湖南省柑橘潰瘍病菌gyrB和16S rRNA的擴增和序列分析
2016-07-23 20:07:43
江蘇農業科學 2016年5期
摘要:為了解湖南省不同地區柑橘潰瘍病菌遺傳多樣性,從湖南省黔陽、祁陽、衡山、道縣、茶陵、永興、沅江、零陵8個柑橘主產區采集樣本,分離病原菌,PCR擴增其gyrB基因和16S rRNA序列,陽性樣品送樣測序。利用生物信息學軟件,將測序結果與GenBank中柑橘潰瘍病菌相近的黃單胞桿菌構建系統發生樹,對比2個不同序列擴增的分辨率。結果表明,gyrB基因比16S rRNA序列分辨率高;利用gyrB基因除了能分析親緣較近的外源菌株,還能對同屬但是生長地區有異的菌株進行更深入的區分。
關鍵詞:柑橘潰瘍病;遺傳多樣性;gyrB基因;16S rRNA
中圖分類號: S436.661.1+2文獻標志碼: A文章編號:1002-1302(2016)05-0076-03
1912年首次在美國發現[1],中國相關最早的報道出現在1918年[2]。至今全球已有30多個國家和地區報道發現了該病害,占世界柑橘種植國家和地區的50%以上[3-4]。柑橘潰瘍病對我國柑橘產業有較大影響,不僅降低了全國柑橘產量和品質,還制約了我國柑橘產業的發展[5];其中發病嚴重的地區包括湖南、福建、廣東、廣西等。柑橘潰瘍病是一種細菌性病害,病原菌為地毯草黃單胞柑橘致病變種(Xanthomonas axonopodis pv. citri),病菌主要通過自然孔口和傷口感染健康植株。該病菌在葉、枝梢及果實的病斑中越冬,翌年春條件適宜時從病部溢出,借風雨、昆蟲傳播,經寄主的氣孔、皮孔和傷口侵入,使葉片、嫩梢和果實發病[6]。葉片受害,開始在葉背產生針頭大小的黃色、油漬狀小點,后逐漸擴大。同時,在葉片正反兩面逐漸隆起成圓形病斑,病部表皮破裂,病斑木栓化如海綿狀,表面粗糙,灰褐色。其后中央凹陷并有細微輪紋,周圍有黃色暈圈,在暈圈的內側常有褐色釉光邊緣,后期病斑中心開裂并且凹陷,呈火山口狀。柑橘潰瘍病自然條件下傳播距離較近,主要途徑是風雨、蟲害和農作時造成的傷口以及植株間摩擦等[7]。
rRNA是細菌系統分類學研究中最常用的“分子鐘”,其種類少,含量大,可占到細菌RNA總量的80%[8]。由于其大小適中且保守性很好,所以廣泛應用于物種間的遺傳分化距離和親緣關系的研究中。16S rRNA是一種常用核糖體RNA,其高度的保守性[9]使之成為分子生物學研究的一個熱點。
促旋酶(gyrase)B亞單位基因簡稱為gyrB,是一種細菌共有的蛋白質編碼基因[10]。隨著分子生物學的發展,gyrB基因序列擴增技術在細菌親緣關系及系統發育研究中的作用越來越明顯[11],可以用其作為分子標記來研究細菌的系統發育水平,還可區分近源種和菌株。已知的gyrB序列,包括其相關產物的gyrB數據被存放于由日本建立的ICB[12]數據庫中,用于細菌分類鑒定方面的研究和細菌親緣種的鑒定。Dauga等研究表明,比較腸桿菌科(Enterobacteriaceae)不同屬的系統進化關系時,gyrB基因比16S rRNA更加適用于種屬內或種屬間的精確遺傳距離分析和發育關系比較[13] 。
中國柑橘潰瘍病研究表明,部分地區的菌種中仍然存在更細致的分化[14],本研究采集湖南省主要柑橘產區的柑橘潰瘍病樣本,分離致病菌進行純化培養及致病性檢測,提取病原菌DNA選取多種引物進行PCR擴增,最終選出自行設計的gyrB基因片段的特異性引物[15]和16S rRNA引物[16]來分析湖南柑橘潰瘍病的遺傳多樣性。
1材料與方法
1.1材料
供試材料取自于湖南省黔陽、祁陽、衡山、道縣、茶陵、永興、沅江、零陵等地的柑橘主要種植區,大部分為葉片,少部分為發病果實。采用的致病菌標準菌由湖南農業大學國家柑橘改良中心長沙分中心提供。
1.2方法
