不同生長時(shí)期羅漢果果實(shí)轉(zhuǎn)錄因子的轉(zhuǎn)錄組分析及酵母單雜交文庫的構(gòu)建△
2016-11-01 20:55:45張凱倫羅祖良郭玉華石宏武馬小軍
中國現(xiàn)代中藥 2016年8期
關(guān)鍵詞:差異
張凱倫,羅祖良,郭玉華,石宏武,馬小軍,2*
(1.中國醫(yī)學(xué)科學(xué)院 北京協(xié)和醫(yī)學(xué)院 藥用植物研究所,北京 100193;2.中國醫(yī)學(xué)科學(xué)院 藥用植物研究所 云南分所,云南 景洪 666100)
·基礎(chǔ)研究·
不同生長時(shí)期羅漢果果實(shí)轉(zhuǎn)錄因子的轉(zhuǎn)錄組分析及酵母單雜交文庫的構(gòu)建△
張凱倫1,羅祖良1,郭玉華1,石宏武1,馬小軍1,2*
(1.中國醫(yī)學(xué)科學(xué)院 北京協(xié)和醫(yī)學(xué)院 藥用植物研究所,北京 100193;2.中國醫(yī)學(xué)科學(xué)院 藥用植物研究所 云南分所,云南 景洪 666100)
目的研究羅漢果果實(shí)中的轉(zhuǎn)錄因子(transcription factors,TFs)。方法在授粉后3、50、70 d(days after flowering,DAF)的羅漢果果實(shí)轉(zhuǎn)錄組中查詢注釋為轉(zhuǎn)錄因子的所有非重復(fù)Unigene,并在表達(dá)譜中查找其表達(dá)情況;應(yīng)用Matchmaker Yeast One-Hybrid Library Construction系統(tǒng)構(gòu)建酵母單雜交cDNA文庫。結(jié)果羅漢果表達(dá)譜數(shù)據(jù)中共有38個(gè)家族的119個(gè)轉(zhuǎn)錄因子Unigene具有表達(dá)差異,其中以70 DAF/3 DAF和50 DAF/3 DAF組中具有差異表達(dá)的轉(zhuǎn)錄因子數(shù)目較多,且上調(diào)和下調(diào)的轉(zhuǎn)錄因子Unigene數(shù)目相近。此外,初步篩選到4個(gè)可能參與羅漢果甜苷合成調(diào)控的bHLH(basic helix-loop-helix,bHLH)轉(zhuǎn)錄因子Unigene(bHLH014、bHLH025、bHLH093、bHLH096)及8個(gè)可能參與黃酮類化合物合成調(diào)控的MYB轉(zhuǎn)錄因子Unigene;成功構(gòu)建了酵母單雜交文庫pGADT7-Rec2-Lib,其庫容量為2.3×106 cfu,插入片段的平均長度約為1.5 kbp。結(jié)論轉(zhuǎn)錄組數(shù)據(jù)分析和酵母單雜交實(shí)驗(yàn)有助于篩選羅漢果果實(shí)中具有調(diào)控作用的轉(zhuǎn)錄因子。
羅漢果;轉(zhuǎn)錄因子;轉(zhuǎn)錄組;酵母單雜交文庫
