生物炭對鎘脅迫下水稻生長及光合產量的影響
2016-07-23 18:07:12張學艷曹瑩孟軍
江蘇農業科學 2016年5期
張學艷++曹瑩++孟軍
摘要:選用遼寧地區水稻主推品種沈農9816,采用盆栽試驗,研究了生物炭對鎘脅迫下水稻農藝性狀以及光合產量的影響。結果表明,水稻農藝性狀以及產量指標均呈現Cd1(1 mg/kg)>Cd2(2 mg/kg)>Cd0(0 mg/kg)>Cd4(4 mg/kg)的趨勢,添加生物炭后,水稻的生物量、光合參數、產量等指標均呈現C21(4 t/hm2 )>C0(0 t/hm2 )>C210(40 t/hm2 )的趨勢。另外,在各鎘處理下,添加生物炭C21、C210與C0相比,C21處理各指標增幅均大于對照,且在Cd2濃度下增幅最大。說明生物炭可減輕鎘脅迫、促進水稻生長以及提高光合作用,并且最佳用量為C21處理,在Cd2濃度下效果最好。
關鍵詞:生物炭;鎘脅迫;光合;產量;水稻;生長
中圖分類號: S511.01文獻標志碼: A文章編號:1002-1302(2016)05-0097-05
隨著現代工農業的不斷發展,重金屬尤其是土壤重金屬污染日趨嚴重,據統計,我國已有超過28萬hm2的農田遭受重金屬污染[1]。水稻作為全球尤其是亞洲最主要的糧食作物,為65%的中國人口提供主食[2]。鎘作為“五毒之首”[3],是稻田中最主要的重金屬污染元素[4],它能夠通過植物根系吸收,在植物體內產生富集,并通過食物鏈直接威脅食品生產安全以及人類健康[5-6]。生物炭具有較大的比表面積,較強的強吸附力、抗微生物和化學分解能力,可以在土壤中存留更長時間,為植物提供養分[7-8]。此外,生物炭可以降低重金屬的生物有效性,對重金屬起到很好的固定效果,減少農作物對重金屬的吸收,降低毒害作用[9],成為研究土壤重金屬污染改良的熱點。近年來,生物炭被認為是解決全球性環境、氣候、糧食問題的有效途徑,引起了國際國內的普遍重視[10-11]。在我國,以生物炭為核心的秸稈炭化還田不僅可以有效利用生物質資源,增加土壤輸入,而且對于保障糧食有效供給尤其具有重要意義[12]。生物炭取之于農,用之于農,在治理重金屬污染、改良土壤方面展現出巨大的應用價值。目前,利用生物炭來緩解鎘對水稻脅迫影響的研究報道尚不多見,因此,本試驗以水稻為研究對象,探討不同用量生物炭對重金屬鎘脅迫下水稻生長及光合產量的影響,旨在探明減輕鎘脅迫、促進水稻生長以及提高光合作用的最佳生物炭用量。
1材料與方法
1.1試驗材料