1.2.1柑橘潰瘍病菌分離、鑒定按稀釋分離法對采集的柑橘潰瘍病樣本進行病原菌分離[17],得到的病原物經JYF5/JYR5引物進行PCR擴增、驗證。JYF5/JYR5引物序列為:JYF5:5′-TTCGGCGTCAACAACCTG-3′,JYR5:5′-AACTCCAGCACATACGGGTC-3′,目標片段長度在410左右,基本都擴增到了目標條帶,從 8個地區的樣品中經過分離和鑒定共獲得了22個病原菌的菌株。
1.2.2gyrB引物gyrB序列引物是通過Primer Premier 5.0軟件[18]設計的引物GGS1F/GGS2R,其序列為GGS1F:5′-CCCTGCTGCTGACCTTCTTCT-3′和GGS2R:5′-GGTTGACCGTGG TTTCCCATA-3′。反應參數:預變性95 ℃ 4 min;變性95 ℃ 30 s,退火62 ℃ 30 s,延伸72 ℃ 30 s,35個循環;延伸72 ℃ 7 min,4 ℃保存。
1.2.316S rRNA序列16S rRNA序列的引物為27F:5′-AGAGTTTGATCMTGGCTCAG-3′和1492R:′-TACGGHTACCTT ACGACTT-3′。 反應參數:預變性95 ℃ 4 min;變性 95 ℃ 30 s,退火58 ℃ 30 s,延伸72 ℃ 30 s,40個循環;延伸72 ℃ 7 min,4 ℃保存。
1.2.4序列分析和系統發育樹的構建gyrB及16S rRNA序列引物PCR擴增產物送至上海生工測序,測序結果用軟件MEGA 4.0的非加權平均數聚類法(Unweighted Pair Group Method with Arithmatic mean,UPGMA)構建系統發育樹。
2結果與分析
2.1gyrB引物的PCR擴增結果
通過GGS1F/GGS2R引物對22個已經鑒定的柑橘潰瘍病病原菌進行PCR擴增,目標片段的大小約為760 bp,擴增產物進行凝膠電泳,結果見圖1。
2.216S rRNA序列PCR擴增結果
16S rRNA序列引物PCR擴增22個經過鑒定的菌株,目標片段的大小約為1.5 ku,PCR擴增產物進行凝膠電泳,結果見圖2。
2.3構建gyrB和16SrRNA序列的系統發育樹
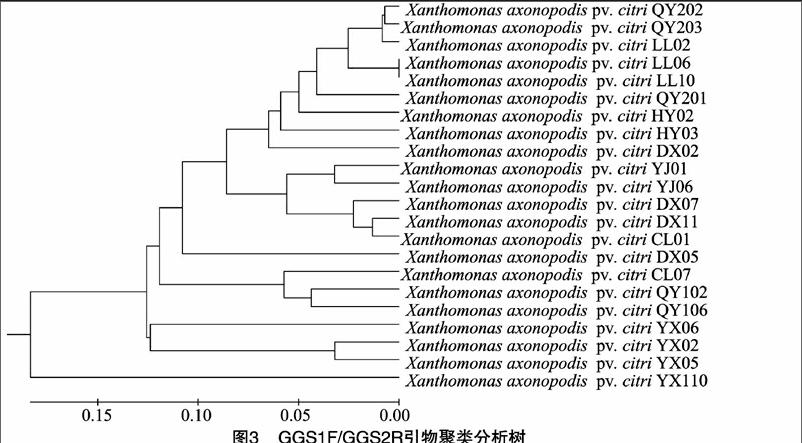
2.3.1gyrB序列系統發育樹gyrB序列引物擴增產物序列建樹結果(圖3)表明,不同地區的病原菌樣品間的遺傳距離比較明顯。在遺傳距離大于0.18時22個供試菌株為同一簇,當遺傳距離為0.055時,所有的菌株可以分成7個不同的簇,其分布情況基本與菌株采樣地點在湖南省的地理位置相關。
2.3.216S rRNA序列系統發育樹16S rRNA 引物擴增產物序列建樹結果(圖4)表明,所采用的菌株樣品之間的遺傳距離非常小。遺傳距離為0.0046左右時,所有的菌株一共可以分為3個不同的簇,其分布規律基本與地域的差距有很大關系。
3討論
通過16S rRNA及gyrB序列引物PCR擴增結果建樹可以明顯看到湖南省中部、東部及北部部分樣品,西部和部分北部樣品,南部、部分西部樣品分成了3個遺傳距離相對大一點的3部分。其分布與病原菌來源地的位置有著較大聯系,而且寄主植物差異也是造成遺傳差距的主要因素之一,且gyrB基因序列分辨率要明顯高于16S rRNA序列。16S rRNA序列分析僅能在屬的水平區分這些菌株,gyrB序列分析還可將區分程度擴展到屬內小種。
16S rRNA序列在目前使用比較廣泛,但其不足之處也逐漸顯露。16S rRNA基因序列相比gyrB基因序列所攜帶的信息量多,長度適中,但其分辨率較低。gyrB引物擴增序列在菌株分類程度上有了進一步的提高,在湖南省柑橘潰瘍病的小種分析中gyrB基因序列的分辨率較前者高,效果比較好;但是由于gyrB基因的序列較短,鑒定細菌的準確度依然不夠。目前,gyrB基因序列在分子生物學中應用不多,還需要進一步開發利用。
參考文獻:
[1]Dopson R N. The eradication of citrus canker[J]. Plant Dis Rept,1964,48:30-31.
