特有植物厚殼紅瘤果茶自然種群生命表及生存力分析
2016-07-23 15:59:31袁叢軍楊冰戴曉勇
江蘇農業科學 2016年5期
袁叢軍++楊冰++戴曉勇

摘要:為揭示貴州省特有種厚殼紅瘤果茶(Camellia rubituberculata)種群結構與分布格局特征,基于模式產地及集中分布區野外踏查資料,選取天然種群集中分布的地段,采用相鄰格子法設置典型群落樣地,對比分析不同干擾生境下厚殼紅瘤果茶自然種群的胸徑結構特征,制定靜態生命表并對生存力進行分析。結果表明,興仁縣和晴隆縣厚殼紅瘤果茶萌生、實生種群數量整體上均呈現隨著徑級結構增大逐漸減小;興仁縣種群以實生種群為主(87.4%),而萌生種群在晴隆縣分布地占據重要比重(45.5%);XRM、QLM、XRT種群呈增長型,而XRS、QLS、QLT為衰退型種群。除QLS種群存活曲線呈 Deevey-Ⅲ型外,其余植株種群存活曲線均呈Deevey-Ⅱ型;各種群存活曲線類型方程檢驗解釋量為XRS>QLS>XRM>QLM;XRM、QLS種群死亡率和虧損率均呈現逐漸降低趨勢,且降低速率表現為XRM>QLS;QLM、XRS種群死亡率和虧損率均呈現逐漸升高趨勢,且升高速率表現為QLM>XRS。種群的生存率總體上逐漸下降,隨后保持平穩趨勢,不同干擾強度與起源的下降速率及平穩點有所不同。其中,XRM種群到Ⅴ齡級時、QLS種群到達Ⅵ齡級時、晴隆縣萌生種群到達Ⅸ齡級時基本達到平穩,而興仁縣實生種群并無明顯的平穩點;種群累積死亡率[F(t)]與種群生存率[S(i)]相反。各種群的平均死亡密度率[f(t)]呈現QLM(0.090)>XRS(0.089)>QLS(0.084)>XRM(0.078),而平均危險率[λ(t)]則呈現QLS(0.372)>XRM(0.335)>QLM(0.323)>XRS(0.265)。
關鍵詞:厚殼紅瘤果茶;自然種群;靜態生命表;存活曲線;貴州省
中圖分類號: S685.140.1文獻標志碼: A文章編號:1002-1302(2016)05-0275-06
厚殼紅瘤果茶(Camellia rubituberculata)是山茶科(Theaceae)山茶屬(Camellia)常綠小喬木,高度可達8 m,1984年發現于晴隆縣,為貴州省特有種[1],主要分布于晴隆縣和興仁縣,現有資源量極少,僅分布于貴州省興仁縣田灣鄉新寨村和晴隆縣紫馬鄉上捧碧,分布面積共計超過 46.6 hm2。該物種性喜光,葉常綠、花大且花期長(花期10月至翌年3月)、樹形優美,可作為優良鄉土園林植物優選;木材堅硬、細致,可作為農具、細木工等優良用材優選[2];其葉、油、花在化妝品領域有重要的科研價值[3];因其自然生境極為特殊,該樹種適生于喀斯特石山地,可作為石漠化區植被恢復生態經濟型鄉土樹種。然而,有關該種的研究并引起足夠的關注,尤其對厚殼紅瘤果茶天然種群生命表及生存分析尚未見報道。本研究以典型厚殼紅瘤果茶天然群落為研究對象,采用種群生命表及生存分析理論對厚殼紅瘤果茶天然種群進行調查研究,揭示該區域厚殼紅瘤果茶種群的數量動態變化特征、種群變化趨勢,從而為野生厚殼紅瘤果茶天然群落的保護、該種資源的示范推廣以及在退化生態系統適宜地段上的人工林培育開發利用提供科學的理論依據和指導。
