黃顙魚、烏蘇里擬鲿雜交子代胚胎發育和胚后30d的體長變化
2016-07-23 16:21:47董少杰孫潔畢相東
江蘇農業科學 2016年5期
董少杰++孫潔++畢相東

摘要:研究了黃顙魚(Pelteobagrus fulvidraco)×烏蘇里擬鲿(Pseudobagrus ussuriensis)雜交卵的受精率、孵化率、畸形率、胚胎發育時序以及魚苗破膜后30 d內的體長變化。結果表明,黃顙魚×烏蘇里擬鲿正交組和烏蘇里擬鲿×黃顙魚反交組卵的受精率和畸形率與兩親本自交組并無顯著差異,黃顙魚×烏蘇里擬鲿正交組與黃顙魚自交組受精卵的孵化率無顯著差異,且均顯著高于烏蘇里擬鲿×黃顙魚反交組和烏蘇里擬鲿自交組。兩雜交組胚胎發育時序分別與各自母本自交組相近。胚后30 d內,烏蘇里擬鲿×黃顙魚反交組和烏蘇里擬鲿自交組魚苗在體長方面較其他試驗組具有明顯優勢,且前者在破膜后第20天開始,體長顯著超過后者。
關鍵詞:黃顙魚;烏蘇里擬鲿;雜交;胚胎發育;體長
中圖分類號: S961.5文獻標志碼: A文章編號:1002-1302(2016)05-0313-03
黃顙魚(Pelteobagrus fulvidraco)、烏蘇里擬鲿(Pseudobagrus ussuriensis)均隸屬于鲇形目鲿科,分屬黃顙魚屬、擬鲿屬。鲿科魚類大部分曾是江蘇省境內江河湖泊水域的常見經濟魚類[1],其中黃顙魚屬有黃顙魚(P. fulvidraco)、瓦氏黃顙魚(P. vachelli)、長須黃顙魚(P. eupogon)、中間黃顙魚(P. intermedius)、光澤黃顙魚(P. nitidus)[2]。黃顙魚因其肉質細嫩、味道鮮美、無肌間刺且營養豐富,深受消費者喜愛[3],是我國北方地區廣泛養殖的重要淡水經濟魚類。黃顙魚的生長速度比較慢,常見個體質量100~200 g,250 g以上即為大個體,即使在人工飼養條件下長到125 g左右也需要2年,且其雌魚個體規格遠小于同齡雄魚[3]。利用親緣關系超過種內關系的親本進行遠緣雜交往往能夠獲得具有雜種優勢的群體[4]。近年來,為了改良黃顙魚的生長性能,國內有學者利用黃顙魚和同屬魚類作為親本進行了雜交育種,如黃顙魚(♀)×瓦氏黃顙魚(♂)[5],黃顙魚(♀)×江黃顙魚(♂)[6],雜交卵的受精率或出苗率均不低于自交卵。烏蘇里擬鲿俗稱牛尾巴,主要分布于我國黑龍江水系,體型較大,最大可達1 m,且肉質鮮美、細膩,無肌間刺[7]。邱叢芳等報道了黃顙魚(♀)×烏蘇里擬鲿(♂)的雜交試驗,指出雜交品種生長速度較快、成活率較高,該研究證明了黃顙魚(♀)×烏蘇里擬鲿(♂)雜交的可能性,沒有提及烏蘇里擬鲿(♀)×黃顙魚(♂)雜交,該研究使用了黃顙魚母本10尾,烏蘇里擬鲿父本1尾,共孵化魚苗543尾,沒有提供雜交卵的受精率、孵化率、畸形率等數據[8]。為了進一步確定黃顙魚×烏蘇里擬鲿雜交是否適合在生產中推廣應用,筆者在邱叢芳等研究[8]的基礎上,開展了進一步探索。
1材料與方法
1.1親魚來源
試驗用親魚均由天津藍科水產養殖有限公司提供。黃顙魚雌魚20尾,體質量(320±10) g,雄魚6尾,體質量(110±5) g;烏蘇里擬鲿雌魚20尾,體質量(200 g±17) g,雄魚6尾,體質量(302±15) g。試驗魚體格健壯,無外傷,性腺發育成熟度良好。
1.2試驗設計
試驗分為正交組(組合Ⅰ):黃顙魚(♀)(10尾)×烏蘇里擬鲿(♂)(3尾);反交組(組合Ⅱ):烏蘇里擬鲿(♀)(10尾)×黃顙魚(♂)(3尾);黃顙魚自交組(組合Ⅲ):黃顙魚(♀)(10尾)×黃顙魚(♂)(3尾);烏蘇里擬鲿自交組(組合Ⅳ):烏蘇里擬鲿(♀)(10尾)×烏蘇里擬鲿(♂)(3尾)。
1.3催產
催產激素選用絨毛膜促性腺激素(HCG)+促黃體素釋放激素類似物(LRH-A2)+馬來酸地歐酮(DOM)。注射劑量:HCG 3 000 IU/kg、LRH-A210 μg/kg、DOM 3 mg/kg。雌魚采用2次注射法,第一針注射催產激素總量的25%,第二針注射余下的催產激素,2次注射時間間隔12 h;雄魚一次注射,與雌魚第二次注射同步進行,注射劑量為雌魚的50%。第二次注射16 h后,達到效應時間。