選用遼寧地區主推水稻品種沈農9816,生物炭由玉米秸稈350 ℃低溫厭氧燒制而成,其中鎘含量為0.073 mg/kg,pH值為9.23,全氮8.73 mg/kg,全磷8.6 mg/kg,全鉀 48.7 mg/kg,硫2.97 mg/kg。土壤采用農田土,棕壤,有機質含量1.18%,堿解氮含量107.33 mg/kg,速效磷含量 34.52 mg/kg,速效鉀含量98.41 mg/kg,全鎘0.15 mg/kg,有效鎘0.09 mg/kg,pH值5.9,陽離子交換量17.21 cmol/kg。
1.2試驗設計
采用盆栽試驗,在沈陽農業大學北山試驗基地進行。試驗設有2個處理因素:鎘和生物炭。鎘試驗濃度0、1、2、4 mg/kg,記為Cd0、Cd1、Cd2、Cd4,生物炭添加量:0、4、40 t/hm2,折合每盆炭施用量為0、21、210 g,記為C0、C21、C210。盆栽用土風干過篩,裝入聚乙烯桶(高30 cm,直徑28 cm),每盆裝風干土12.75 kg,于秧苗移栽前1個月將Cd以CdCl2·2.5H2O的形態與土混合,保持盆內水層,使土壤與Cd進行平衡。每盆栽3穴,每穴2苗,各處理12盆,一共144盆,肥料施入量(按土壤質量計算)分別為:尿素0.428 g/kg、CaHPO4·2H2O 0.523 g/kg、K2SO4 0.247 g/kg,于分蘗期每盆追施尿素0.118 g/kg,抽穗期每盆追施尿素0063 g/kg,其余正常管理。取樣時期:分蘗期、灌漿期和蠟熟期;成熟期考種測產。在水稻整個生育期,保持水肥條件一致,及時防治病蟲草害,保證水稻正常生長發育。
1.3測定項目與方法
1.3.1生物量測定在各取樣時期,采用常規方法測量水稻的株高、分蘗數;采用加拿大產WinRHIZO根系分析系統測量總根長、總根體積、總根表面積;取各器官鮮樣在105 ℃下殺青30 min,75 ℃烘干至恒質量后用百分之一的電子天平測各部分的干質量。
1.3.2葉綠素取劍葉去掉葉脈,剪碎,稱0.04 g,加入10 mL 95%乙醇避光浸泡48 h,測前搖勻,用分光光度計測定,按以下公式計算葉綠素含量:Ca=13.95D665 nm-688D649 nm;Cb=24.96D649 nm-7.32D665 nm;Wa=(Ca·V·k)/m;Wb=(Cb·V·k)/m;Wc=Wa+Wb。式中:Ca、Cb分別為葉綠素a和葉綠素b的濃度;Wa、Wb 、Wc分別為葉綠素a、葉綠素b以及總葉綠素含量;V為提取液體積;k為稀釋倍數;m為樣品鮮質量。
1.3.3葉面積采用Epson Expression 11000XL葉面積掃描系統測量。
