畢赤酵母X33/pGAPZαA-MLcc1產(chǎn)漆酶的高密度發(fā)酵
2016-09-07 08:37:04黃亮劉逸寒劉曉光李玉路福平工業(yè)發(fā)酵微生物教育部重點(diǎn)實(shí)驗(yàn)室天津市工業(yè)微生物重點(diǎn)實(shí)驗(yàn)室國家工業(yè)酶工程實(shí)驗(yàn)室天津科技大學(xué)生物工程學(xué)院天津300457天津農(nóng)學(xué)院農(nóng)學(xué)與資源環(huán)境學(xué)院天津300384
食品與生物技術(shù)學(xué)報 2016年3期
黃亮,劉逸寒,劉曉光,李玉,路福平*(.工業(yè)發(fā)酵微生物教育部重點(diǎn)實(shí)驗(yàn)室,天津市工業(yè)微生物重點(diǎn)實(shí)驗(yàn)室,國家工業(yè)酶工程實(shí)驗(yàn)室,天津科技大學(xué)生物工程學(xué)院,天津300457;2.天津農(nóng)學(xué)院農(nóng)學(xué)與資源環(huán)境學(xué)院,天津300384)
畢赤酵母X33/pGAPZαA-MLcc1產(chǎn)漆酶的高密度發(fā)酵
黃亮1,2,劉逸寒1,劉曉光1,李玉1,路福平*1
(1.工業(yè)發(fā)酵微生物教育部重點(diǎn)實(shí)驗(yàn)室,天津市工業(yè)微生物重點(diǎn)實(shí)驗(yàn)室,國家工業(yè)酶工程實(shí)驗(yàn)室,天津科技大學(xué)生物工程學(xué)院,天津300457;2.天津農(nóng)學(xué)院農(nóng)學(xué)與資源環(huán)境學(xué)院,天津300384)
在5 L的發(fā)酵罐中進(jìn)行了重組畢赤酵母X33/pGAPZα A-MLcc1表達(dá)重組漆酶的發(fā)酵試驗(yàn),漆酶活性的變化趨勢與菌體干質(zhì)量變化趨勢相一致,當(dāng)菌體增長進(jìn)入穩(wěn)定期46 h左右時,產(chǎn)酶達(dá)至高峰47.2 U/L。進(jìn)一步研究了工程菌在5 L容積發(fā)酵罐中的高密度發(fā)酵工藝,并獲得了高水平表達(dá)漆酶的優(yōu)化策略:在補(bǔ)料階段采用溶氧反饋控制方式,并且保持溶氧水平在15~30 g/dL,發(fā)酵過程中通過在線補(bǔ)加氨水和鹽酸來調(diào)整pH恒定在5。通過高密度發(fā)酵可獲得最高酶活力達(dá)216.3 U/L,是分批發(fā)酵獲得最高酶活力的4.6倍。
畢赤酵母;漆酶;高密度發(fā)酵
漆酶具有特異的催化性能和寬泛的底物范圍,從而在國民經(jīng)濟(jì)的多個領(lǐng)域如紡織業(yè)廢水處理、紙張和紙漿的低污染生產(chǎn)、芳香族化合物轉(zhuǎn)化,生物傳感器、生物燃料電池的制造,以及飲料加工、食藥用菌生產(chǎn)、食品保護(hù)等均有廣闊的應(yīng)用前景,但漆酶的低產(chǎn)量、高溫易失活等又使其應(yīng)用不能深入推廣[1]。