羅漢果是多年生藤本植物葫蘆科(Cucurbitaceae)羅漢果屬(Siraitia)羅漢果Siraitiagrosvenorii的果實(shí),主要分布于中國廣西壯族自治區(qū),其主要化學(xué)成分為葫蘆烷型三萜皂苷羅漢果甜苷。作為藥食同源植物,羅漢果一方面長期作為傳統(tǒng)中藥具有鎮(zhèn)咳平喘、抗氧化、調(diào)節(jié)免疫等療效[1];另一方面由于其成熟果實(shí)提取物的高甜度(比蔗糖還要甜300倍)[2],在作為食品添加劑及糖尿病患者的非糖甜味劑方面具有廣闊的應(yīng)用前景。
轉(zhuǎn)錄因子(transcription factors,TFs)即反式作用分子,在細(xì)胞核內(nèi)能夠特異性識別并結(jié)合基因啟動子上的順式作用元件(cis-acting element),調(diào)控基因的時(shí)空表達(dá)。它們與植物的生長發(fā)育[3]、次生代謝[4]和抗逆反應(yīng)[5]有著密切的關(guān)系。典型的植物轉(zhuǎn)錄因子是由核定位信號區(qū)(nuclear localization signal,NLS)、DNA結(jié)合區(qū)(DNA binding domain,DBD)、轉(zhuǎn)錄調(diào)控區(qū)(transcription regulation domain,TRD)和寡聚化位點(diǎn)(oligomerization site,OS)組成。根據(jù)其中DNA結(jié)合區(qū)氨基酸序列的不同,可分為具有不同結(jié)構(gòu)域的轉(zhuǎn)錄因子家族,如MYB、bHLH、WRKY、bZIP、AP2/EREBP等。常用的分離轉(zhuǎn)錄因子的方法為酵母單雜交法(yeast one-hybrid,Y1H),它是在體外分析DNA(目的基因順式作用元件)與蛋白質(zhì)(轉(zhuǎn)錄因子)相互作用的有效方法,主要通過構(gòu)建誘餌載體和酵母單雜交文庫,對陽性克隆進(jìn)行測序從而篩選出與基因啟動子區(qū)結(jié)合的轉(zhuǎn)錄因子。現(xiàn)已用酵母單雜交技術(shù)克隆到多種植物中的轉(zhuǎn)錄因子,如豌豆[6]、長春花[7]、中國紅豆杉等[8]。
2011年,Tang等[9]應(yīng)用Solexa高通量測序技術(shù)對授粉后3、50、70 d(days after flowering,DAF)的羅漢果果實(shí)進(jìn)行轉(zhuǎn)錄組測序和表達(dá)譜分析,開啟了關(guān)于羅漢果分子生物學(xué)的研究。現(xiàn)已從轉(zhuǎn)錄組數(shù)據(jù)中鑒定了羅漢果甜苷骨架生物合成途徑中的所有酶基因,同時(shí)發(fā)現(xiàn)了多個(gè)家族轉(zhuǎn)錄因子的存在。本研究主要通過分析轉(zhuǎn)錄組和表達(dá)譜數(shù)據(jù)中的轉(zhuǎn)錄因子及建立酵母單雜交cDNA文庫,旨在為羅漢果果實(shí)中轉(zhuǎn)錄因子的篩選及其調(diào)控機(jī)制的研究奠定基礎(chǔ)。
1 材料與儀器
1.1 材料
實(shí)驗(yàn)材料為于30 DAF經(jīng)500 μmol·L-1茉莉酸甲酯(methyl jasmonate,MeJA)誘導(dǎo)后24 h的羅漢果果實(shí)及已提取的果實(shí)cDNA(均保存于-80 ℃),3、50、70 DAF的羅漢果果實(shí)轉(zhuǎn)錄組和表達(dá)譜數(shù)據(jù)[9]。
