棉花陸海雜交親本及子代光合葉綠素熒光參數(shù)分析
2016-07-23 18:29:00王志軍謝宗銘董永梅
江蘇農(nóng)業(yè)科學 2016年5期
王志軍++謝宗銘++董永梅
摘要:以海島棉(新海22號)、陸地棉(自育品系589)和以其為親本配置的正反交雜種F1、F2為試驗材料,大田自然環(huán)境條件下,測定花鈴期倒四葉的葉綠素含量、光合參數(shù)、葉綠素熒光參數(shù),結(jié)果發(fā)現(xiàn),海島棉的葉綠素a、葉綠素b、總?cè)~綠素含量、Fm、ΦPSⅡ、Fv/Fm顯著高于陸地棉(P<0.05);胞間CO2濃度(Ci)、Fo、Y(NO)、Y(NPQ)顯著低于陸地棉(P<0.05);葉綠素a/b、蒸騰速率(Tr)、氣孔導度(Gs)、光合速率(Pn)、水分利用效率(WUE)、ETR、qP、NPQ差異不顯著;陸海雜交F1在Fm、NPQ、Y(NPQ)、Fv/Fm等熒光參數(shù)表現(xiàn)出中親優(yōu)勢,葉綠素a含量、葉綠素b含量、總?cè)~綠素含量、蒸騰速率(Tr)、氣孔導度(Ci)、凈光合速率(Pn)、電子傳遞速率(ETR)表現(xiàn)出超親優(yōu)勢,其余參數(shù)均沒有表現(xiàn)出雜種優(yōu)勢;陸海雜交F2的大部分光合葉綠素熒光參數(shù)表現(xiàn)出不同程度的優(yōu)勢退化,變化較為復(fù)雜,這可能與陸海雜交F2代性狀瘋狂分離有關(guān)。通過從光合生理方面挖掘陸海雜交后代潛在優(yōu)勢,以期探明陸海雜交后代光合及葉綠素熒光參數(shù)的雜種優(yōu)勢表現(xiàn)。
關(guān)鍵詞:海島棉;陸地棉;光合作用;葉綠素熒光參數(shù)
中圖分類號: S562.03文獻標志碼: A文章編號:1002-1302(2016)05-0134-04
水稻、玉米、油菜等農(nóng)作物的雜種優(yōu)勢利用,已取得了巨大的經(jīng)濟效益和社會效益,棉花雜種優(yōu)勢是提高棉花產(chǎn)量、改善纖維品質(zhì)、增強抗逆性的一種有效途徑。陸地棉產(chǎn)量高、適應(yīng)性廣,但纖維品質(zhì)性狀不足,海島棉纖維品質(zhì)優(yōu)良,但生育期較長、產(chǎn)量低[1],棉花海陸雜種表現(xiàn)出巨大的雜種優(yōu)勢:產(chǎn)量高、鈴多而大、品質(zhì)好。張小全等研究認為海陸種間雜交在株高、果枝數(shù)、果節(jié)數(shù)、單株結(jié)鈴數(shù)顯著高于陸陸雜交[2]。菲力申等研究認為陸海雜種F1代較親本在產(chǎn)量和品質(zhì)性狀均具有超親優(yōu)勢[3-4]。石玉珍等研究認為海陸雜交種纖維長度、伸長率、整齊度和強度等性狀均具有正向優(yōu)勢[5]。李哲等研究認為,海陸雜交F1代纖維長度、比強度、馬克隆值3大品質(zhì)性狀均可達到或超過海島棉親本[6]。對于海島棉及陸海雜交棉的光合及葉綠素熒光參數(shù)的研究也有較多報道。鄧仲萀等研究認為,陸海雜種的光合速率介于雙親之間,不表現(xiàn)雜種優(yōu)勢[7]。姜磊等研究認為,棕色棉與白色棉雜交F1的凈光合速率也沒有表現(xiàn)出明顯的雜種優(yōu)勢[8]。伍維模等研究認為,吐絮期陸地棉對高光強光能的利用能力強于海島棉,而對低光強光能的利用能力弱于海島棉[9]。關(guān)于棉花海陸雜種優(yōu)勢的研究多集中在農(nóng)藝性狀的優(yōu)勢表現(xiàn),而對于光合特性和葉綠素熒光參數(shù)雜種優(yōu)勢表現(xiàn)的研究仍相對較少,本研究將對陸海雜交種光合葉綠素熒光參數(shù)的雜種優(yōu)勢表現(xiàn)進行深入研究。