白腐菌是真菌中漆酶主要的生產(chǎn)者,但培養(yǎng)過程周期較長,容易被污染而不利于工業(yè)化生產(chǎn),解決酶源問題的一個重要方法就是通過漆酶的異源表達(dá)實(shí)現(xiàn)漆酶生產(chǎn)低成本的目標(biāo)[2-4]。作者在前期的研究工作中,設(shè)計定點(diǎn)突變位點(diǎn)獲得了突變基因MLcc1,構(gòu)建了重組表達(dá)載體pGAPZα A-MLcc1,并在畢赤酵母中成功實(shí)現(xiàn)了表達(dá)。在前期搖瓶培養(yǎng)研究中,考察了培養(yǎng)基組成、溫度及pH值對重組漆酶表達(dá)量的影響。然而即使分批發(fā)酵處于較優(yōu)的培養(yǎng)基組成及理化條件,最終獲得的細(xì)胞量和重組漆酶的產(chǎn)量依然較低。在此基礎(chǔ)上,本文作者設(shè)計了不同的調(diào)控策略,通過高密度發(fā)酵以期獲得重組酵母發(fā)酵產(chǎn)酶的最佳條件。
1 材料與方法
1.1材料
1.1.1菌株畢赤酵母工程菌TJL22(X33/pGAPZα
A-MLcc1),由作者所在實(shí)驗(yàn)室構(gòu)建。
1.1.2培養(yǎng)基
1)YEPD培養(yǎng)基:蛋白胨20 g/L,葡萄糖20 g/ L,酵母提取物10 g/L;固體培養(yǎng)基加20 g瓊脂粉,用蒸餾水配制。
2)YPG培養(yǎng)基:酵母膏1 g/L、蛋白胨2 g/L、甘油3 g/L,用蒸餾水配制。
3)BMGY培養(yǎng)基:YNB 13.4 g/L,蛋白胨20 g/ L,酵母提取物10 g/L;甘油10 mL/L,100 mmol/L磷酸鉀緩沖液(pH 6),生物素4×10-4,用蒸餾水配制。
4)BSM培養(yǎng)基:體積分?jǐn)?shù)85%H3PO426.7 mL/ L,CaSO40.93 g/L,K2SO418.2 g/L,MgSO4·7H2O 14.9 g/L,KOH 4.13 g/L,甘油40.0 g/L。
5)補(bǔ)料用培養(yǎng)基:葡萄糖30 g/dL,PTM43 mL/ L,硫酸銅0.1 g/dL。
1.1.3試劑及溶液
1)pH 6.0的磷酸鉀緩沖液:1 mol/L K2HPO4132 mL,1 mol/L KH2PO4868 mL,pH 6.0,濕熱滅菌,4℃保存。
2)PTM1微量元素液:CuSO4·5H2O 6.0 g/dL,NaI 0.08 g/dL,MnSO4·H2O 3.0 g/dL,NaMoO4·2H2O 0.2 g/dL,H3BO30.02 g/dL,CoCl20.5 g/dL,ZnCl220.0 g/dL,F(xiàn)eSO4·7H2O 65.0 g/dL,生物素0.2 g/dL,H2SO45.0 mL/L,用0.22 μm的濾膜過濾除菌,室溫保存。
3)PTM4微量元素液:CuSO4·5H2O 2.0 g/dL,NaI 0.08 g/dL,MnSO4·H2O 3.0 g/dL,NaMoO4·2H2O 0.2 g/dL,H3BO30.02 g/dL,CaSO4·2H2O 0.5 g/dL,CoCl20.5 g/dL,ZnCl27.0 g/dL,F(xiàn)eSO4·7H2O 22.0 g/ dL,生物素0.2 g/dL,H2SO41.0 mL/L,用0.22 um的濾膜過濾除菌,室溫保存。
1.1.4發(fā)酵設(shè)備5 L的離位滅菌發(fā)酵罐(型號BIOTECH-5BG),上海保興生物設(shè)備工程有限公司制品。發(fā)酵液溶氧在線檢測,溫度和pH可自動控制。
1.2方法
1.2.1培養(yǎng)方法
1)種子培養(yǎng):從新鮮YEPD平板上挑取單菌落接種到50 mL YPG培養(yǎng)基中,30℃200 r/min振蕩培養(yǎng)過夜,250 mL的搖瓶裝液量50 mL。