種群不僅是連接群落與個體之間的紐帶,也是生物群落、生態系統的基本組成成分。對植物種群研究的最終目的是要闡明種群的動態分化與適應的過程[4]。種群的數量特征是種群的動態分化研究核心內容,包括種群數量、密度、年齡、性比等種群數量統計基本參數,可以反映種群在空間上的變動規律,從而闡釋種群動態及其所在群落的演替趨勢[5]。種群結構及其變化規律一直是植物生態學的研究熱點之一,種群動態研究是種群生態學的核心。植物種群靜態生命表和存活曲線的刻畫,能夠揭示種群生活史中的生死規律和期望壽命,可以為分析珍稀瀕危植物種群的保護和珍貴植物速生期利用提供科學依據。通過對種群生命表的編制可得出死亡率、損失率等重要參數,從而為種群數量統計提供更多信息。因此,開展植物種群數量與動態變化規律的研究,對種群的保育管護、預估種群變化趨勢和種群資源的開發利用具有重要意義。
1材料與方法
1.1研究區概況
研究區位于貴州省興仁縣田灣鄉(新寨村石峩廠組石峩廠)和晴隆縣紫馬鄉(龍頭村上捧碧),屬于中亞熱帶季風氣候,雨熱同期。年均氣溫為13.9~15.2 ℃,≥10 ℃積溫 4 098.0~4 531.2 ℃,極端最低氣溫為-6.2 ℃,最高氣溫為21.7 ℃,年日照時間1 513.2 h,無霜期281~290 d,年均降水量1 323.7~1 577.2 mm,4—9月降水量 1 065.1~1 249.7 mm,占全年降水量的80.0%。該區屬于喀斯特與砂頁巖鑲嵌的丘陵地貌,生境為石灰巖山地,巖石裸露率70%~90%。群落為常綠落葉闊葉混交林,其中喬木層主要植物有星毛石櫟(Lithocarpus petelotii)、青岡櫟(Cyclobalanopsis glauca)、厚殼紅瘤果茶(Camellia rubituberculata)、皂莢(Gleditsia sinensis)、云南樟(Cinnamomum glanduliferum)、川黔潤楠(Machilus chuanchienensis)、黃連木(Pistacia chinensis)等; 灌木層植物有厚殼紅瘤果茶、清香木(Pistacia weinmanifolia)、火棘(Pyracantha fortuneana)、長葉凍綠(Rhamnus crenata)、齒葉鐵仔(Myrsine semiserrata)、尖瓣瑞香(Daphne acutiloba);主要藤本植物有大葉酸藤子(Embelia subcoriacea)、杠柳(Periploca sepium)、山橙(Melodinus suaveolens)等;草本植物分布較少。
1.2研究方法
1.2.1野外調查方法在野外實地踏查的基礎上,選取特有植物厚殼紅瘤果茶種群天然分布區貴州省興仁縣田灣鄉、晴隆縣紫馬鄉天然群落為研究對象,分別設置具代表性的樣地(30 m×30 m) 3個,共12塊樣地,總樣地面積2 700 m2 。記錄每個樣地的海拔、坡向、坡度等指標,對樣地內物種進行每木調查,調查各物種株高、地徑(胸徑)、冠幅、樹高、枝下高等指標,同時在每個樣地中等距設置6個2 m×2 m樣方進行灌木層調查、6個1 m×1 m樣方進行草本層調查(表1)。