本研究將以海島棉、陸地棉為親本,測定了親本與正反交雜種F1、F2代,花鈴期倒四葉的葉綠素含量及光合-葉綠素熒光參數(shù),探討陸海雜交后代光合性能及葉綠素熒光特性的雜種優(yōu)勢表現(xiàn),以期從光合生理方面挖掘海陸雜交后代的潛在優(yōu)勢,進一步為棉花利用雜種優(yōu)勢提供理論依據(jù)。
1材料與方法
1.1植物材料
試驗材料為海島棉(新海22號)、陸地棉(自育品系589)和以其為親本配置的正反交后代F1代和F2代,于2013年5—11月在新疆農(nóng)墾科學院生物技術(shù)研究所試驗地(45°19′ N,86°03′ E,10 ℃以上活動積溫為3 570~3 729 ℃,7月份平均氣溫23.9~25.5 ℃)進行,采用隨機區(qū)組設(shè)計,行長5 m,膜下滴灌栽培,膜寬2.05 m,1膜3管(滴灌帶)6行,株距10 cm,行距20 cm,每材料播種2行,3次重復(fù),田間管理同大田,灌水方式為有壓滴灌。
1.2測定方法和參數(shù)
1.2.1光合色素測定當棉花發(fā)育至花鈴期,于09:30取倒四葉,每個材料取3張葉,葉綠素的提取與測定采用高俊鳳的方法[10] 。
1.2.2光合參數(shù)隨機選取各材料長勢一致的植株掛牌標記,晴天11:00—13:00,使用德國WALZ公司的GFS-3000光合儀,利用自然光強,大氣CO2濃度約為(370±10) μmol/mol,葉室溫度25 ℃,快速測定標記植株倒四葉的凈光合速率[Pn,μmol/(m2·s)]、氣孔導度[Gs,mmol/(m2·s)]、胞間CO2濃度(Ci,μmol/mol)等光合參數(shù)、每個材料測10株,分別取平均值。
1.2.3葉綠素熒光參數(shù)選擇晴天09:30—13:00 利用德國WALZ公司的MINI-PAM熒光儀,測量上述標記葉片的葉綠素熒光參數(shù),先暗適應(yīng)30 min,測定暗適應(yīng)下的初始熒光(Fo)、最大熒光(Fm)得到Fv/Fm,此時打開光化光,可測定對應(yīng)光強下葉片的實際光量子產(chǎn)量ΦPSⅡ及其他所有的熒光參數(shù),每個材料測10張葉,取平均值。
1.2.4雜種優(yōu)勢計算
中親優(yōu)勢率=F1-MPMP×100%;
超親優(yōu)勢率=F1-HPHP×100% 。
式中:F1為F1代雜交種觀察值;MP為雙親平均值;HP為高值親本觀察值。
2結(jié)果與分析
2.1光合色素
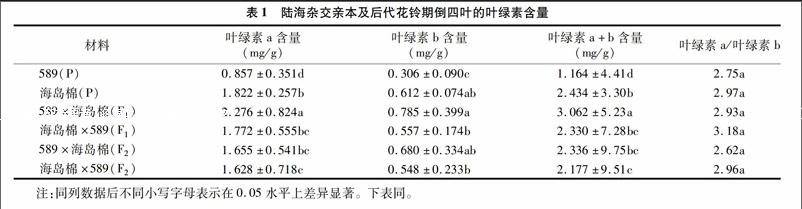
由表1可知,589為陸地棉親本,海島棉新海22號為海島棉親本,589×海島棉(F1)為正交F1,海島棉×589(F1)為反交F1,589×海島棉(F2)為正交F2,海島棉×589(F2)為反交F2。海島棉的葉綠素a、葉綠素b、總?cè)~綠素含量均顯著高于陸地棉品系589(P<0.05),589分別是新海22號的47%、50%、48%。正交F1葉綠素a、葉綠素b、總?cè)~綠素含量的中親優(yōu)勢率為70%、71%、70%,超親優(yōu)勢率分別為25%、28%、26%。