.2.5多倍體鑒定取出小叢生芽塊,放入卡諾氏固定液(無水乙醇 ∶冰乙酸體積比 3 ∶1)中,0 ℃固定1周后,將小叢生芽塊取出放到載破片上,用鑷子搗碎小叢生芽塊,用改良
`寶品紅染色液染色1~2 min,蓋上蓋破片后在光學顯微鏡下觀察,拍照。根據染色體數確定加倍處理材料的倍性。
2結果與分析
2.1切種方式對種子出苗的影響
從表3可知,沿背腹線縱切成半的種子出苗率最高(325%),橫切成半的次之,不切半的種子出苗率最低(75%);縱切成半的種子平均出苗天數最短(15 d),不切半的種子平均出苗天數最長(35 d)。表明縱切種子能加快種子出苗,其原因可能是縱切處理使種子的胚暴露于培養基中,胚無種皮限制而得以直接從培養基中吸收養分而快速生長;橫切可能破壞了胚的完整性或沒有使胚完全暴露于培養基中而對種子出苗沒有太大影響。
2.26-BA濃度對種子出苗的影響
消毒好的種子沿背腹線縱切成半,接種于培養基中,培養15 d左右種子便開始萌發,繼續培養15 d后無菌苗高度可達2 cm左右(圖1)。由表4可知,MS培養基添加6-BA后能提高種子出苗率。隨著6-BA濃度升高,種子出苗率呈先升高后下降的趨勢。當6-BA濃度為1.0 mg/L時出苗率最高,為60%。
2.3繼代增殖培養基篩選
由表5可知,當培養基中不添加6-BA時,NAA處理的無菌苗增殖系數均為1;當6-BA濃度一定時,添加 0.3 mg/L NAA與不添加NAA的相比,無菌苗增殖系數均有所提高;當NAA濃度一定,隨著6-BA濃度升高,無菌苗增殖系數先上升后下降,當6-BA濃度為2.0 mg/L時無菌苗增殖系數最高(5.70)。無菌苗轉接到增殖培養基中培養一段時間后其基部可分化出白色的不定芽(圖2);對不定芽繼代增殖后,能得到數目眾多的叢生芽,這些叢生芽具有分化成完整植株的能力。
2.4秋水仙素對叢生芽多倍體誘導的影響
秋水仙素濃度越低,叢生芽死亡率也越低(表6)。0.1%的秋水仙素處理下的叢生芽死亡率低,0.5%的秋水仙素處理過的叢生芽死亡率最高。0.1%的秋水仙素處理的叢生芽雖然有較低的死亡率,但并未得到加倍的叢生芽;0.5%的秋水仙素處理過的叢生芽得到了2塊加倍的叢生芽;0.3%的秋水仙素處理叢生芽后得到加倍的叢生芽塊數最多,其中以處理24 h的效果較好,誘導率為12%。同一濃度的秋水仙素在不同時間處理下多倍體誘導率并未呈現常見的先上升后下降趨勢,其原因可能是高濃度的秋水仙素對叢生芽的傷害大,使得存活下來的叢生芽數減少,并且生活力下降,從而使檢測到加倍的叢生芽塊數相應很少。
2.5多倍體鑒定
對三葉木通正常的根尖和叢生芽進行染色體計數,得出正常的三葉木通染色體數2n=2x=16,為二倍體(圖3),這與熊大勝等得出的結果[4]一致。成功加倍成多倍體的叢生芽染色體數為2n=4x=32,為四倍體(圖4)。這說明用秋水仙素誘導三葉木通獲得多倍體的方法是有效的。
3結論與討論
沈國林等以三葉木通葉片、莖段為外植體進行愈傷組織的誘導,但污染率高[5]。本試驗選用三葉木通幼嫩種子為外植體,建立三葉木通組織培養體系。選用幼嫩種子為外植體具有材料多(1個果實約有種子150~200粒)、污染率低(幼嫩種子在果實里處于一種相對無菌的環境)等優勢。
繼代增殖培養是組織培養的關鍵,篩選合適的增殖培養基才能達到離體快繁目的[6]。在無菌苗的繼代增殖過程中,6-BA和NAA組合是常用的激素組合[7-8]。在三葉木通無菌苗繼代增殖培養中發現,單獨使用NAA不能使無菌苗增殖,單獨使用6-BA可以使無菌苗增殖,附加NAA后增殖系數提高,表明6-BA在三葉木通繼代增殖培養中起主導作用。