1.2.2齡級劃分為避免破壞野生植物資源,本研究采用“空間替代時間”的方法,即將林木依胸徑大小分級,以立木級結構代替年齡結構分析種群動態。關于種群大小結構立木級的劃分標準,許多學者提出了不同的標準[6-10]。根據該種的生物學特性,本試驗以灌叢為單株,采用植株胸徑(DBH)作為個體大小的指標研究種群大小結構。徑級劃分方法:Ⅰ級(DBH<1.0 m)、Ⅱ級(1.0≤DBH<2.0 cm)、Ⅲ級(2.0≤DBH<3.0 cm)、Ⅳ級(3.0≤DBH<4.0 cm)、Ⅴ級(4.0≤DBH<5.0 cm)、Ⅵ級(5.0≤DBH<6.0 cm)、Ⅶ級(6.0≤DBH<7.0 cm)、Ⅷ級(7.0≤DBH<8.0 cm)、Ⅸ級(8.0≤DBH<9.0 cm)。采用陳曉德的數學模型,推導種群年齡結構的動態指數對厚殼紅瘤果茶種群動態進行定量描述[11]。
1.2.3生命表編制及生存函數計算由于靜態生命表反映多個世代重疊的年齡動態歷程中的一個特定時間,而不是對這一種群的全部生活史的追蹤,并且調查中存在系統誤差,在生命表中會出現死亡率為負的情況,因此本研究采用勻滑技術對數據進行處理。采用生存率函數S(t)、累積死亡率函數F(t)、死亡密度函數f(t)和危險率函數λ(t)等指標對厚殼紅瘤果茶自然種群的生存規律及其種群動態進行分析[12]。
2結果與分析
2.1種群起源及年齡結構特征
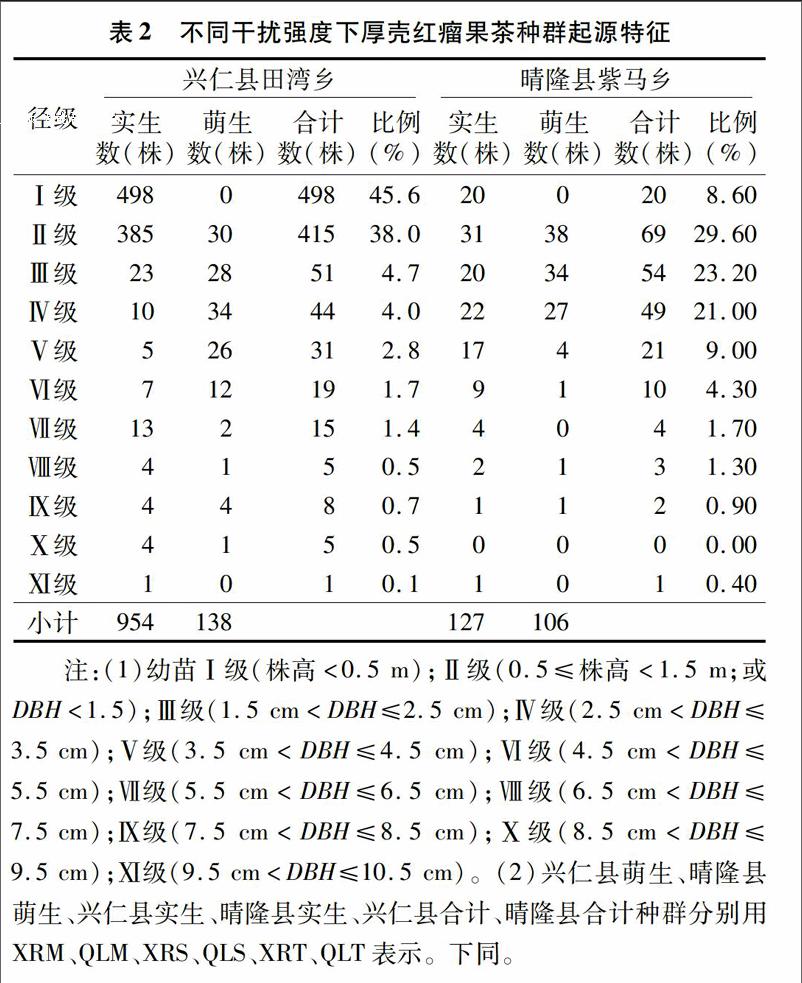
對不同干擾強度下厚殼紅瘤果茶種群起源特征統計發現,興仁縣田灣鄉厚殼紅瘤果茶種群數量顯著高于晴隆縣紫馬鄉;其中田灣鄉分布的Ⅰ級、Ⅱ級實生幼苗共有883株,萌生幼苗30株,占總植株數的83.6%;而紫馬鄉分布的Ⅰ級、Ⅱ級實生幼苗共有51株,萌生幼苗38株,占總植株數的 38.2%。不同徑級統計表明,實生苗呈現954株(田灣鄉)>127株(紫馬鄉)、萌生苗呈現138株(田灣鄉)>106株(紫馬鄉),而萌生 ∶實生則呈現0.835(紫馬鄉)>0.145(田灣鄉)。研究結果表明,田灣鄉(干擾較弱,生境保護水平較好)苗木種子繁育較好,而紫馬鄉較弱(表2)。
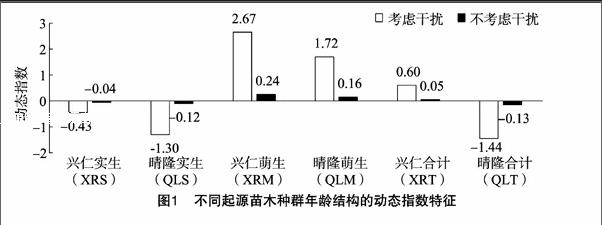
種群的齡級結構是指各齡級組的個體在種群內的比例和配置情況。總體看來,興仁縣和晴隆縣厚殼紅瘤果茶萌生、實生種群數量均呈現隨著徑級結構增大逐漸減小。基于量化分析方法繪制不同厚殼紅瘤果茶種群齡級結構的動態指數(Vpi)如圖1所示,可知:(1)考慮干擾情況下,種群齡級結構的動態指數大小依次為XRM種群(2.67)>QLM種群(1.72)>XRT種群(0.60)>XRS種群(-0.43)>QLS種群(-1.30)>QLT種群(-1.44);(2)不考慮干擾情況,種群齡級結構的動態指數大小依次為XRM種群(0.24)>QLM種群(0.16)>XRT種群(0.05)>XRS種群(-0.04)>QLS種群(-0.12)>QLT種群(-0.13);研究結果表明,XRM、QLM、XRT種群呈增長型,而XRS、QLS、QLT為衰退型種群。