1.2 試劑
Trizol Reagent(Invitrogen)、熒光定量試劑盒SYBR Premix Ex TaqTMⅡ(TliRNaseH Plus)(TaKaRa,Dalian,China)、酵母單雜交系統(tǒng)MatchmakerTMOne-Hybrid Library Construction & Screening Kit(Catalog No.630304,Clontech)、酵母質(zhì)粒提取試劑盒(D1160-100,Solarbio)。DNA合成由生工生物工程股份有限公司完成;各種酶制劑及大腸桿菌感受態(tài)購自Takara公司(大連)。
1.3 儀器
NanoDrop 2000 spectrophotometer (Thermo Scientific,USA)、AB 7500 Fast Real-Time PCR System (Applied Biosystem,USA)、電轉(zhuǎn)化儀(Bio-Rad)、PCR儀(Bio-Rad)。
2 方法
2.1 轉(zhuǎn)錄組數(shù)據(jù)中轉(zhuǎn)錄因子的分析
對羅漢果3、50、70 DAF轉(zhuǎn)錄組數(shù)據(jù)中注釋為轉(zhuǎn)錄因子的Unigene進(jìn)行整理,然后在表達(dá)譜中查找包括50 DAF/3 DAF、70 DAF/3 DAF和70 DAF/50 DAF組中所有表達(dá)有差異的轉(zhuǎn)錄因子Unigene并進(jìn)行分類。
2.2 熒光定量PCR篩選可能參與甜苷合成的bHLH轉(zhuǎn)錄因子
由于羅漢果甜苷V的含量在羅漢果果實(shí)生長發(fā)育的后期才開始激增(50~70 DAF),有表達(dá)差異的轉(zhuǎn)錄因子Unigene中bHLH轉(zhuǎn)錄因子家族數(shù)目最多,因此選擇在70 DAF/3 DAF和50 DAF/3 DAF組中均有表達(dá)差異的bHLH轉(zhuǎn)錄因子Unigene作為研究對象,用經(jīng)驗(yàn)證施加MeJA后甜苷含量和合成酶基因表達(dá)量均增加的羅漢果果實(shí)cDNA作為實(shí)驗(yàn)材料,應(yīng)用熒光定量PCR檢測表達(dá)譜中表達(dá)差異較大(2個(gè)組的基因表達(dá)量log2值均大于2)的6個(gè)候選bHLH轉(zhuǎn)錄因子Unigene在MeJA誘導(dǎo)下的表達(dá)情況,選擇UBQ5為內(nèi)參基因,引物見表1。

表1 用于熒光定量PCR實(shí)驗(yàn)的引物
2.3 總RNA的提取及mRNA的分離
選用前期實(shí)驗(yàn)室已驗(yàn)證經(jīng)MeJA處理后,甜苷含量及合成酶基因表達(dá)量有明顯提高的羅漢果果肉,用改良Trizol法[10]提取樣品中的總RNA。使用Oligotex mRNA Kits(Qiagen)分離純化樣品的mRNA。
2.4 cDNA的合成與純化