2)發(fā)酵罐培養(yǎng):裝料前校準(zhǔn)發(fā)酵罐的pH電極及溶氧電極。加入3 L含0.4 mmol/L銅離子的BMGY培養(yǎng)基,于121℃高壓滅菌30 min,校正溶氧(DO)至0 g/dL。待發(fā)酵罐內(nèi)培養(yǎng)液冷卻到30℃時,用質(zhì)量分?jǐn)?shù)25%氨水(過濾除菌)調(diào)節(jié)pH值。添加PTM1微量元素液12 mL;接入10 g/dL的搖瓶菌種后,通過自控系統(tǒng)調(diào)節(jié)溫度在設(shè)定的范圍內(nèi);攪拌轉(zhuǎn)速400~800 r/min,通氣量控制在4~12 L/(L· min),罐壓0.2~0.3 MPa,校正DO至100 g/dL。由于畢赤酵母在發(fā)酵過程中會產(chǎn)生有機(jī)酸而導(dǎo)致pH值逐漸降低,所以整個發(fā)酵培養(yǎng)pH通過質(zhì)量分?jǐn)?shù)20%氨水和1 mol/L鹽酸進(jìn)行調(diào)節(jié);培養(yǎng)到一定時間后通過添加補(bǔ)料培養(yǎng)基來實(shí)現(xiàn)高密度培養(yǎng)。
3)高密度培養(yǎng):初始培養(yǎng)基配方為添加含0.4 mmol/L銅離子及3 mL/L PTM4微量元素液的改進(jìn)BSM培養(yǎng)基(補(bǔ)充磷酸鉀緩沖液1 g/dL以控制pH,補(bǔ)充蛋白胨3 g/dL以控制蛋白酶對目的蛋白質(zhì)的降解),5 L的發(fā)酵罐裝液1 L。第一階段是菌體生長階段,實(shí)施溫度和pH控制,通過調(diào)整攪拌轉(zhuǎn)速和通氣量控制DO在20 g/dL以上,至溶氧快速反彈表示第一階段結(jié)束。第二階段是補(bǔ)料培養(yǎng)階段,實(shí)施溫度和pH控制,通過不同流加方式并調(diào)整攪拌轉(zhuǎn)速和通氣量控制菌體的生長。
1.2.2分析方法
1)菌體干質(zhì)量:取4 mL發(fā)酵液置于空管質(zhì)量為W0的5 mL的離心管中,8 000 r/min離心10 min,取上清液用于蛋白質(zhì)分析及酶活力測定,沉淀于65℃烘箱烘干至恒質(zhì)量為W1,計算公式
W=250×(W1-W0)。
2)漆酶酶活力的測定(ABTS法[5]):3 mL反應(yīng)混合液中含0.5 mmol/L的ABTS 0.2 mL,pH 4.6的0.1 mol/L醋酸鈉緩沖液2.7 mL和0.1 mL適當(dāng)稀釋的酶液,30℃反應(yīng)3 min,于420 nm處測定吸光度。(ABTS在420 nm處的摩爾消光系數(shù)ε420=36 000 L/ (mol·cm)。在測定中以含滅活酶液的反應(yīng)混合液為對照,酶活力單位定義為:1 min內(nèi)催化氧化1 μmol底物的酶量為1個酶活力單位。
3)還原糖濃度測定:3,5-二硝基水楊酸法。
2 結(jié)果與討論
2.1工程菌種子生長曲線的測定
待種子液培養(yǎng)至一定渾濁度后,每隔一定時間取樣,測定種子液的干質(zhì)量和pH值。繪制細(xì)胞干質(zhì)量—時間、pH—時間曲線,見圖1。由圖可知,培養(yǎng)至48 h時,工程菌對數(shù)生長期結(jié)束,這與原始出發(fā)菌株生長曲線相一致(數(shù)據(jù)略)。在整個培養(yǎng)過程中pH值先下降,到對數(shù)期后保持在3~4之間。
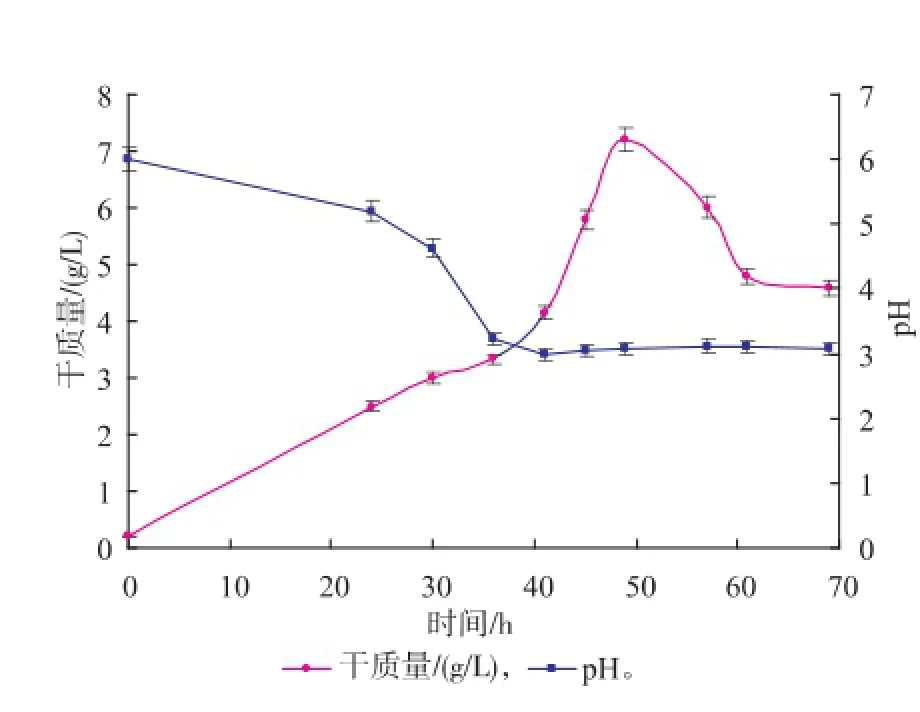