2.2不同種群靜態生命表特征
靜態生命表是根據一定時期樣地內一個種群所有個體的年齡數據編制而成[13],不僅可以反映種群從出生到死亡的數量動態,還可用于預測種群未來發展的趨勢。以生存分析理論為基礎,編制不同干擾強度下不同起源厚殼紅瘤果茶種群靜態生命表(表3)。
由表 3可以看出, 不同干擾下實生和萌生種群均呈現種
群數量隨著齡級結構的增加逐漸減小,而萌生種群均缺乏Ⅰ齡級種群植株。ex反映了x齡級內個體的生命期望壽命,研究發現,XRS與QLM幼年階段(Ⅰ~Ⅲ級)表現出高的生命期望,說明此階段厚殼紅瘤果茶植株種群的生存質量較高、生理活動旺盛;隨著齡級的增加,植株間的競爭和其他人為干擾影響導致高齡級的生命期望逐漸減小。
2.3不同種群存活曲線特征
存活曲線借助于存活個體數量以描述特定年齡的存活率,反映種群個體在各年齡級的存活狀況[14]。本研究以標準化存活量lx為縱坐標、以徑級相對的齡級作橫坐標,繪制了存活曲線(表4)。由此可知,除晴隆縣實生植株種群存活曲線呈Deevey-Ⅲ型外,其余植株種群存活曲線均呈Deevey-Ⅱ型;各不同起源厚殼紅瘤果茶植株種群存活曲線類型方程檢驗解釋量依次為興仁縣實生(R2=0.963)>晴隆縣實生(R2=0.935)>興仁縣萌生(R2=0.829)>晴隆縣萌生(R2=0.815)。
2.4死亡率和虧損率曲線
以厚殼紅瘤果茶不同種群的齡級為橫坐標,死亡率(qx)和虧損率(Kx)為縱坐標,繪制該種群的死亡率和虧損率曲線(圖2)。由圖 2可知,厚殼紅瘤果茶各種群的死亡率和虧損率曲線的變化基本一致。總體看來,XRM、QLS種群死亡率和虧損率均呈現逐漸降低趨勢,且降低速率表現為XRM>QLS;QLM、XRS種群死亡率和虧損率大致呈現逐漸升高趨勢,且升高速率表現為QLM>XRS。
2.5種群生存函數分析
厚殼紅瘤果茶植株種群的生存率總體上逐漸下降,隨后保持平穩趨勢,不同干擾強度與起源的下降速率及平穩點有所不同(圖3)。其中,興仁縣萌生種群到Ⅴ齡級時、晴隆縣實生種群到達Ⅵ齡級時、晴隆縣萌生種群到達Ⅸ齡級時基本達到平穩,而興仁縣實生種群并無明顯的平穩點;與之相應,種群累積死亡率與種群生存率相反。各種群的平均死亡密度呈現QLM(0.090)>XRS(0.089)>QLS(0.084)>XRM(0078),而平均危險率則呈現QLS(0.372)>XRM(0.335)>QLM(0.323)>XRS(0.265)。XRM種群死亡密度表現為先保持不變(Ⅰ~Ⅲ級),后迅速減小并保持不變(Ⅴ~Ⅸ級),隨后在Ⅹ級處出現小波峰;QLM種群死亡密度則表現為先在Ⅱ級處出現小波峰,后保持不變(Ⅲ~Ⅷ級),后逐漸迅速減小并在Ⅹ級處又出現小波峰;XRS種群死亡密度表現為先保持不變(Ⅰ~Ⅲ級),隨后即在Ⅳ處出現小波峰,后保持不變(Ⅳ~Ⅸ級),隨后在Ⅹ級處出現小波峰;QLS種群死亡密度則表現為先保持不變(Ⅰ~Ⅲ級),后先減小后迅速增
加,并在Ⅴ級處出現小波峰,后保持不變(Ⅵ~Ⅸ級),隨后在Ⅹ級處出現小波峰。而XRM種群危險率表現為先增加后減小,再保持平穩不變(Ⅵ~Ⅹ級);QLM種群危險率表現為先增加后減小;XRS種群危險率表現為先增加后減小,隨后又