[2]Lee H A. Further data on the susceptibility of rutaceous plants to citrus canker[J]. Agr Res,1918,15:661-665.
[3]Gottwald T R,Graham J H,Schubert S T. Citrus canker:the pathogen and its impact [R/OL].[2015-12-20]. http://www.plantmanagementnetwork.org/pub/php/review/citruscanker/.
[4]Schubert T S,Rizvi X,Sun T R. Meeting the challenge of eradicating citrus in Florida-Again[J]. Plant Disease,2001,85:340-356.
[5]楊楓. 多胺在提高柑橘潰瘍病抗性中的作用及其機理研究[D]. 武漢:華中農業大學,2014.
[6]陳小帆,莫瑾,左靜,等. 利用雙重PCR技術檢測柑橘潰瘍病菌[J]. 華南農業大學學報,2010,31(3):32-35.
[7]Gottwald T R,Timmer L W. The efficacy of windbreaks in reducing the spread of citrus canker caused by Xanthomonas campestris pv. citri[J]. Tropical Agriculture,1995,72(3):194-201.
[8]Hou W G,Wang S,Dong H L,et al. A comprehensive census of microbial diversity in hot springs of Tengchong,Yunnan Province,China using 16S rRNA gene pyrosequencing[J]. PLoS One,2013:e81.
[9]Hiroshi M,Maruyama F,Kato H,et al. Design and experimental application of a novel non-degenerate universal primer set that amplifies prokaryotic 16S rRNA genes with a low possibility to amplify eukaryotic rRNA genes[J]. DNA Research,2014,21:217-227.
[10]Kirby B M,Everest G J,Meyers P R. Phylogenetic analysis of the genus Kribbella based on the gyrB gene:proposal of a gyrB-sequence threshold for species delineation in the genus Kribbella[J]. Antonie Van Leeuwenhoek,2010,97(2):131-142.
[11]Carrasco G,Valdezate S,Garrido N,et al. Identification,typing,and phylogenetic relationships of the main clinical nocardia species in Spain according to their gyrB and rpoB genes[J]. Journal of Clinical Microbiology,2013,51(11):3602-3608.
[12]Teng L J,Hsueh P R,Huang Y H,et al. Identification of bacteroides thetaiotaomicron on the basis of an unexpected specific amplicon of universal 16S ribosomal DNA PCR[J]. Journal of Clinical Microbiology,2004,42(4):1727-1730.
[13]Dauga C. Evolution of the gyrB gene and the molecular phylogeny of Enterobacteriaceae:a model molecule for molecular systematic studies[J]. International Journal of Systematic and Evolutionary Microbiology,2002,52(2):531-547.
[14]彭耀武. 中國柑橘潰瘍病病原菌多態性研究[D]. 重慶:西南大學,2014.
[15]安然,易圖永,肖啟明,等. gyrB基因在細菌分類和檢測中的應用[J]. 江西農業學報,2010,22(4):18-20,24.
[16]李霞,高謙.16S rRNA基因序列分析在臨床微生物學中的應用[J]. 微生物與感染,2006,1(3):184-186.
[17]董玉蘭. 湖南省柑橘潰瘍病菌系分化及拮抗菌篩選[D]. 長沙:湖南農業大學,2012.
[18]張新宇,高燕寧. PCR引物設計及軟件使用技巧[J]. 生物信息學,2004,2(4):15-18,46.曾云英,萬強,馬存琛. 抑菌劑及接種條件在矮牽牛開放式快速繁殖中的效果[J]. 江蘇農業科學,2016,44(5):79-80.