反交F1中親優(yōu)勢率分別為32%、21%、30%,超親優(yōu)勢率分別為-3%、-9%、-4%,正交F2的葉綠素a、葉綠素b、總?cè)~綠素含量分別比F1降低37.52、15.44、31.08百分點,顯著高于589,但顯著低于海島棉(P<0.05),反交F2的葉綠素a、葉綠素b、總?cè)~綠素含量與F1代差異不顯著,分別降低885、1.64、7.03百分點,但高于父本589,低于海島棉,差異均顯著(P<0.05)。說明海島棉較陸地棉葉綠素含量高,海島棉做父本時,雜種F1代在葉綠素含量方面有較強的雜種優(yōu)勢,而且正反交之間差異顯著,F(xiàn)2代時,雜種優(yōu)勢退化顯著,正反交F2之間差異不顯著。親本及雜種后代的葉綠素a/葉綠素b值差異均不顯著(P<0.05)。
2.2光合參數(shù)
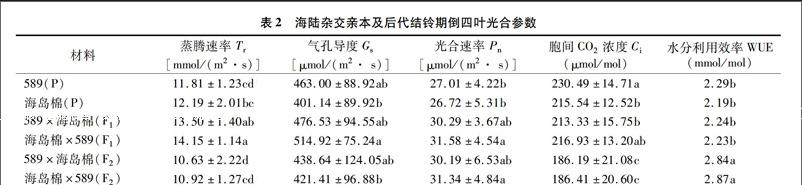
由表2可知,陸地棉589和海島棉新海22的蒸騰速率(Tr)、凈光合速率(Pn)、氣孔導度(Gs)、水分利用效率(WUE)等參數(shù)無顯著差異;胞間CO2濃度(Gs)海島棉是陸地棉的93.51%,差異顯著(P<0.05)。正反交F1之間光合參數(shù)及WUE均無顯著差異(P<0.05)。正交F1的Tr顯著高于雙親,中親優(yōu)勢率為12.50%,超親優(yōu)勢率為10.74%,Gs差異均不顯著,中親優(yōu)勢率10.30%,超親優(yōu)勢率為2.9%,Pn中親優(yōu)勢率為12.75,超親優(yōu)勢率為12.21%,Gs較海島棉升高但差異不顯著,較陸地棉親本顯著降低,是陸地棉親本(589)的73.44%,其他參數(shù)與雙親均沒有顯著差異(P<005)。反交F1的Tr顯著高于雙親,中親優(yōu)勢率為17.92%,超親優(yōu)勢率為16.08%,Gs的中親優(yōu)勢率為 19.18%,超親優(yōu)勢率為11.21%,Pn中親優(yōu)勢率為17.55%,超親優(yōu)勢率為16.92%,Ci,WUE與雙親差異均不顯著(P<0.05)。正交F2的Tr比 F1降低26.99百分點、比兩親本降低且差異顯著,Gs比F1下降8.64百分點,比陸地棉親本降低,比海島棉親本升高,差異均不顯著,Ci比F1下降14.58百分點,比兩親本均降低且差異顯著,Pn和F1差異不顯著,比親本升高,差異不顯著,WUE比F1增加26.78百分點,比兩親本顯著升高。反交F2的Tr比F1降低29.58百分點,比兩親本降低,Gs比F1下降22.19百分點,比陸地棉親本降低,但比海島棉親本升高,差異均不顯著、Ci比F1下降16.37百分點,比兩親本均降低且差異顯著,Pn和F1差異不顯著,比親本升高,差異顯著,WUE比F1增加28.70%,比兩親本顯著升高(P<0.05)。
2.3葉綠素熒光參數(shù)
Fo為最小熒光,當光系統(tǒng)Ⅱ的所有反應(yīng)中心均處于開放態(tài)時得到Fo。表3親本之間差異顯著,海島棉是589的8106%,正反交F1之間差異不顯著,正交F1中親優(yōu)勢率為5.69%,負超親優(yōu)勢率為18.03%,反交F1中親優(yōu)勢率為-6.79%,負超親優(yōu)勢率為4.1%,正交F2比F1降低8.19百分點,差異不顯著,較陸地棉親本降低,差異不顯著,較海島棉親本顯著升高,反交F2比F1顯著升高,增幅達23.23百分點,較兩親本均升高。