增加趨勢;QLS種群則表現為先增加后減小,后又呈現先增加后減小,最后又再次呈現增加趨勢。
3結論與討論
3.1結論
興仁縣和晴隆縣厚殼紅瘤果茶萌生、實生種群數量均隨著徑級結構增大逐漸減小;其中,興仁縣種群以實生種群(種子繁育)為主(87.4%),而萌生種群在晴隆縣分布地占據重要比重(45.5%);XRM、QLM、XRT種群呈增長型,而XRS、QLS、QLT為衰退型種群。
除QLS種群存活曲線呈Deevey-Ⅲ型外,其余植株種群存活曲線均呈Deevey-Ⅱ型;各種群存活曲線類型方程檢驗解釋量為XRS>QLS>XRM>QLM;XRM、QLS種群死亡率和虧損率均呈現逐漸降低趨勢,且降低速率表現為XRM>QLS;QLM、XRS種群死亡率和虧損率均呈現逐漸升高趨勢,且升高速率表現為QLM>XRS。
厚殼紅瘤果茶植株種群的生存率總體上逐漸下降,隨后保持平穩趨勢,不同干擾強度與起源的下降速率及平穩點有所不同。其中,興仁縣萌生種群到Ⅴ齡級時、晴隆縣實生種群到達Ⅵ齡級時、晴隆縣萌生種群到達Ⅸ齡級時基本達到平穩,而興仁縣實生種群并無明顯的平穩點;與之相應,種群累積死亡率與種群生存率相反。
3.2討論
植物地理學研究中,常常將分布狹限或是異常狹限分布區的物種或其他分類單元稱為特有植物[1]。貴州省特有植物是僅分布在貴州省地理區域內,而其他地區沒有的植物種類(目前未見報道或發現),其起源、發育演化和傳播呈間斷或不間斷狹限分布在貴州省地理區域內。山茶屬植物是貴州省特有植物種類最多的屬,約分布有16或21種[15-16],作為貴州省特有植物(280種)的重要組成部分,其表征著貴州省特殊生境條件下植物適應進化演變的結果。貴州省特有植物的保護與利用作為區域生物多樣化保護過程中一項極其重要研究內容和熱點,厚殼紅瘤果茶隸屬山茶科山茶屬瘤果茶組常綠小喬木,是典型的生長于巖溶區山旮旯地段鄉土適生油脂兼園林觀賞植物;分布區狹窄,資源量較少,多散生于常綠落葉闊葉林中,群落建群種以殼斗科植物為主,該種為伴生種[17-18]。因此,開展以野生自然種群動態變化和現有種群生存力分析具有重要意義。
種子繁育(實生苗)和萌蘗繁育(萌生苗)作為種子植物種群天然更新的2種重要方式,種群采取何種生存對策取決于該物種的生物學特性和外界環境的壓力影響[19-20]。前人對特有植物種群動態變化開展了許多研究工作[19,21-24],而從種群不同起源方式及干擾型對種群自然更新研究工作較少[25-26]。本研究對瘤果茶組(Sect. tuberculate)貴州省特有典型生態經濟型厚殼紅瘤果茶實生與萌生植株種群特征進行研究表明,該種種群繁育以實生苗為主(XRS、QLS所占比例分別為87.4%、54.5%),QLM種群數量是QLT種群數量的重要組成部分(45.5%)。晴隆縣紫馬鄉分布點由于種群資源量少,資源分布面積較小,且群落及生境地受人為干擾破壞較嚴重,萌生植物種群所占比例較大,成為該地厚殼紅瘤果茶種群更新繁育的重要組成部分;興仁縣田灣鄉分布點由于種群資源量較大,資源分布面積較大,且群落及生境地受人為干擾破壞較弱,實生植株種群占據了大量的比重(87.4%),人為干擾對種群更新,尤其是特有植物的種群更新具有顯著影響。研究結果與袁叢軍等對貴州省特有植物巖生紅豆研究結果一致,強干擾型顯著影響了目的樹種的種群更新與繁育[27]。然而有關強干擾型如何影響厚殼紅瘤果茶種群更新生態學過程的研究有待進一步深入和開展。