1.4人工授精
采用濕法受精。進入效應期后,解剖雄魚,取出精巢。將精巢剪碎并置于燒杯中,用7‰生理鹽水稀釋精液。將雌魚卵子擠入另一干燥的燒杯中,加入精液稀釋液并立即快速、充分攪拌,使精卵充分接觸、受精。將受精卵置于聚乙烯網片上,放入孵化池中孵化。孵化溫度24.5~26.5 ℃,其間不間斷充氣。魚卵孵化后,將網片撤出。待魚苗體色由透明轉至淺黑色開始添加生物餌料,每天換水,保持水質清新。
1.5受精率、孵化率、出苗率、畸形率計算方法
采用王衛民等的方法[5]計算受精率、孵化率、出苗率、畸形率,并略作修改。從人工授精后的卵中抽取300粒卵,分為3個平行組,每組包括100粒卵。將每組卵分別放置在3個 5 000 mL 燒杯中靜水孵化,孵化過程中不間斷充氣,達到原腸晚期后計算每組卵的總數、受精卵,挑出受精卵,按組分別放入3個5 000 mL燒杯中繼續充氣孵化。魚苗破膜后計數魚苗總數、畸形魚苗數。計算公式如下:
受精率= 受精卵粒數檢查卵總數×100%;(1)
孵化率= 出膜魚苗數受精卵總數×100%;(2)
畸形率= 畸形魚苗數出膜魚苗總數×100%。(3)
1.6胚后發育
繁殖魚苗在養殖桶中培育,魚苗破膜48 h后,每隔1 d測量并記錄魚苗體長。
1.7數據處理
試驗數據用平均值±標準差表示。使用SPSS 13.0軟件對試驗數據進行方差分析,選用Duncan法進行多重比較,進行數據顯著性檢驗。使用Excel 2007軟件繪制圖形。
2結果與分析
2.1受精率、孵化率、畸形率
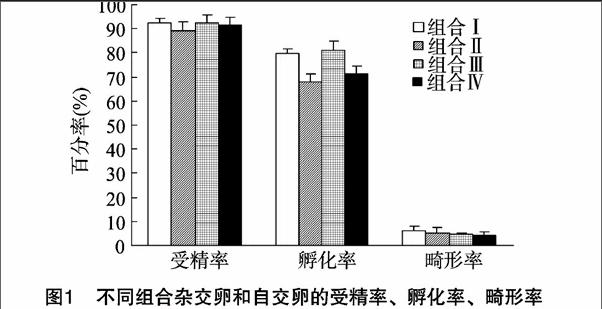
如圖1所示,黃顙魚×烏蘇里擬鲿正交、烏蘇里擬鲿×黃顙魚反交、黃顙魚自交、烏蘇里擬鲿自交的受精率分別為(92.67±1.53)%、(89.33±3.97)%、(92.33±3.51)%、(91.67±3.21)%,各試驗組間差異不顯著。黃顙魚×烏蘇里擬鲿正交試驗組與黃顙魚自交試驗組受精卵孵化率分別為(79.67±2.08)%、(81.33±3.51)%,二者之間差異不顯著,但均顯著高于黃顙魚×烏蘇里擬鲿反交試驗組和烏蘇里擬鲿自交試驗組。黃顙魚×烏蘇里擬鲿正交、烏蘇里擬鲿×黃顙魚反交、黃顙魚自交、烏蘇里擬鲿自交試驗組幼苗畸形率分別為(6.00±2.00)%、(5.33±2.08)%、(4.67±0.58)%、(4.33±1.53)%,各試驗組間差異不顯著。
2.2雜交卵和自交卵的胚胎發育
如表1所示,相同孵化條件下,黃顙魚自交卵發育時長最久(約65 h 15 s),黃顙魚×烏蘇里擬鲿正交卵次之(約60 h 10 s),烏蘇里擬鲿自交卵發育時長最短(約50 h 53 s),黃顙魚×烏蘇里擬鲿反交卵發育時長與烏蘇里擬鲿自交卵相近,較之略長(約51 h 23 s)。各試驗組受精卵的發育時序有差異,雜交組受精卵與母本自交受精卵發育時序相接近。
2.3胚后30 d內體長變化
如圖2所示,除了在孵化后第22天黃顙魚自交組與烏蘇里擬鲿自交組魚苗體長相近外,在孵化后第2天至第30天,黃顙魚×烏蘇里擬鲿反交組和烏蘇里擬鲿自交組魚苗體長均顯著高于黃顙魚×烏蘇里擬鲿正交組和黃顙魚自交組魚苗。孵化后第18天之前,黃顙魚×烏蘇里擬鲿反交組和烏蘇里擬鲿自交組魚苗體長差異不顯著,自孵化后第20天起直到試驗結束(第30天),前者魚苗體長極顯著大于后者。各試驗組魚苗出膜后30 d內的體長增長均呈典型的指數函數關系(表2)。
3結論與討論
3.1黃顙魚和烏蘇里擬鲿雜交在生產上的可行性