1.3.4光合指標采用CIRAS-3光合儀(美國PP Systems 公司)在晴朗的上午,09:00—11:00,測定凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)、胞間CO2濃度(Ci)等指標。選擇功能葉為長勢一致的水稻劍葉進行測定,每次測量均為葉片中部(正面)。
1.3.5產量指標成熟期每處理取3盆水稻考種,測定每盆穗數、穗粒數、結實率、千粒質量、每盆產量等指標,采用電子分析天平(感量0.001 g)稱質量。
1.3.6數據處理采用Excel 2010、SPSS 17.0對數據進行統計分析。
2結果與分析
2.1生物炭對鎘脅迫下水稻生長的影響
由表1可知,生物炭添加下,隨鎘濃度的增大,水稻株高、分蘗數、地上干質量、地下干質量、總根長、總根體積、總根表面積整體上均呈現先增加后降低的趨勢,且均在Cd1濃度達到最大值,呈現Cd1>Cd2>Cd0>Cd4的趨勢,與空白組趨勢一致。
相同鎘濃度條件下,隨著生物炭用量的增加,除了總根長、總根體積、總根表面積在分蘗期呈現C210>C21>C0的趨勢,其余各農藝性狀在整個生育時期均呈現先增加后降低的趨勢,C21處理效果最好(表1)。
在不同鎘濃度處理下,添加生物炭即C21、C210與C0相比,C21處理各農藝性狀均有所增加,C210有所降低,說明C21處理對水稻生長的促進作用更明顯。在分蘗期,C21與C210相比,2種生物炭添加量均在Cd4處理增幅最大,且C21各指標增幅更大,C210除在分蘗數、地上干質量比對照有所降低外,其余指標均增加,說明生物炭在水稻生長前期對高濃度鎘處理效果好,且C21處理比C210處理的促進作用更明顯。添加 C21處理后,株高、地上干質量、地下干質量均在 Cd1濃度達到最大增幅,根系形態指標在Cd2濃度達到最大增幅,分蘗數在Cd4濃度達到最大增幅。蠟熟期,C21處理均比對照有所增加,株高、分蘗數、總根長、總根體積均在Cd1濃度達到最大增幅,地上干質量、總根表面積均在Cd4濃度達到最大增幅,地下干質量在Cd2濃度達到最大增幅,說明在水稻的生長后期,添加生物炭對各農藝性狀來看,增幅不如生長前期明顯(表1)。2.2生物炭對鎘脅迫下水稻葉綠素及光合能力的影響
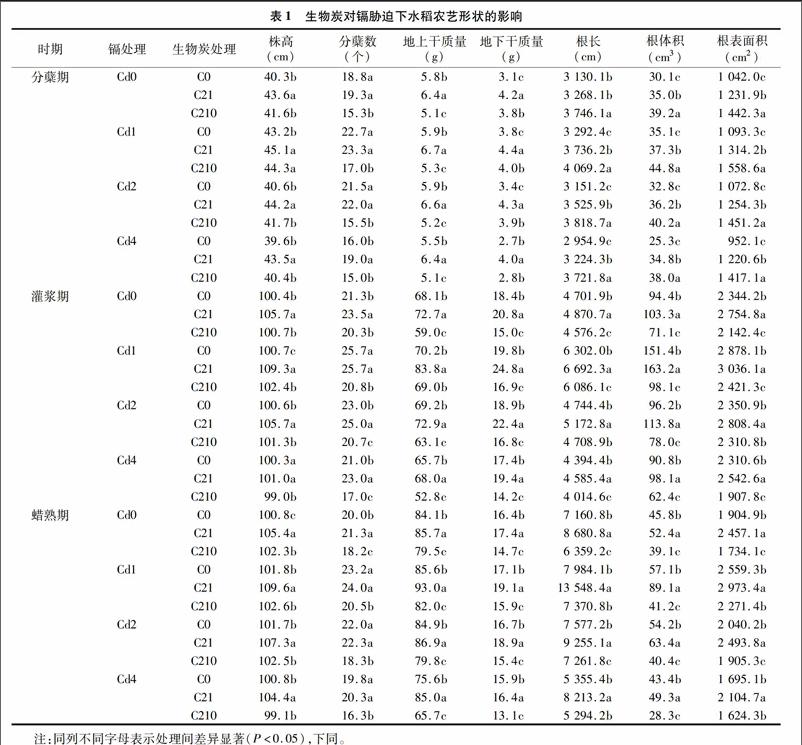
2.2.1對葉綠素含量的影響圖1顯示,同一炭量添加下,葉綠素總含量隨著鎘濃度的增加先增加后降低,即Cd1>Cd2>Cd0>Cd4。同一鎘濃度添加條件下,葉綠素總含量隨著生物炭量的增加呈現C21>C0>C210的趨勢,且達到差異顯著水平(P<0.05)。