植物種群的生命表繪制和編制,反映該種群生存現狀,對揭示植物種群與環境適應的結果和闡明種群或群落未來發展趨勢具有重要意義[28]。由于現有科研試驗條件限制,要完全追蹤種群從發生在最后一個個體死亡的真實過程,獲取植物種群動態生命表是一項極其困難的工作,尤其是針對一些喬木長壽命植物種群而言[29]。生存分析理論用于研究種群的動態和結構變化,在生命表分析中具有較高的實際應用價值,能更合理、直觀地說明種群的結構和動態變化[30-31]。本研究鑒于該種群的自身生物學特性和生境特征,結合前人有關山茶屬植物研究結果[32-33],研究采用以1 cm為1個徑級單位對不同種群劃分齡級,編制不同起源和干擾情況下的植物種群生命表[34],并對各種群進行生存力分析。研究結果表明,QLS種群存活曲線呈Deevey-Ⅲ型(凹曲線型),說明晴隆縣實生種群在幼苗早期死亡率高,但一旦活至某一年齡,種群個體死亡率就會處于較低水平;而QLM、XRS、XRM種群存活曲線均呈Deevey-Ⅱ型(直線型),該類種群各年齡的死亡率基本一致。結合研究區的人為干擾活動情況,研究認為強干擾型的人為活動極大促進厚殼紅瘤果茶實生植株種群的幼齡(Ⅱ級→Ⅲ級)苗木的死亡數量及速率,對萌生植株種群(QLM種群死亡數量和速率)影響不顯著;弱干擾型的人為活動對XRM植株種群(Ⅰ級→Ⅵ級)影響顯著強于XRS種群。因此,建立保護小區并減少人為活動強度對厚殼紅瘤果茶野生種群的干擾,可有效保護野生實生種群,豐富和保持更多的遺傳多樣性。
參考文獻:
[1]林蒙嘉,盧其明. 貴州山茶新紀錄[J]. 中山大學學報:自然科學版,1984(2):83-85.
[2]鄧忠治,潘德權,劉正本,等. 貴州省厚殼紅瘤果茶初步研究[J]. 貴州林業科技,2011(2):39-42.
[3]周麗,王苑,張應祝,等. 厚殼紅瘤果茶果實特征與種子萌發特性[J]. 北方園藝,2015(2):34-37.
[4]姜漢僑,段昌群,楊樹華,等. 植物生態學[M]. 北京:高等教育出版社,2010.
[5]張金屯,吳冬麗,張斌,等. 北京地區幾種重點保護植物生存群落生態學研究[M]. 北京:中國科學技術出版社,2013.
[6]張治國,王仁卿. 中國分布北界的山茶 (Camellia japonica)種群大小結構和空間格局分析[J]. 植物生態學報,2000(1).
[7]洪偉,王新功,吳承禎,等. 瀕危植物南方紅豆杉種群生命表及譜分析[J]. 應用生態學報,2004(6):1109-1112.
[8]茹文明,張桂萍,畢潤成,等. 瀕危植物脫皮榆種群結構與分布格局研究[J]. 應用與環境生物學報,2007(1):14-17.
[9]閆淑君,洪偉,吳承禎,等. 絲栗栲種群生命過程及譜分析[J]. 應用與環境生物學報,2002(4):351-355.
[10]吳承禎,洪偉,謝金壽,等. 珍稀瀕危植物長苞鐵杉種群生命表分析[J]. 應用生態學報,2000(3):333-336.