圖1 X33/pGAPZα A-MLcc1的生長曲線及pH值曲線Fig.1 Growth and pH curves of X33/pGAPZα A-MLcc1
2.2工程菌的5 L容積發(fā)酵罐培養(yǎng)試驗(yàn)
在發(fā)酵罐中進(jìn)行畢赤酵母的高密度發(fā)酵,外源蛋白質(zhì)表達(dá)水平可比普通搖瓶高出數(shù)10倍,最重要的原因是搖瓶發(fā)酵的通氣不理想,氧氣供應(yīng)不足,影響了畢赤酵母菌體的高密度生長,造成表達(dá)量偏低。為考察獲得的重組菌株應(yīng)用于工業(yè)化發(fā)酵生產(chǎn)漆酶的潛力,試驗(yàn)在搖瓶發(fā)酵基礎(chǔ)上進(jìn)行5 L的發(fā)酵罐培養(yǎng):以BMGY改進(jìn)培養(yǎng)基(甘油1 g/dL,蛋白胨1 g/dL)為初始發(fā)酵培養(yǎng)基,同時添加銅離子至終濃度0.5 mmol/L;培養(yǎng)溫度為25℃,發(fā)酵罐轉(zhuǎn)速為400 r/min,通氣量控制在4 L/min。以體積分?jǐn)?shù)10%的接種量進(jìn)行發(fā)酵罐試驗(yàn),發(fā)酵開始后每隔4~8 h取樣一次,離心發(fā)酵液進(jìn)行菌體生物量和漆酶活力的監(jiān)測,結(jié)果如圖2所示。

圖2 X33/pGAPZα A-MLcc1的產(chǎn)酶曲線及干質(zhì)量變化曲線Fig.2 Time courses of dry cell weight and enzyme activity in batch culture
發(fā)酵16 h之前菌體處于生長期,16~36 h菌體進(jìn)入分裂生殖期,36 h左右重組菌開始進(jìn)入生長及產(chǎn)酶的穩(wěn)定期,這一時期大概10 h左右。比較菌體干質(zhì)量的變化曲線與漆酶活力的監(jiān)測曲線可以得出,重組菌產(chǎn)生的胞外漆酶活力與菌體干質(zhì)量的增長在對數(shù)期均呈現(xiàn)有較大幅度的提高,當(dāng)菌體進(jìn)一步生長進(jìn)入穩(wěn)定期,發(fā)酵時間46 h時,漆酶產(chǎn)量達(dá)至峰值為47.2 U/L。隨著發(fā)酵的延長,菌體量仍有所增長,但漆酶活力已開始下降。
2.3工程菌的高密度發(fā)酵
通過搖床至發(fā)酵罐的放大培養(yǎng),漆酶活力得到了一定的提高,但遠(yuǎn)達(dá)不到工業(yè)化生產(chǎn)的目的,研究中進(jìn)一步對工程菌實(shí)施高密度培養(yǎng)策略,期望能為漆酶的規(guī)模化生產(chǎn)提供一定的依據(jù)。發(fā)酵初始培養(yǎng)基以BSM培養(yǎng)基為基礎(chǔ)進(jìn)行適當(dāng)?shù)母倪M(jìn)(添加1 g/dL蛋白胨及一定pH值的磷酸緩沖液),并通過在發(fā)酵過程中不同pH、攪拌轉(zhuǎn)速及補(bǔ)料方式的考察,確定較優(yōu)的高密度發(fā)酵條件。
2.3.1補(bǔ)料方式對重組工程菌發(fā)酵產(chǎn)漆酶的影響設(shè)計了恒速飽和流加方式、葡萄糖反饋控制流加方式及溶氧反饋流加方式,考察不同流加策略對重組工程菌發(fā)酵產(chǎn)漆酶的影響,具體實(shí)施方式如下:
1)恒速飽和流加方式,在補(bǔ)料階段以100 mL/h的體積流量添加補(bǔ)料培養(yǎng)基,連續(xù)補(bǔ)料2 L;
2)葡萄糖反饋控制流加方式,至甘油耗盡時補(bǔ)料維持殘?zhí)琴|(zhì)量濃度為3~6 g/L;