在Cd1濃度下,C21在3個生育時期分別比對照高2.17%、1.60%、3.04%;在Cd2濃度下,C21在3個生育時期分別比對照高2.60%、6.89%、4.59%;在Cd4濃度下,C21在3個生育時期分別比對照高2.20%、5.15%、3.41%,且C210均比對照低,說明適量的生物炭(C21處理)對水稻體內葉綠素的形成具有促進作用,而過量的生物炭(C210處理)會產生抑制作用,且C21處理在Cd2濃度下效果最好。在水稻的整個生育時期,葉綠素總含量呈現逐漸降低的趨勢,即分蘗期>灌漿期>蠟熟期,說明隨著水稻的逐漸衰老,葉片失綠變黃,體內葉綠素總含量也隨之降低。
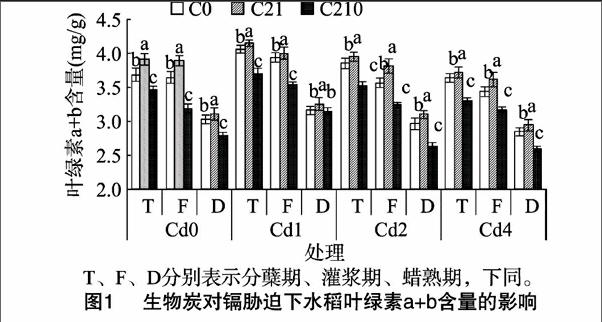
2.2.2對葉面積的影響圖2顯示,在無生物炭添加條件下,隨著鎘濃度的增加,水稻分蘗期葉面積變化趨勢不顯著(P>0.05),相同炭量施加條件下,隨著鎘濃度的增加,葉面積呈現升高后降低的趨勢,即Cd1>Cd2>Cd0>Cd4。在相同鎘濃度處理下,葉面積隨著生物炭量的增加呈現先增加后降低的趨勢,C21處理的葉面積達到最大值,C210處理達到最小值,且達到差異顯著水平(P<0.05),這與水稻的農藝性狀趨勢吻合。在整個生育時期,各處理組均呈現灌漿期>蠟熟期>分蘗期的趨勢,這與生長前期葉片數量少且小、生長后期葉片衰老死亡有關。
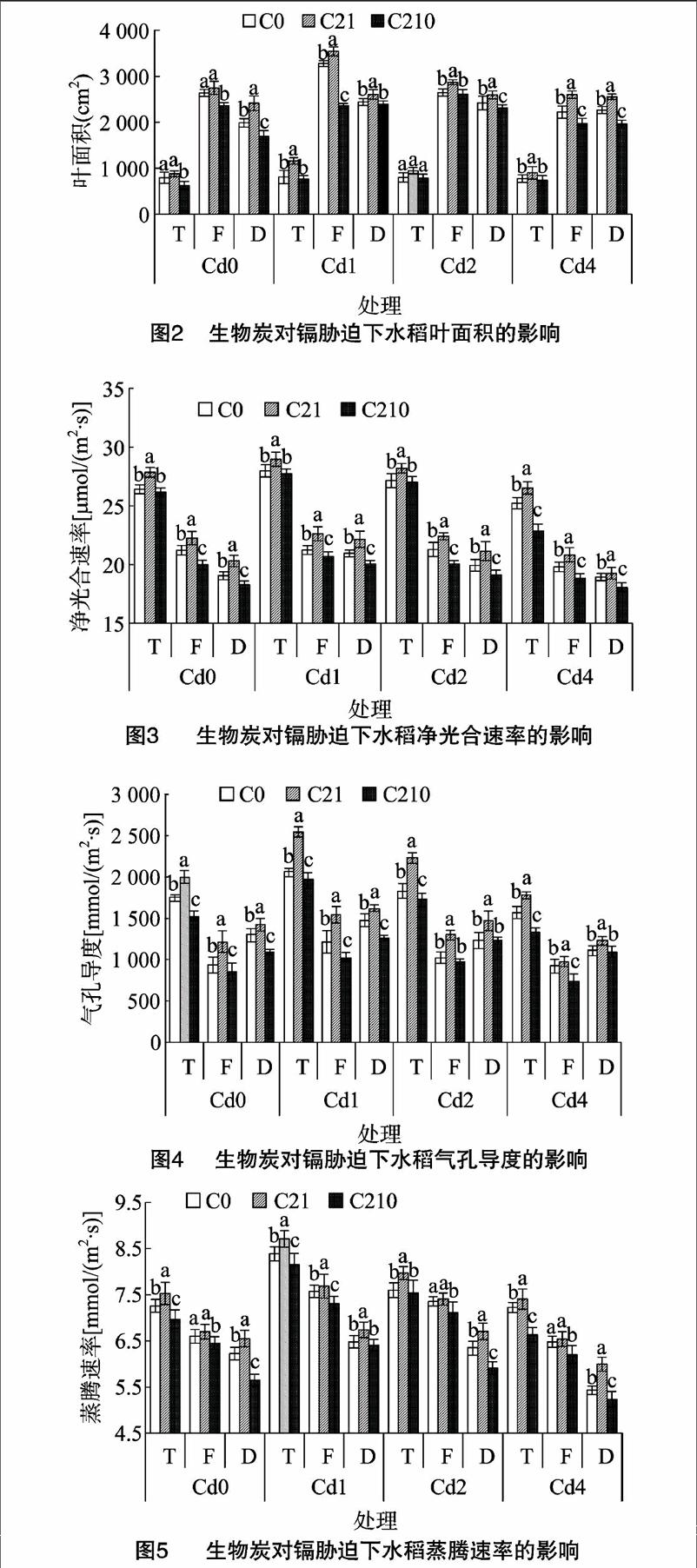
2.2.3對光合能力的影響圖3至圖6顯示,同一生物炭量添加下,水稻凈光合速率、氣孔導度、蒸騰速率、胞間CO2濃度隨著鎘濃度的增加呈現先升高后降低的趨勢。同一鎘濃度條件下,水稻凈光合速率、氣孔導度、蒸騰速率、胞間CO2濃度隨著生物炭量的增加呈現先增加后降低的趨勢,說明不管是否有鎘脅迫作用,添加生物炭都可以促進水稻凈光合速率、氣孔導度、蒸騰速率、胞間CO2濃度的增加,其中C21處理的效果最好,C210處理光合能力最弱,各處理達到差異顯著水平(P<0.05)。在水稻的整個生長期,水稻的凈光合速率、蒸騰速率均在分蘗期達到最大值,其次為灌漿期,最后為蠟熟期,這是因為隨著水稻生長期的不斷推移,葉片開始衰老,導致光合能力也隨之減弱;氣孔導度在分蘗期達到最大值,灌漿期達到最小值,蠟熟期又有所回升;胞間CO2濃度在蠟熟期達到最大值。
2.3生物炭對鎘脅迫下水稻產量的影響
降低,Cd1濃度達到最大值,Cd4濃度達到最小值。在相同鎘濃度處理下,隨著炭量的增加,每盆穗數、穗粒數、結實率、每盆產量均隨呈現先增加后降低的趨勢,其中C21處理表現最好,其次是C0處理,即C21>C0>C210,且各處理達到差異顯著水平(P<0.05)。添加生物炭C21、C210與C0對比發現,在各產量指標中,C21處理的增幅均大于C210,在C21處理下,每盆穗數、結實率、產量等指標均在Cd2濃度下增幅最