將分離得到的樣品mRNA轉(zhuǎn)移到RNase free離心管1中,加入7 μL mRNA、1 μL 3′RTPrimer(30 pmol)和1 μL dNTPs(10 mmol·L-1),65 ℃處理5 min,立即置于冰上。在一個(gè)新的RNase free 離心管2中加入4 μL 5×RT Buffer、2 μL 0.1 mol·L-1DTT、5 μL RT 酶,并置于冰上待用。待管1降至45 ℃后,維持在45 ℃2 min,再加入管2中的混勻液,繼續(xù)在50 ℃條件下孵育1 h合成第一鏈cDNA。
繼續(xù)在反應(yīng)管中加入92 μL DEPC-treated water、30 μL 5×Second Strand Buffer、3 μL 10 mmol·L-1(each) dNTPs、1 μLE.coliDNA Ligase(10 U·μL-1)、4 μLE.coliDNA Polymerase I和1 μLE.coliRNaseH(2 U·μL-1),在16 ℃孵育2 h。然后加入2 μLT4DNA Polymerase,于16 ℃孵育5 min,加入10 μL 0.5 mol·L-1EDTA(pH8.0)及160 μL酚-三氯甲烷-異戊醇(25∶24∶1)溶液,充分混勻30 s,在室溫條件下,于14 000 r·min-1離心5 min,小心將上清液倒入新的離心管中,用乙醇沉淀,最后溶于105 μL DEPC水中。
在新的反應(yīng)管中加入34 μL cDNA、5 μL 10×T4 ligase Buffer、10 μL 5′ Adapter(1 μg·μL-1)和1 μL T4 DNA Ligase(40 U·μL-1,NEB)。混勻后,在16 ℃放置16~24 h。使用低熔點(diǎn)瓊脂糖電泳cDNA,切膠回收1000 bp以上的條帶,乙醇沉淀后溶于14 μL水中,并進(jìn)行電泳檢測。
2.5 cDNA與載體的連接
在1.5 mL離心管中加入14 μL cDNA、2 μL pGADT7(300 ng·μL-1)和4 μL 5×Infusion enzyme mix,混勻后置于50 ℃孵育30 min,然后向重組反應(yīng)體系中加入2 μL的ProteinaseK,在37 ℃孵育15 min,繼續(xù)在75 ℃孵育10 min,補(bǔ)水到100 μL。
然后向反應(yīng)體系中依次加入1 μL Glycogen(20 μg·μL-1)、50 μL 7.5 mol·L-1NH4OAc和375 μL 100% ethanol,混合均勻后置于-80 ℃ 1 h,于4 ℃、16 000 r·min-1離心30 min;小心去上清液,加入150 μL的70%乙醇,于4 ℃、16 000 r·min-1離心3 min;重復(fù)上述步驟1次,去盡上清液,在室溫下將cDNA晾干5~10 min;用10 μL的DEPC水重懸cDNA沉淀,用移液槍吹吸30~40次。瞬時(shí)離心2 s收集cDNA,立即放置于冰上。
2.6 酵母單雜交文庫的構(gòu)建及質(zhì)量鑒定