3)溶氧反饋流加方式,在補(bǔ)料階段根據(jù)溶氧的變化進(jìn)行補(bǔ)料培養(yǎng)基的添加,通過變速補(bǔ)料保持DO一直處于20~60 g/dL。發(fā)酵過程中漆酶的酶活力監(jiān)測曲線見圖3—5。
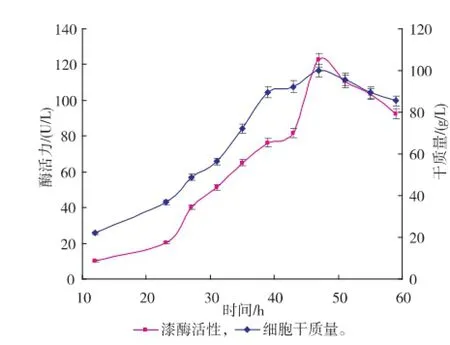
圖3 恒速飽和流加方式下的X33/pGAPZα A-MLcc1的產(chǎn)酶曲線及干質(zhì)量變化曲線Fig.3 Time courses of dry cell weight and laccase activity in fed-batch culture using constant feeding
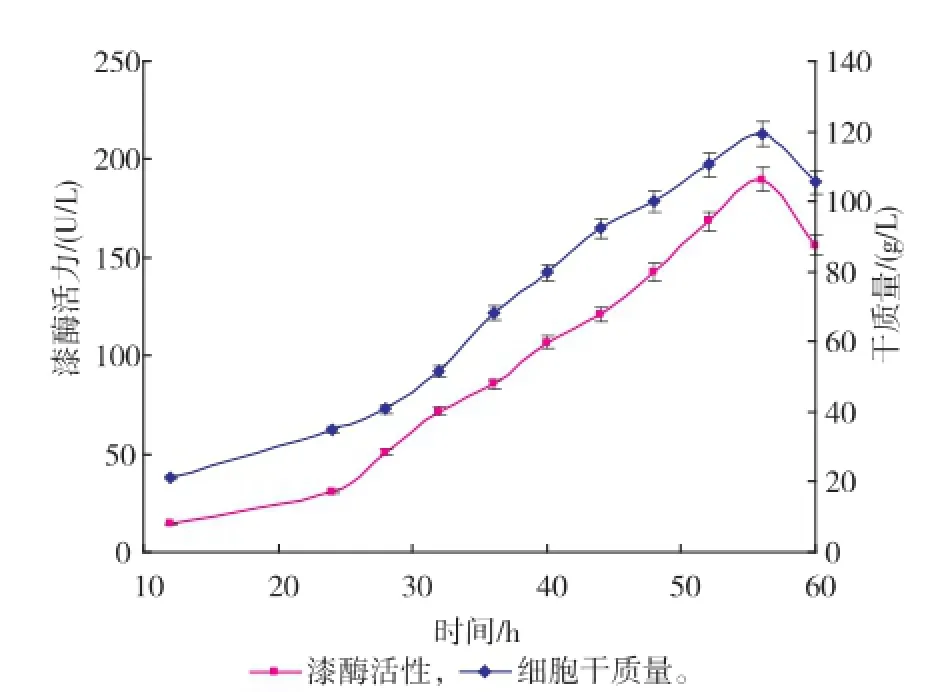
圖4 葡萄糖反饋控制流加方式下的X33/pGAPZα AMLcc1的產(chǎn)酶曲線及干質(zhì)量變化曲線Fig.4 Time courses of dry cell weight and laccase activity in fed-batch culture using glucose concentration feed-back control strategy

圖5 溶氧反饋流加方式下的X33/pGAPZα A-MLcc1的產(chǎn)酶曲線及干質(zhì)量變化曲線Fig.5 Time courses of dry cell weight and laccase activity in fed-batch culture using DO feed-back control strategy
由圖可以看出,漆酶酶活力的增加和細(xì)胞干質(zhì)量的提高呈正相關(guān),也表明了本研究中的漆酶基因是組成型表達(dá),不需要更換甲醇再進(jìn)行誘導(dǎo),故發(fā)酵產(chǎn)酶研究可通過菌體的密度增加實(shí)現(xiàn)。但補(bǔ)料階段方式的不同,獲得的酶活力高低及菌體干質(zhì)量的增長也有差別,其中溶氧反饋控制模式獲得的酶活力最高。恒速飽和流加方式可以最大限度地滿足重組菌生長的營養(yǎng)需求,以致于菌體在補(bǔ)料初期快速增長,代謝的加快也使得菌體的衰老期提前,導(dǎo)致產(chǎn)酶后勁不足。葡萄糖反饋控制模式能很好地限制菌體在幾乎不受有害代謝物影響的情況下增長,使得菌體在補(bǔ)料階段呈較好的指數(shù)型生長,但后期營養(yǎng)物的比例失調(diào)及有害代謝物的累積,菌體最終走向衰亡;菌體限制性的生長也使得組成型表達(dá)的漆酶活力不能夠快速提升,最終達(dá)至最高點(diǎn)189.8 U/L后即隨著干質(zhì)量一起下降。溶氧反饋模式通過菌體的實(shí)時生長情況進(jìn)行補(bǔ)料,既保證了菌體養(yǎng)分的供應(yīng),又能在一定程度上促進(jìn)漆酶的組成型表達(dá);漆酶的酶活力雖然在補(bǔ)料過程中被稀釋而表現(xiàn)出輕微下降情況,但在總體培養(yǎng)環(huán)境上優(yōu)于前兩者控制模式,最終可收獲194.1 U/L的酶活力。在此模式下,每次降低溶氧需要補(bǔ)料200~500 mL,使得每次補(bǔ)料后漆酶被稀釋而酶活呈現(xiàn)短時下降的趨勢;漆酶在后期(尤其是在酶活力達(dá)至最高點(diǎn)后)存在降勢,此時補(bǔ)料量持續(xù)增加仍不能維持設(shè)計的溶氧水平,菌體的高密度培養(yǎng)狀態(tài)被打破,通過檢測得出此時蛋白酶活力較高(數(shù)據(jù)未列出),菌體進(jìn)入衰亡期。