大,穗粒數、千粒質量在Cd4濃度下增幅最大。在各處理中,C21-Cd1處理每盆產量達到最大值,C210-Cd4處理達到最小值。在各鎘處理組,C21處理產量分別比對照高7.21%、806%、7.48%,C210處理產量分別比對照低10.42%、928%、10.41%,由此說明,添加適量的生物炭可以促進水稻產量的增加,過量的生物炭反而會導致水稻減產,并且在鎘脅迫下,C21增產效果在Cd2濃度最好,這與水稻農藝性狀在生育后期Cd2濃度增幅效果最好相吻合。
2.4水稻葉綠素、葉面積、光合特性與產量之間的相關性
通過相關性分析(表3),葉綠素總含量與葉面積、蒸騰速率呈顯著正相關(P<0.05),與凈光合速率、氣孔導度、胞間CO2濃度、產量呈極顯著正相關(P<0.01)。葉面積與凈光合速率、氣孔導度、蒸騰速率呈顯著正相關(P<0.05),與胞間CO2濃度在分蘗期、灌漿期呈顯著正相關(P<0.05),與產量呈極顯著正相關(P<0.01)。凈光合速率與氣孔導度、胞間CO2濃度、產量呈極顯著正相關(P<0.01),與蒸騰速率呈顯著正相關(P<0.05)。氣孔導度與蒸騰速率、胞間CO2濃度、產量呈極顯著正相關(P<0.01)。蒸騰速率與胞間CO2濃度在分蘗期、蠟熟期呈顯著正相關(P<0.05),與產量呈極顯著正相關(P<0.01)。胞間CO2濃度與產量呈極顯著正相關(P<0.01)。
3討論與結論
株高、根系形態以及生物量等性狀指標常被用作評價作物重金屬耐性的重要指標[13]。本研究表明,同一炭量添加下,隨著鎘濃度的增加,水稻的株高、分蘗數、生物量以及總根長、總根體積、總根表面積會隨之先升高后降低,這與“少促多抑”的研究結果相同[14]。同一鎘濃度條件下,在水稻的整個生育時期,適量的生物炭(C21處理)可以促進水稻的生長發育,過量的生物炭(C210處理)在水稻生長后期有抑制作用,張晗芝等對玉米苗期的研究也表明,過量的生物炭(48 t/hm2) 對玉米的株高、生物量有顯著抑制作用[15]。根系作為植物主要的吸收器官,其生長和分布直接影響植物對重金屬的吸收能力和吸收效率,根系的變化也直接反映了重金屬對植株的毒害程度。本試驗表明,添加生物炭后,在分蘗期明顯促進了水稻根系的生長,說明在水稻生長前期,生物炭有利于促進根系生長,隨著時間的推移,生物炭的促進作用不明顯,過量的生物炭甚至會抑制根系的生長。這可能與生物炭的理化性質有關,生物炭具有較高的含碳量,礦物養分含量低,施入土壤后會降低土壤養分[16],如果施入量過多,會影響水稻根系對土壤養分的吸收,進而影響地上部分的生長。