Fm為暗適應(yīng)樣品的最大熒光,當光系統(tǒng)Ⅱ的所有反應(yīng)中心均處于關(guān)閉狀態(tài)時測得Fm,親本之間差異顯著,589是海島棉的84.90%,正反交F1之間沒有顯著差異。正交F1中親優(yōu)勢率為8.61%,超親優(yōu)勢率為0.41%,反交F1中親優(yōu)勢率為1.18%,負超親優(yōu)勢率為6.46%,正交F2較F1下降達1415百分點,較陸地棉親本升高,比海島棉親本降低,差異均不顯著,反交F2較F1顯著增高,增幅13.38百分點,較兩親本均增高。
ΦPSⅡ是光系統(tǒng)Ⅱ的實際量子產(chǎn)量,親本之間差異顯著,589是海島棉的88.36%,正反交F1之間沒有顯著差異。正交F1中親優(yōu)勢率為-0.59%,超親優(yōu)勢率為-6.82%,反交F1中親優(yōu)勢率為2.33%,超親優(yōu)勢率為-3.77%,正反交F2之間也沒有顯著差異,正交F2較F1下降3.31百分點,差異不顯著,較親本589升高,較海島棉下降,反交F2較F1降低105百分點,差異也不顯著,較589升高,較海島棉下降。
ETR及相對電子傳遞速率,在光化光強為 1 116 μmol/(m2·s) 下測定,親本之間沒有顯著差異,589是海島棉的87.05%,正反交F1之間也沒有顯著差異,正交F1中親優(yōu)勢率為21.66%,超親優(yōu)勢率為15.52%,反交F1中親優(yōu)勢率為24.21%,超親優(yōu)勢率為16.17%,正反交F2之間差異顯著,正交F2是反交的184.48%,正交F2較F1顯著升高16.30百分點,較雙親顯著升高,反交F2較F1下降62.03百分點,較雙親下降,但差異不顯著(P<0.05)。
qP是光化學淬滅系數(shù),由光合作用引起的熒光淬滅,反映了光合活性的高低。由表3可以看出,親本之間qP差異不顯著,589是海島棉的98.65%,正反交F1之間差異也不顯著,正交F1中親優(yōu)勢率為-1.09%,超親優(yōu)勢率為-1.77%,反交F1中親優(yōu)勢率為0.05%,負超親優(yōu)勢率為0.63%,正反交F2之間差異顯著,正交F2較F1降低2.27百分點,較雙親較低,反交F2較F1上升1.46百分點,較雙親增高。
NPQ為非光化學熒光淬滅,反映了植物耗散過剩光能為熱的能力,是衡量植物光保護能力的重要指標。由表3可知,親本之間NPQ差異不顯著(P<0.05),海島棉NPQ是589的3571%,正反交F1代差異也不顯著,正交F1中親優(yōu)勢率為49.47%,超親優(yōu)勢率為1.43%,反交F1中親優(yōu)勢率為1759%,負超親優(yōu)勢率為20%,正反交F2之間差異不顯著,正交F2較F1下降12.68百分點,較親本589降低,較海島棉親本升高,反交F2較F1增高39.13百分點,較雙親升高。
Y(NO)為非調(diào)節(jié)性能量耗散,是光損傷的重要指標,親本之間差異顯著,海島棉是589的71.9%,正反交F1之間差異不顯著,正交F1中親優(yōu)勢率為-0.64%,負超親優(yōu)勢率為14.6%,反交F1中親優(yōu)勢率為-7.8%,負超親優(yōu)勢率為208%,正交F2較F1升高11.11百分點,較親本589降低,較海島棉升高,反交F2和F1無顯著差異,較親本589降低,較海島棉升高。