將2 μL的重組產(chǎn)物轉(zhuǎn)化50 μL大腸桿菌感受態(tài)細(xì)胞,取轉(zhuǎn)化后的細(xì)菌原液10 μL稀釋100倍,從中取出10 μL涂布LB平板(含氨芐抗性),第二天計(jì)數(shù),計(jì)算公式為:總克隆數(shù)=平板上克隆數(shù)×稀釋倍數(shù)×菌液體積。挑取平板上的單克隆,進(jìn)行PCR擴(kuò)增,并用凝膠電泳檢測PCR產(chǎn)物大小。使用Qiagen大抽試劑盒提取質(zhì)粒,保存于-80 ℃冰箱內(nèi)。
3 結(jié)果與分析
3.1 羅漢果轉(zhuǎn)錄組與表達(dá)譜數(shù)據(jù)中轉(zhuǎn)錄因子的分析
3.1.1 轉(zhuǎn)錄組數(shù)據(jù)分析 在羅漢果轉(zhuǎn)錄組數(shù)據(jù)中發(fā)現(xiàn)已注釋家族的轉(zhuǎn)錄因子共468個(gè),其中數(shù)目最多的3個(gè)轉(zhuǎn)錄因子家族為bHLH、WRKY和MYB轉(zhuǎn)錄因子,它們的數(shù)量分別為79、57、50個(gè)。此外,轉(zhuǎn)錄組數(shù)據(jù)中還有其他類型的轉(zhuǎn)錄因子,包括19個(gè)HSF、16個(gè)TCP、9個(gè)HB、6個(gè)MYB-related和5個(gè)bZIP等轉(zhuǎn)錄因子家族。
3.1.2 表達(dá)譜數(shù)據(jù)分析 對3、50、70 DAF的羅漢果果實(shí)表達(dá)譜數(shù)據(jù)進(jìn)行分析,以任意一個(gè)組有差異變化為標(biāo)準(zhǔn),發(fā)現(xiàn)了共38個(gè)家族的119個(gè)轉(zhuǎn)錄因子Unigene(占總轉(zhuǎn)錄因子數(shù)目的25.4%)具有表達(dá)差異,其中數(shù)目最多的3個(gè)轉(zhuǎn)錄因子家族為bHLH、WRKY和MYB轉(zhuǎn)錄因子,它們的數(shù)量分別為22、14、14個(gè)。在70 DAF/3 DAF、50 DAF/3 DAF和70 DAF/50 DAF 3個(gè)組中具有表達(dá)差異的Unigene數(shù)目分別為86、91和40個(gè),且在70 DAF/3 DAF、50 DAF/3 DAF組中上調(diào)和下調(diào)的轉(zhuǎn)錄因子Unigene數(shù)目相近,在70 DAF/50 DAF組中上調(diào)的轉(zhuǎn)錄因子Unigene數(shù)目比下調(diào)的數(shù)目多(見圖1)。對3個(gè)組進(jìn)行比較,發(fā)現(xiàn)在3個(gè)組中均有表達(dá)差異的轉(zhuǎn)錄因子Unigene個(gè)數(shù)為10;對2個(gè)組分別進(jìn)行比較,發(fā)現(xiàn)在70 DAF/3 DAF和50 DAF/3 DAF組中均有表達(dá)差異的轉(zhuǎn)錄因子Unigene共58個(gè),在70 DAF/3 DAF和70 DAF/50 DAF組中均有表達(dá)差異的轉(zhuǎn)錄因子Unigene共18個(gè),在50 DAF/3 DAF和70 DAF/50 DAF組中均有表達(dá)差異的轉(zhuǎn)錄因子Unigene共27個(gè)。