遠緣雜交是雜交育種中應用較多的一種獲得具有優良經濟性狀雜交后代的方式,它是指種間、屬間乃至親緣關系更遠的生物類型之間的雜交[6]。黃顙魚與烏蘇里擬鲿為同科異屬,前者生長速度較快,但個體較小,后者個體較大,對環境適應能力強,但生長速度相對較慢。王明寶等對黃顙魚(P. fulvidraco)與瓦氏黃顙魚雜交技術進行了研究,指出雜交后代生長速度快、形態特征好,當年可達到125 g/尾[3]。蔡永祥等研究了江黃顙和烏蘇里擬鲿雜交,得到了大量魚苗,指出二者的屬間雜交在育種實踐中是可行的[9]。本研究結果表明,黃顙魚與烏蘇里擬鲿正交組卵和反交組卵的受精率、畸形率與親本自交組并無顯著差異。孵化率方面,雜交組與母本自交組相近,證明了兩者屬間雜交在實踐中的可行性。黃顙魚的核型為24 m+14 sm+10 st+4 t[10],烏蘇里擬鲿的核型為24 m+18 sm+10 st[10-11],兩者二倍染色體數(2n)均為52,核型相近。王祖熊等認為,雙親間的染色體組型越相近,雜交親和性越強且成功概率也越高[12]。本研究結果符合這一觀點。
3.2雜交與自交卵胚胎發育和胚后30 d體長變化比較
本研究結果表明,黃顙魚與烏蘇里擬鲿正交組和反交組胚胎發育時序均介于父母本自交組之間。前者發育時長約為60 h 10 s,較母本自交組約提前5 h,較父本自交組約延遲9 h;后者發育時長約為51 h 23 s,較母本自交組約延遲0.5 h,較父本自交組約縮短14 h。雜交卵在胚胎發育過程中會發生“偏母遺傳”現象,即雜種的胚胎發育速度受到細胞質因子的調控,胚胎發育的時序介于父母本之間,并偏向于各自母本胚胎發育時間[7,13]。本研究結論與上述觀點相符。因條件所限,本研究僅觀測了魚苗破膜后30 d內的體長變化。結果表明,黃顙魚×烏蘇里擬鲿正交組魚苗與黃顙魚自交組體長差異不顯著。烏蘇里擬鲿×黃顙魚反交組和烏蘇里擬鲿自交組魚苗在體長方面較其他試驗組具有明顯優勢,且前者自破膜后第20天開始,體長顯著超過后者。
參考文獻:
[1]王明華,蔡永祥,陳校輝,等. 烏蘇里擬鲿、瓦氏黃顙魚雜交與自交子代生長性能比較[J]. 水產科學,2013,32(1):50-54.
[2]張國松,侯新遠,尹紹武,等. 黃顙魚屬(Pelteobagrus)魚類遺傳多樣性的研究進展[J]. 江蘇農業科學,2014,42(3):174-179.
[3]王明寶,陳強,陳耀炳,等. 黃顙魚與瓦氏黃顙魚雜交技術研究[J]. 現代農業科技,2012(24):273,278.
[4]樓允東. 魚類育種學[M]. 北京:中國農業出版社,2001.
[5]王衛民,嚴安生,張志國,等. 黃顙魚♀與瓦氏黃顙魚♂的雜交研究[J]. 淡水漁業,2002,32(3):3-5.
[6]樓允東,李小勤. 中國魚類遠緣雜交研究及其在水產養殖上的應用[J]. 中國水產科學,2006,13(1):151-158.
[7]秦欽,梁丹妮,王明華,等. 雜交鲿(烏蘇里擬鲿♀×瓦氏黃顙魚♂)胚胎發育的研究[J]. 南京師大學報:自然科學版,2012,35(3):81-86.
[8]邱叢芳,王彬,余祥勝. 黃顙魚♀×烏蘇里擬鲿♂苗種繁殖養殖試驗[J]. 科技信息,2009(4):314.
[9]蔡永祥,陳友明,陳校輝,等. 江黃顙(Pelteobagrus vachelli)和烏蘇里擬鲿(Pseudobagrus ussuriensis)雜交F1代形態差異[J]. 湖泊科學,2011,23(2):264-270.
[10]洪云漢,周暾. 鮠科九種魚的核型研究[J]. 動物學研究,1984,5(增刊2):21-28,85-86.
[11]薛淑群,尹洪濱. 黃顙染色體組型的初步分析[J]. 水產學雜志,2006,19(1):11-13.
[12]王祖熊,張錦霞,靳光琴. 魚類雜交不親和性的研究[J]. 水生生物學報,1986,10(2):171-179.
[13]夏仕玲,余來寧,吳作文. 魚類卵細胞質對胚胎發育速度的調控[J]. 淡水漁業,1990,18(3):17-18.郭霄峰,向建國,李煙亮,等. 湘江野生鳡消化道早期發育及食性轉化[J]. 江蘇農業科學,2016,44(5):316-318.