3結(jié)論與討論
海島棉的品質(zhì)優(yōu)于陸地棉,纖維長、細度高、強度大,是重要的紡織品原料[7]。陸地棉皮棉產(chǎn)量高,但品質(zhì)較差,陸地棉與海島棉種間的雜種一代,可綜合雙親的優(yōu)良性狀,利用其雜種優(yōu)勢,在保持高產(chǎn)的同時,可提高纖維品質(zhì),被認為是一條可行的途徑[12]。
葉綠素含量與光合作用密切相關(guān),是衡量光合作用強弱的重要因素[13-14]。植物葉綠素含量及葉綠素a/葉綠素b值變化是植物本身的遺傳特性[15],同時在一定程度上受環(huán)境條件的影響[16-18]。本研究結(jié)果表明,花鈴期海島棉(新海22號)的葉綠素a、葉綠素b、總?cè)~綠素含量均高于陸地棉(589),這與張亞黎等研究發(fā)現(xiàn)海島棉新海22光合色素含量高于陸地棉新陸早13的結(jié)果[15]一致。陸海雜交F1表現(xiàn)出超親優(yōu)勢,優(yōu)勢率為25%~28%,海陸雜交F1葉綠素a、葉綠素b、總?cè)~綠素含量高于陸地棉但低于海島棉,正反交之間有顯著差異(P<0.05)正反交F2優(yōu)勢退化明顯,略高于陸地棉親本,但顯著低于海島棉,說明以海島棉做父本,陸海雜交F1代葉綠素含量具有明顯優(yōu)勢。
已有研究表明,海島棉葉片的實際光合能力低于陸地棉,但兩者具有相似的光合潛力[19]。張亞黎等研究發(fā)現(xiàn),早晨和下午外界光強較低時,海島棉新海22號和陸地棉新陸早13號的Pn無明顯差異外,其余時間段新海22號葉片的Pn值顯著低于新陸早13號。本研究結(jié)果表明,陸地棉589Pn、Gs、Ci、WUE略高于海島棉新海22號,但差異不顯著(P<0.05),Tr低于新海22號。Gs反映外界環(huán)境CO2通過氣孔進入到葉肉細胞葉綠體羧化部位的限制程度,是決定Pn高低的一個重要因素[20],新海22號Gs下降,導致Pn的降低,而Tr卻高于陸地棉,可能與海島棉葉片柵欄組織較薄以及葉片形態(tài)有關(guān)。正反交F1在Tr、Gs、Pn、WUE等光合參數(shù)表現(xiàn)出雜種優(yōu)勢,和Chen等研究認為雜種棉通過維持相對較高的光合能力來提高皮棉產(chǎn)量的研究結(jié)果[21]一致,正反交F2表現(xiàn)出明顯的優(yōu)勢退化現(xiàn)象,但WUE顯著增加,可能與陸海雜交后代葉型改變有關(guān)。
葉綠素熒光是光合作用的有效探針,通過對各種熒光參數(shù)的分析,可以得到有關(guān)光能利用途徑的信息[22]。Fo和Fm分別是暗適下的最小熒光和最大熒光,它們是光系統(tǒng)Ⅱ(PSⅡ)反應(yīng)中心處于完全開放和關(guān)閉時的熒光產(chǎn)量,F(xiàn)o和Fm在陸海雜交F1代并未表現(xiàn)出明顯優(yōu)勢,海島棉做父本,雜交F2代的Fo和Fm值高于F1和親本,ΦPSⅡ=Yield=(Fm′-Fs)/Fm′[23]任一光照狀態(tài)下PSⅡ的實際量子產(chǎn)量,在相同環(huán)境條件下,海島棉的實際量子產(chǎn)量顯著高于陸地棉,正反交F1的實際量子產(chǎn)量介于兩者之間,沒有表現(xiàn)出雜種優(yōu)勢,正反交F2的實際量子產(chǎn)量也介于海島棉和陸地棉之間。