葉綠素是光合作用的物質基礎,其變化必然直接或間接地影響光合作用[17-18]。本研究表明,隨著鎘濃度的增加,葉綠素含量降低,使得光合速率也隨之降低,而光合速率的降低是葉片衰老的表現,說明鎘脅迫會導致水稻葉片衰老,降低光合產量。同一鎘濃度條件下,隨著生物炭添加量的增加,水稻葉綠素總含量、葉面積、光合參數以及產量指標均隨之先增加后降低,表明適量的生物炭(C21處理)對其有良好的促進作用。另外,分蘗期、灌漿期追施了一定量的尿素,也保證了水稻生長所需的養分,兩方面的共同作用使得葉面積增加,單位面積葉綠素的含量得到提升,進而為進行更高效的光合作用提供了物質基礎,同時,較大的葉面積可以獲取更多的光能,提高水稻的光合作用,積累更多的光合產物,增加產量。
Pn、Tr、Ci、Gs 作為重要的光合參數,是衡量植物逆境脅迫下光合強度的重要指標[19]。本試驗研究表明,同一炭量添加下,隨著鎘濃度的增加,水稻凈光合速率、氣孔導度、蒸騰速率、胞間CO2濃度均隨之先升高后降低,Cd1濃度達到最大值,說明低濃度鎘脅迫下,葉綠素含量越高,越有利于提升水稻對鎘的耐性,增強光合能力,提高產量。隨著鎘濃度的增加,鎘耐性降低,產量隨之降低,這與Daud等[20]、錢雷曉等[21]的研究結果相同。施加生物炭(C21)后,不管有無鎘脅迫,均可以提高凈光合速率、氣孔導度、蒸騰速率與胞間CO2濃度,提高水稻光合能力。按照Farquhar等[22]的研究,Ci與Gs 協同下降時,Pn的下降是由氣孔限制,而本試驗表明,胞間CO2濃度隨著凈光合速率的降低而增大,說明是非氣孔限制。
對產量指標的研究表明,隨著鎘濃度的增加,水稻每盆產量先增加后降低,Cd1濃度產量最高,Cd4濃度產量最低,這與鎘脅迫下水稻分蘗有關,低濃度鎘可以促進水稻分蘗,增加有效穗數,提高產量;隨著鎘濃度的增加,水稻分蘗減少,有效穗數降低,產量下降。張秀芝等的研究也表明,隨著重金屬Cd、As、Cr濃度的增加,籽粒產量明顯下降[22]。本試驗也表明,添加生物炭后,C21處理組的每盆產量大于無炭添加C0處理,也大于過量生物炭C210處理,說明生物炭能緩解鎘脅迫、增加水稻產量,最佳用量為C21處理,即每盆施加21 g生物炭,折合大田用量為4 t/hm2,且在Cd2濃度下效果最好。
參考文獻:
[1]雷武生,楊寶林,戴金平.硫肥對鎘脅迫下不同基因型水稻抗氧化系統和光合特性的影響[J]. 河北農業大學學報,2014,37(2):12-17.
[2]鄭悅,鄭桂萍,趙洋,等. 生物炭對粳稻墾鑒稻5號穗部性狀及產量的影響[J]. 江蘇農業科學,2015,43(6):59-62.
[3]陳志良,莫大倫,仇榮亮.鎘污染對生物有機體的危害及防治對策[J]. 環境保護科學,2001,27(4):37-39.