[11]陳曉德. 植物種群與群落結構動態量化分析方法研究[J]. 生態學報,1998(2):104-107.
[12]盧杰,潘剛,羅大慶,等. 瀕危植物急尖長苞冷杉種群生命表分析[J]. 水土保持研究,2010(5):212-216.
[13]吳承禎,洪偉,謝金壽,等. 珍稀瀕危植物長苞鐵杉種群生命表分析[J]. 應用生態學報,2000(3):333-336.
[14]李玲,張光富,王銳,等. 天目山自然保護區銀杏天然種群生命表[J]. 生態學雜志,2011(1):53-58.
[15]屠玉麟. 貴州特有植物初步研究(一)[J]. 貴州林業科技,1991(3):68-81.
[16]張華海,班平原. 貴州山茶屬植物地理分布及開發利用[J]. 貴州科學,2007(1):68-72.
[17]鄒天才,劉海燕,周洪英,等. 貴州重要野生觀賞植物的引種繁殖和應用栽培技術研究[J]. 種子,2007,26(6):12-15,20.
[18]鄧忠治,潘德權,劉正本,等. 貴州省厚殼紅瘤果茶初步研究[J]. 貴州林業科技,2011(2):39-42.
[19]剛群,閆巧玲,劉煥彬,等. 種子更新與萌蘗更新蒙古櫟一年生幼苗生長特性的比較[J]. 生態學雜志,2014(5):1183-1189.
[20]李小雙,彭明春,黨承林. 植物自然更新研究進展[J]. 生態學雜志,2007(12):2081-2088.
[21]龔濱,夏洋潔,張光富,等. 中國特有珍稀瀕危樹種銀縷梅種群結構和空間格局[J]. 生態與農村環境學報,2012,28(6):638-646.
[22]秦鳳,鄧洪平,由永飛,等. 重慶市特有植物縉云衛矛的種群動態[J]. 貴州農業科學,2011(4):27-31.
[23]袁春明,孟廣濤,方向京,等. 珍稀瀕危植物長蕊木蘭種群的年齡結構與空間分布[J]. 生態學報,2012(12):3866-3872.
[24]楊乃坤,鄒天才,劉海燕,等. 貴州特有植物長柱紅山茶種群年齡結構及空間分布格局研究[J]. 熱帶亞熱帶植物學報,2015(2):205-210.
[25]王賽宵,李清河,徐軍,等. 克隆植物唐古特白刺不同層次的種群結構與生命表分析[J]. 東北林業大學學報,2013,41(1):72-76.
[26]郭有燕,周建云,何景峰,等. 文冠果種群繁殖方式及其在種群更新中的作用[J]. 應用生態學報,2014(11):3110-3116.
[27]袁叢軍,安明態,嚴令斌,等. 兩種干擾強度下巖生紅豆天然群落特征及物種多樣性分析[J]. 南方農業學報,2013(2):280-284.
[28]張悅,易雪梅,王遠遐,等. 采伐對紅松種群結構與動態的影響[J]. 生態學報,2015(1):38-45.
[29]何亞平,費世民,蔣俊明,等. 不同齡級劃分方法對種群存活分析的影響——以水災跡地油松和華山松種群生存分析為例[J]. 植物生態學報,2008(2):448-455.
[30]洪偉,王新功,吳承禎,等. 瀕危植物南方紅豆杉種群生命表及譜分析[J]. 應用生態學報,2004,15(6):1109-1112.
[31]李建貴,潘存德,彭世揆,等. 天山云杉種群統計與生存分析[J]. 北京林業大學學報,2001(1):84-86.
[32]詹明勛,何坤益,徐震騏,等. 臺灣中部暖溫帶闊葉林年輪特征、生長與樹齡結構[J]. 北京林業大學學報,2015(3):84-93.
[33]杜興喬,劉映良. 荔波瘤果茶種群生命表分析[J]. 安徽農業科學,2009(33):16613-16615.
[34]謝佳燕,林佳. 亞致死濃度吡蟲啉對禾谷縊管蚜實驗種群生命表參數的影響[J]. 江蘇農業科學,2014,42(5):125-126.何遵衛,周章義,馬鳳翔. 4個頻率下不同生長等級國槐的電生理分析[J]. 江蘇農業科學,2016,44(5):281-283.