ETR代表了光電子傳遞的能力[24],雜種F1表現(xiàn)出了一些優(yōu)勢,但不明顯,陸海雜交F2顯著高于F1和親本,反交F2電子傳遞速率顯著降低,因此可以選用陸地棉做母本,比較容易選育高光效品種。qP=(Fm′-Fs)/Fv′=1-(Fs-Fo′)/(Fm′-Fo′)[25](基于“沼澤模型”)即由光合作用引起的熒光淬滅,反映了光合活性的高低,植物吸收的光能主要用于光化學反應(yīng)、熒光、熱耗散,本研究發(fā)現(xiàn),海島棉新海22號和陸地棉589光化學熒光淬滅系數(shù)qP差異不顯著,正反交F1沒有表現(xiàn)出雜種優(yōu)勢,陸海雜交F2的qP顯著降低,但其電子傳遞能力顯著高于親本和F1,說明雖然光合電子傳遞能力較強,但其吸收的光能用于光化學反應(yīng)的比例較低。NPQ=(Fm-Fm′)/Fm′=Fm/Fm′-1[25]反映了植物耗散過剩光能為熱的能力,是衡量植物光保護能力的重要指標。589的NPQ值顯著高于海島棉,說明其可將過剩光能轉(zhuǎn)化為熱耗散,避免強光損傷,進行自我保護的能力較強,而陸海雜交后代則差異不顯著,也沒有雜種優(yōu)勢。Y(NO)[26]是指除了熱擴散以外的熒光淬滅部分,若Y(NO)較高,表明入射光強超過了植物能接受的程度。這時,植物可能已經(jīng)受到損傷,或者(盡管還未受到損傷)繼續(xù)照光的話植物將要受到損傷,是光損傷的重要指標。陸地棉589的Y(NO)顯著高于海島棉,一方面說明其耗散的過剩光能較多,另一方面也說明其受到強光損傷的幾率也在增大,陸海雜交后代的Y(NO)值介于陸地棉和海島棉之間,且沒有表現(xiàn)出雜種優(yōu)勢。Y(NPQ)[26]與光保護機制相關(guān)的所有熱擴散,若Y(NPQ)較高,一方面表明植物接受的光強過剩,另一方面則說明植物仍可以通過調(diào)節(jié)(如將過剩光能耗散為熱)來保護自身,是光保護的重要指標,陸地棉的Y(NPQ)值顯著強于海島棉,說明其進行光保護的能力高于海島棉,當強光照射時,可將過剩光能轉(zhuǎn)化為熱擴散的能力高于海島棉,從另外一個方面可說明陸地棉適應(yīng)環(huán)境的能力較強,陸海雜種后代沒有表現(xiàn)出雜種優(yōu)勢。Fv/Fm=(Fm-Fo)/Fm是PSⅡ[23]的最大量子效率,反映植物潛在最大光合能力,高等植物一般在 0.8~0.84 之間,當植物受到脅迫(stress)時,F(xiàn)v/Fm顯著下降,陸地棉Fv/Fm值顯著低于海島棉,說明海島棉的最大光合能力顯著高于陸地棉,陸海雜種后代的Fv/Fm介于陸地棉和海島棉之間,也沒有表現(xiàn)出明顯的雜種優(yōu)勢。
參考文獻:
[1]錢能. 陸地棉遺傳多樣性與育種目標性狀基因(QTL)的關(guān)聯(lián)分析[D]. 南京:南京農(nóng)業(yè)大學,2009.
[2]張小全,王學德. 細胞質(zhì)雄性不育陸地棉與海島棉間雜種優(yōu)勢初步研究[J]. 棉花學報,2005,17(2):79-83.
[3]菲力申. 陸地棉與海島棉雜種優(yōu)勢利用研究[D]. 南寧:廣西大學,2008.
[4]常俊香. 棉花海陸雜交主要經(jīng)濟性狀的遺傳與雜種優(yōu)勢研究[D]. 鄭州:河南農(nóng)業(yè)大學,2007.
[5]石玉真,劉愛英,李俊文,等. 陸海種間雜交纖維品質(zhì)性狀的遺傳及其F1群體優(yōu)勢分析[J]. 棉花學報,2008,20(1):56-61.