2.3.2pH值對重組工程菌發(fā)酵產(chǎn)漆酶的影響pH值可通過抑制菌體中某些酶的活性,影響微生物細(xì)胞膜的通透性及改變培養(yǎng)基中某些組分和中間代謝產(chǎn)物的離解等,對菌體生長和目的蛋白質(zhì)分泌表達(dá)的影響較大。畢赤酵母在pH 3~8的范圍內(nèi)均可生長,當(dāng)pH低于2.2時菌體會停止生長,當(dāng)pH值超過8,菌體生長和表達(dá)外源蛋白質(zhì)也會很不理想。盡管畢赤酵母生長的pH值范圍較寬,但是具體到不同的重組工程菌,則各有不同的最適生長pH值。另外,對于同一菌株,其生長和表達(dá)的最適pH值往往不同[6]。試驗(yàn)選取pH值為4、5、6、7,發(fā)酵過程中pH的酸堿性調(diào)控是通過在線補(bǔ)加氨水和鹽酸來調(diào)整,以溶氧反饋控制補(bǔ)料方式實(shí)施高密度發(fā)酵,考察工程菌分泌表達(dá)的最適pH值。不同pH值下的重組菌高密度發(fā)酵獲得的最高酶活力見圖6。
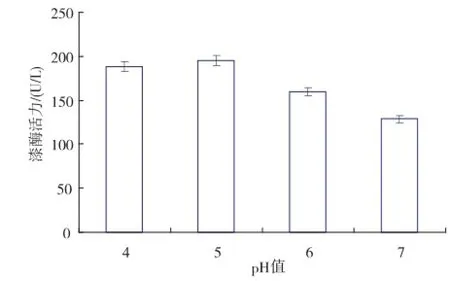
圖6 pH值對重組菌發(fā)酵產(chǎn)漆酶的影響Fig.6 Effects of pH values on the yields of laccase from recombinant stain fermentation
在基因工程菌分泌表達(dá)外源蛋白質(zhì)過程中,重組漆酶對pH值存在一定的敏感性,容易受到蛋白水解酶的降解,選擇合適的pH值可以使菌體高效表達(dá)目的蛋白質(zhì)又能抑制蛋白酶的水解。從圖6得知,通過高密度發(fā)酵,酶活力水平較之分批發(fā)酵有了很大的提高,均達(dá)至100 U/L以上;在恒定pH為5的條件下,可通過高密度發(fā)酵獲得最高酶活力約200 U/L,是搖瓶發(fā)酵獲得最高酶活力的5倍(數(shù)據(jù)略)。
2.3.3不同溶氧控制水平對重組工程菌發(fā)酵產(chǎn)漆酶的影響發(fā)酵基質(zhì)中DO對于好氧微生物的生長以及發(fā)酵產(chǎn)物量的提高非常重要。提高發(fā)酵罐內(nèi)溶氧水平的方式主要有:增加通氣量、加大罐壓、增加攪拌轉(zhuǎn)速,甚至直接通入純氧。畢赤酵母是好氧微生物,它的生長代謝過程需要氧的參與,但溶解氧的質(zhì)量濃度過高或過低都會影響畢赤酵母的代謝,使后期的生長變得極為緩慢,從而影響到蛋白質(zhì)的表達(dá)。試驗(yàn)中通過補(bǔ)料階段不同溶氧控制水平,研究溶氧對重組菌產(chǎn)漆酶的影響,期間獲得的最大細(xì)胞干質(zhì)量及最高漆酶酶活力結(jié)果見圖7。

圖7 DO對重組工程菌發(fā)酵產(chǎn)漆酶的影響Fig.7 Effects of various DO values on the yields of laccase from recombinant stain fermentation