叢生芽結構完整、材料幼嫩,可直接形成小植株,而且成苗率高,經化學誘變劑誘導處理后,易于獲得純的多倍體植株,所以本試驗采用了叢生芽作為誘導材料。秋水仙素是常用的化學誘變劑,通過抑制紡錘體微管的形成而導致染色體不能移向細胞兩極從而產生多倍體。秋水仙素對不同的植物和植物不同部位的誘導所需要的濃度和處理時間不同。三葉木通為木質藤本,對秋水仙素處理的濃度要求較高,在本試驗中體現在0.1%的秋水仙素處理叢生芽24、48、72 h均未檢測到多倍體細胞。雖然較高濃度秋水仙素處理過的叢生芽存活率小,但憑借其快速增殖的優勢,能獲得大量的加倍的叢生芽。
三葉木通多倍體的研究國內僅見熊大勝等用秋水仙素處理種子和幼苗的報道[9-10]。熊大勝等對加倍誘導的材料只進行了生理學和形態學鑒定。目前流式細胞儀能用于鑒定植株的倍性[11-12],使得常規誘導多倍體方法有了可靠的鑒定方法。但組培技術和化學誘導相結合的離體培養誘導法已成為一種最常用的染色體加倍方法。離體培養誘導法與常規的誘變育種方法相比,具有明顯的優越性。首先在組織培養條件下可以反復大批量地在培養瓶中處理植物愈傷組織、叢生芽,提高多倍體誘導的成功率;其次由于組織培養技術不受節氣等自然調節的影響,可以大大縮短誘導時間,并在短期內快速鑒定出大批量株系,繁殖大量試管苗。本試驗從種子無菌材料的獲得、無菌苗的誘導、繼代增殖培養及叢生芽多倍體誘導過程中建立了三葉木通無菌苗的組織培養體系,進行了三葉木通多倍體誘導和鑒定研究,可為三葉木通多倍體育種奠定基礎。
參考文獻:
[1]貴州植物志編輯委員會.貴州植物志(第二卷)[M]. 貴陽:貴州人民出版社,1985:141-142.
[2]劉倫沛,錢增秀. 三葉木通果中主要營養成分含量的測定[J]. 黔東南民族師范高等專科學校學報,2002,20(6):39-41.
[3]王曄,魯靜,林瑞超.三葉木通藤莖的化學成分研究[J]. 中草藥,2004,35(5):495-498.
[4]熊大勝,朱金桃,李興,等. 三葉木通染色體組型研究[J]. 中國野生植物資源,2000,19(6):18-20.
[5]沈國林,邵愛娟,黃璐琦,等. 三葉木通愈傷組織培養研究[J]. 中國中藥雜志,2007,32(10):899-901.
[6]李秋玲,李青,劉燕,等. 春石斛繼代培養主要影響因素[J]. 東北林業大學學報,2014,42(7):69-73.
[7]鄔秀宏,楊娟,李中林,等. 不同激素配比對‘渝茶1號繼代增殖生長的影響[J]. 西南農業學報,2013,26(4):1454-1458.
[8]周傳明,呂曼芳,陳奎,等. 山蒼子繼代培養中芽增殖效果研究[J]. 廣西科學,2012,19(4):374-376.
[9]熊大勝,雷紅梅,朱金桃,等. 三葉木通體細胞秋水仙素誘變苗期表型變異研究[J]. 經濟林研究,1994(1):58-62.
[10]熊大勝,曹庸,朱金桃,等. 三葉木通根尖秋水仙素誘變芽期根尖性狀及抗旱性變異研究[J]. 林業科技通訊,1994(7):20-21.
[11]方淑桂,陳文輝,曾小玲,等. 不同熟性大白菜小孢子植株倍性變異及倍性鑒定方法[J]. 福建農業學報,2009,24(4):304-307.
[12]王麗花,楊秀梅,吳學尉,等. 非洲菊大孢子再生植株倍性的快速鑒定方法[J]. 西北農業學報,2013,22(1):155-161.李婧,曾媛,龔勝,等. 大花三色堇FPNI-PCR反應體系的優化[J]. 江蘇農業科學,2016,44(5):72-75.