[4]黃冬芬,奚嶺林,王志琴,等. 結實期灌溉方式對水稻品質和不同器官鎘濃度與分配的影響[J]. 作物學報,2008,34(3):456-464.
[5]Weng B S,Xie X Y,Weiss D J,et al. Kandelia obovata (S.,L.) Yong tolerance mechanisms to cadmium:subcellular distribution,chemical forms and thiol pools[J]. Marine Pollution Bulletin,2012,64(11):2453-2460.
[6]Su,Y,Liu,et al. Effect of iron deficiency on subcellular distribution and chemical forms Cd in peanut roots in relation to its translocation[J]. Environmental and Experimental Botany,2014,97:40-48.
[7]楊曉慶,侯仔堯,常夢婷,等. 生物炭對鎘污染土壤的修復研究[J]. 江蘇農業科學,2015,43(6):335-337.
[8]楊放,李心清,王兵,等. 生物炭在農業增產和污染治理中的應用[J]. 地球與環境,2012,40(1):100-107.
[9]徐楠楠,林大松,徐應明,等. 生物炭在土壤改良和重金屬污染治理中的應用[J]. 農業環境與發展,2013,30(4):29-34.
[10]Kleiner K. The bright prospect of biochar[J]. Nature Reports-Climate Change,2009,3(6):72-74.
[11]Woolf D,Amonette J E,Street-Perrott F,et al. Sustainable biochar to mitigate global climate change[J]. Nature Communications,2010,1(56):1-9.
[12]孟軍,張偉明,王紹斌,等. 農林廢棄物炭化還田技術的發展與前景[J]. 沈陽農業大學學報,2011,42(4):387-392.
[13]慈敦偉,姜東,戴廷波,等. 鎘毒害對小麥幼苗光合及葉綠素熒光特性的影響[J]. 麥類作物學報,2005,25(5):88-91.
[14]Wei S,Li Y,Zhan J,et al. Tolerant mechanisms of Rorippa globose (Turcz.) Thell. hyperaccumulating Cd explored from root morphology[J]. Bioresource Technology,2012,118:455-459.
[15]張晗芝,黃云,劉鋼,等. 生物炭對玉米苗期生長、養分吸收及土壤化學性狀的影響[J]. 生態環境學報,2010,19(11):2713-2717.
[16]陳心想,何緒生,耿增超,等. 生物炭對不同土壤化學性質、小麥和糜子產量的影響[J]. 生態學報,2013,33(20):6534-6542.
[17]姜永雷,唐探,陳嘉裔,等. 鎘脅迫對水蕨幼苗葉綠素熒光參數和生理指標的影響[J]. 江蘇農業科學,2015,43(9):357-360.
[18]趙素貞,洪華龍,嚴重玲. 鈣對鎘脅迫下秋茄葉片光合作用及超微結構的影響[J]. 廈門大學學報:自然科學版,2014,53(6):875-882.
[19]魏花朵,李悅,陳忠林,等. 氮肥對鎘脅迫下結縷草光合和葉綠素熒光特性的影響[J]. 中國土壤與肥料,2015(4):88-92.
[20]Daud M K,Sun Y Q,Dawood M,et al. Cadmium-induced functional and ultrastructural alterations in Roots of two transgenic cotton cultivars[J]. Journal of Hazardous Materials,2009,161(1):463-473.
[21]錢雷曉,胡承孝,趙小虎,等. 鎘脅迫對不同基因型小白菜氮代謝和光合作用的影響[J]. 華中農業大學學報,2015,34(3):69-75.
[22]Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology,1982,33:317-345.
[23]張秀芝,李強,彭暢,等. 不同添加量重金屬對水稻產量及籽粒重金屬富集的影響[J]. 吉林農業科學,2015,40(4):13-16.閆佳,張均,馬超,等. 氮素水平對冬小麥地上部分鋅積累和轉運的影響[J]. 江蘇農業科學,2016,44(5):102-106.
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
軍事文摘(2021年22期)2021-11-26 00:43:51
小讀者(2021年2期)2021-03-29 05:03:48
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