畢赤酵母在生長初期就需要保證足量的氧供應(yīng),一般在菌體前期生長階段,發(fā)酵罐內(nèi)溶氧水平保持在較高水平(30 g/dL以上),在發(fā)酵過程中溶氧逐漸下降,當(dāng)?shù)陀?0 g/dL時即提高轉(zhuǎn)速并加大通氣量,使其不低于30 g/dL;當(dāng)出現(xiàn)溶氧突然陡直上升時,開始進(jìn)行補(bǔ)料,控制DO低于45 g/dL,因?yàn)榧?xì)胞在高溶氧條件下容易發(fā)生氧中毒,不利于菌體的生長,導(dǎo)致外源蛋白質(zhì)表達(dá)量不高。由圖7可以看出,溶氧水平控制在15~30 g/dL之間可以獲得最高的酶活力216.3 U/L。溶氧過低(0~15 g/dL)時菌體受到氧的限制呼吸作用減弱,故而影響到了漆酶的表達(dá)。溶氧在30~45 g/dL時可以獲得最高的菌體干質(zhì)量,但漆酶的表達(dá)水平并沒有達(dá)到最高,可見漆酶組成型表達(dá)過程中還有諸多影響因素,導(dǎo)致漆酶的表達(dá)不能達(dá)到和菌體增加呈完全正相關(guān);溶氧控制在45~60 g/dL時,細(xì)胞干質(zhì)量及漆酶酶活力水平均不是太高,可見氧毒害作用已經(jīng)開始影響到菌體的生長了。本試驗(yàn)結(jié)果與張建勇等[7]的研究結(jié)果相一致。可見,保持溶氧在有利于外源蛋白質(zhì)高效表達(dá)的范圍,對于高密度發(fā)酵來說非常重要。
3 結(jié)語
pGAPZ是一類新型的酵母組成型表達(dá)質(zhì)粒,其啟動子可與表達(dá)載體一起整合到基因組中,可有效地避免其融合的外源基因的丟失,具有帶動外源基因高效、連續(xù)表達(dá)的潛力。目前該表達(dá)系統(tǒng)已經(jīng)應(yīng)用于多種酵母菌株,獲得了異源蛋白質(zhì)表達(dá)并逐步深入和優(yōu)化,高密度發(fā)酵其突出的簡便可行性及多種優(yōu)異特性正受到越來越多研究者的關(guān)注。由于pGAP在工程菌發(fā)酵產(chǎn)外源蛋白質(zhì)時僅需通過簡單的發(fā)酵工藝而不需要AOX啟動子引導(dǎo)的外源蛋白質(zhì)表達(dá)誘導(dǎo)階段,避免了誘導(dǎo)物甲醇等可能引起的“二次污染”及不安全因素[8],因此是后期工程菌在工業(yè)化規(guī)模表達(dá)外源蛋白質(zhì)的首選。
對基因工程菌X33/pGAPZα A-MLcc1進(jìn)行了5 L容積發(fā)酵罐的高密度培養(yǎng),通過在補(bǔ)料階段采用溶氧反饋控制方式獲得了漆酶的胞外高表達(dá)。通過高密度發(fā)酵可獲得漆酶的最高酶活力達(dá)216.3 U/L,是分批發(fā)酵獲得的最高酶活力的4.6倍,采用組成型表達(dá)模式可在較短時間內(nèi)獲得漆酶表達(dá)峰值,簡化了工藝,對漆酶的工業(yè)化應(yīng)用具有參考意義。
[1]高玉千,張利明,張世敏,等.真菌漆酶高級結(jié)構(gòu)研究進(jìn)展[J].食品與生物技術(shù)學(xué)報,2011,30(2):166-171. GAO Yuqian,ZHANG Liming,ZHANG Shimin,et al.Research progress of fungal laccase structure[J].Journal of Food Science and Biotechnology,2011,30(2):166-171.(in Chinese)
[2]Giardina P,Annio R,Martirani L,et al.Cloning and sequencing of a laccase gene from the lignin-degrading basidiomyccte Pleurotus ostreatus[J].Appl Environ Microbiol,1995,61(6):2408-2413.