[6]李哲,楊金玉,崔秀珍,等. 棉花海陸雜交種高支紗纖維特征與應(yīng)用研究[J]. 河南農(nóng)業(yè)科學,2007(6):49-53.
[7]鄧仲篪,孫濟中,張金發(fā),等. 雜交棉及其親本光合特性的研究[J]. 華中農(nóng)業(yè)大學學報,1995,14(5):429-434.
[8]姜磊,王旺華,李廷春,等. 棕色棉與白色棉雜交F1代吐絮期光合特性的雜種優(yōu)勢研究[J]. 棉花學報,2011,23(4):323-328.
[9]伍維模,董合林,危常洲,等. 南疆陸地棉與海島棉光合-光響應(yīng)及葉綠素熒光特性分析[J]. 西北農(nóng)業(yè)學報,2006,15(4):141-146.
[10]高俊鳳. 葉綠素含量定量測定[M]. 北京:高等教育出版社,2006:74-77.
[11]Jiang Y J,Zheng D M,Lu S Q,et al. Salt of soil affect quality of cotton[J]. Journal of Tarlm Agricultural Reclanation University,1994,8(2):23-27.
[12]張小全,王學德. 細胞質(zhì)雄性不育陸地棉與海島棉間雜種優(yōu)勢初步研究[J]. 棉花學報,2005,17(2):79-83.
[13]許大全. 光合速率影響因素[M]. 上海:上海科學技術(shù)出版社,2002:46-47.
[14]Kount R,Ilik P,Nans J,et al. On the limits of applicability of spectrophotometer and spectrofluorime tricmethods for the determination of chlorophyll a/b ratio[J]. Photosynthesis Research,1999,62(5):107-116.
[15]李丹丹,司龍亭,李季,等. 弱光下黃瓜幼苗葉片葉綠素含量的遺傳分析[J]. 華北農(nóng)學報,2009,24(1):133-137.
[16]尚莉,武繼承,管秀娟,等. 河南主栽小麥品種葉綠素含量的變化特征[J]. 河南農(nóng)業(yè)科學,2009(2):25-27.
[17]秦鴻德,張?zhí)煺?棉花葉綠素含量和光合速率的QTL定位[J]. 棉花學報,2008,20(5):394-398.
[18]支金虎,伍維模,危常洲,等. 水分與氮素對膜下滴灌棉花葉片葉綠素含量時空分布的影響[J]. 西北農(nóng)業(yè)學報,2007,16(1):7-12.
[19]張亞黎,姚賀盛,羅 毅,等. 海島棉和陸地棉葉片光合能力的差異及限制因素[J]. 生態(tài)學報,2011,31(7):1803-1810.
[20]Farquhar G D,Sharkeyt D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Biology,1982,33:317-345.
[21]Chen Z H,Wu F B,Wang X D,et al. Heterosis in CMS hybrids of cotton for photosynthetic and chlorophyll fluorescence parameters[J]. Euphytica,2005,144(3):353-361.
[22]劉金山,王有武,高山,等. 陸海雜交棉葉綠素熒光參數(shù)及生理動態(tài)變化研究[J]. 種子,2012,31(6):4-9.
[23]Krause G H,Weis E. Chlorophyll fluorescence and photosynthesis:the basis[J]. Plant Physiol,Plant Mol,Biol,1991,42:313-349.
[24]呂曉菡,柴偉國.低溫弱光下不同起源地辣椒幼苗光合特性的比較研究[J]. 浙江農(nóng)業(yè)學報,2014,26(1):48-53.
[25]van Kooten O,Snel J F. The use of chlorophyll fluorescence nomenclature in plant stress physiology[J]. Photosynthesis Research,1990,25(3):147-150.
[26]Kramer D M,Johnson G,Kiirats O,et al. New fluorescence parameters for the determination of q(a) redox state and excitation energy fluxes[J]. Photosynthesis Research,2004,79(2):209-218.索歡,陳龍正,徐海,等. 小白菜根腫病接種鑒定研究[J]. 江蘇農(nóng)業(yè)科學,2016,44(5):193-195.