[3]郭梅,蒲軍,杜連祥,等.雜色云芝漆酶基因(Lccl)的克隆及在甲醇畢赤酵母中的表達(dá)[J].菌物學(xué)報,2005,24(2):221-226. GUO Mei,PU Jun,DU Lianxiang,et al.Coloning of cDNA encoding laccase(Lccl)from Coriolus versicolor and expression of the gene in Pichia methanolica[J].Mycosystema,2005,24(2):221-226.(in Chinese)
[4]馬文寅,蔡宇杰,廖祥儒,等.一種真菌漆酶的克隆與序列分析[J].食品與生物技術(shù)學(xué)報,2012,31(1):40-46. MA Wenyin,CAI Yujie,LIAO Xiangru,et al.Cloning and sequencing analysis of a laccase gene from fungus[J].Journal of Food Science and Biotechnology,2012,31(1):40-46.(in Chinese)
[5]Liu X,Gillespie M,Ozel A D,et al.Electrochemical properties and temperature dependence of a recombinant laccase from Thermus thermophilus[J].Anal Bioanal Chem,2011,399(1):361-366.
[6]王學(xué)強(qiáng),程相朝,劉一塵,等.重組雞IL-18與新城疫HN基因融合蛋白在畢赤酵母中表達(dá)條件的優(yōu)化[J].河南科技大學(xué)學(xué)報:自然科學(xué)版,2010,31(2):73-77. JIAN Yun,CUI Shujun,GU Likun,et al.Optimation of condition for expression of recombinant ChMIL18-HN in Pichia pastoris [J].Journal of Henan University of Science&Technology:Natural Science,2010,31(2):73-77.(in Chinese)
[7]張建勇,王曉港,王水蓮,等.溶氧對重組畢赤酵母高密度發(fā)酵生產(chǎn)腺苷蛋氨酸的影響[J].齊魯藥事,2007,26(7):428-430. ZHANG Jianyong,WANG Xiaogang,WANG Shuilian,et al.Effects of the dissolved oxygen on producing S-adenosyl-L-methionine by High-density fermentation of recombinant Pichia pastori[J].Qilu Pharmaceutical Affairs,2007,26(7):428-430.(in Chinese)
[8]婁瑞娟,羅利龍,張霞,等.巴斯德畢赤酵母表達(dá)系統(tǒng)的研究進(jìn)展和前景展望[J].生物學(xué)雜志,2010,27(5):73-76. LOU Ruijuan,LUO Lilong,ZHANG Xia,et al.Research progress and prospects on Pichia pastoris[J].Journal of Biology,2010,27(5):73-76.(in Chinese)
Study of High Cell-Density Fermentation of Laccase Produced by Pichia pastoris X33/pGAPZαA-MLcc1
HUANG Liang1,2,LIU Yihan1,LIU Xiaoguang1,LI Yu1,LU Fuping*1
(1.Key Laboratory of Industrial Fermentation Microbiology,Ministry of Education,Tianjin Key Laboratory of Industrial Microbiology,National Engineering Laboratory for Industrial Enzymes,The College of Biotechnology,Tianjin University of Science&Technology,Tianjin 300457,China;2.College of Agronomy and Resources Environmental,Tianjin Agricultural University,Tianjin 300384,China)
The expression of laccase gene by Pichia pastoris X33/pGAPZα A-MLcc1 was investigated in a 5 L fermenter.The change trend of laccase activity was consistent with the change trend dry cell weight.The extracellular laccase activity reached the highest value of 47.2 U/L when the yeast X33 entered the stationary phage at 46 h.High-density fermentation in a 5 L fermenter was further investigated and the optimized method for high-level expression was obtained as follows:by adopting dissolved oxygen(DO)feed-back feeding strategy and maintaining DO at 15%to 30%at feeding stage,as well as adding ammonia solution and hydrochloride to maintain pH value at 5 in the fermentation stage,the laccase activity reached 216.3 U/L,which was 4.6 times higher than that of fed batch fermentation.
Picha pastoris;laccase;high cell-density fermentation
Q 815
A
1673—1689(2016)03—0324—06
2014-12-01
天津市科技支撐計劃項(xiàng)目(14ZCZDSY00014);天津市科技特派員項(xiàng)目(15JCTPJC56500)。
黃亮(1983-),男,江蘇沐陽人,工學(xué)碩士,副教授,主要從事微生物發(fā)酵及食用菌相關(guān)研究。E-mail:huangliang@tjau.edu.cn
路福平(1967—),男,山西運(yùn)城人,工學(xué)博士,教授,主要從事酶與應(yīng)用微生物研究。E-mail:lfp@tust.tjau.edu.cn