由于羅漢果甜苷V的含量在果實(shí)生長后期50~70 DAF激增,因此為了篩選可能參與甜苷合成的轉(zhuǎn)錄因子,選擇在70 DAF/3 DAF和50 DAF/3 DAF組中均上調(diào)表達(dá)的共16個(gè)家族的33個(gè)轉(zhuǎn)錄因子Unigene進(jìn)行分析,它們的表達(dá)變化范圍為1.03~13.89。其中包括12個(gè)bHLH轉(zhuǎn)錄因子,其他家族還包括TCP、MYB、WRKY、GRAS等,數(shù)量均≤3(見表2)。
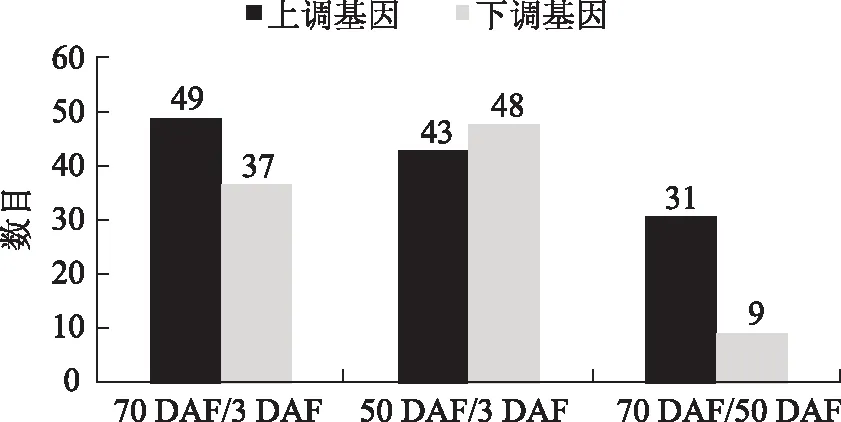
圖1 羅漢果表達(dá)譜中具有表達(dá)差異的轉(zhuǎn)錄因子
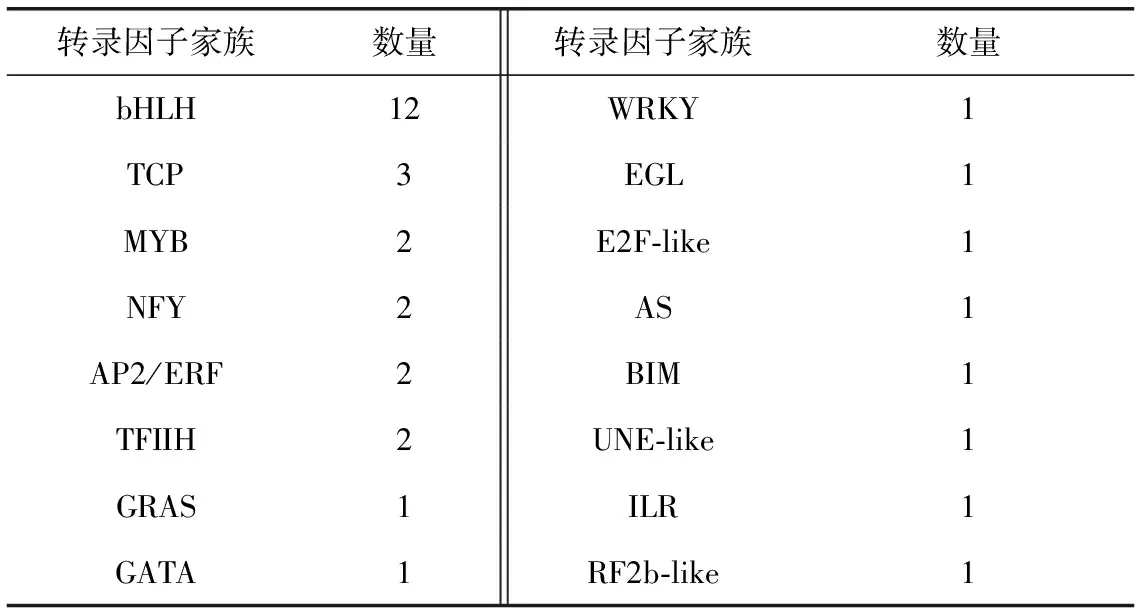
表2 70 DAF/3 DAF和50 DAF/3 DAF組中均上調(diào)表達(dá)的轉(zhuǎn)錄因子Unigene
3.1.3 參與調(diào)控羅漢果甜苷合成途徑的bHLH轉(zhuǎn)錄因子的篩選 選擇在70 DAF/3 DAF和50 DAF/3 DAF組中表達(dá)差異較大的6個(gè)注釋為bHLH轉(zhuǎn)錄因子的Unigene(bHLH014、bHLH025、bHLH061、bHLH071、bHLH093、bHLH096),以經(jīng)MeJA誘導(dǎo)后的羅漢果果實(shí)作為實(shí)驗(yàn)材料,應(yīng)用qRT-PCR篩選與甜苷合成關(guān)鍵酶基因協(xié)同表達(dá)的bHLH轉(zhuǎn)錄因子。結(jié)果顯示除了bHLH061和bHLH071基因外(結(jié)果未顯示),其他4個(gè)bHLH基因bHLH014、bHLH025、bHLH093、bHLH096的表達(dá)量均被不同程度地上調(diào)(見圖2),表明它們可能參與甜苷合成的調(diào)控。其中,bHLH093和bHLH096基因在6 h的表達(dá)量較0 h相比,增加了9~10倍;bHLH025在6 h則增加了1.8倍。bHLH014基因的表達(dá)量在2 h達(dá)到最大值,為0 h的2.5倍。
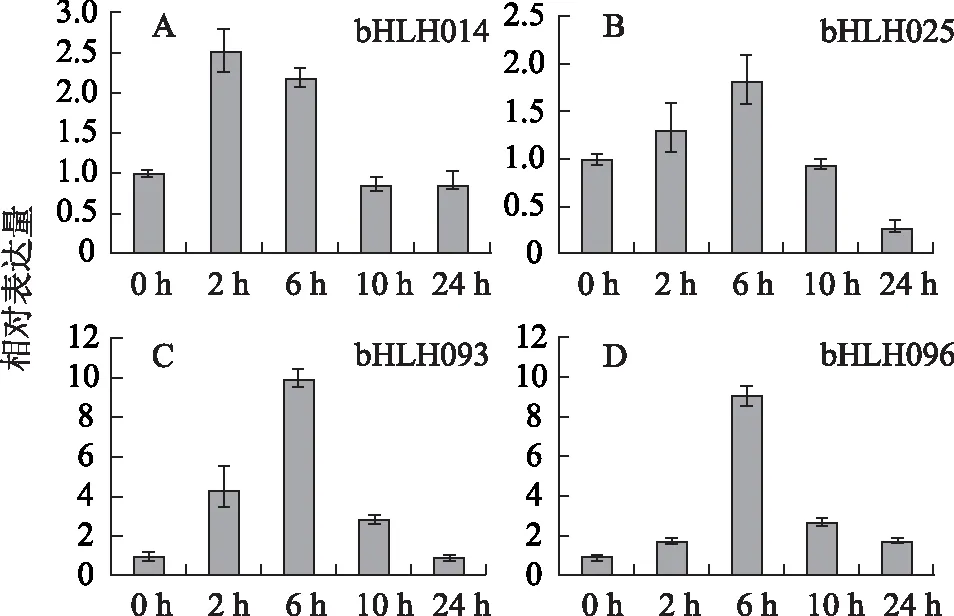
注:A.bHLH014;B.bHLH025;C.bHLH093;D.bHLH096。圖2 4個(gè)bHLH轉(zhuǎn)錄因子的熒光定量PCR測定結(jié)果
3.1.4 參與調(diào)控果實(shí)中黃酮類化合物合成途徑的MYB轉(zhuǎn)錄因子的初步篩選 已報(bào)道羅漢果果實(shí)中含有少量的黃酮類化合物,且總黃酮的含量隨著果實(shí)的生長發(fā)育先逐漸增長,在50 DAF時(shí)達(dá)到含量的峰值,然后下降[11]。MYB類轉(zhuǎn)錄因子是調(diào)控植物中黃酮類化合物生物合成的主要轉(zhuǎn)錄因子家族[12]。因此,在表達(dá)譜50 DAF/3 DAF組數(shù)據(jù)中找到了8個(gè)MYB類轉(zhuǎn)錄因子Unigene,可能參與羅漢果果實(shí)中黃酮類化合物的生物合成,結(jié)果見表3。
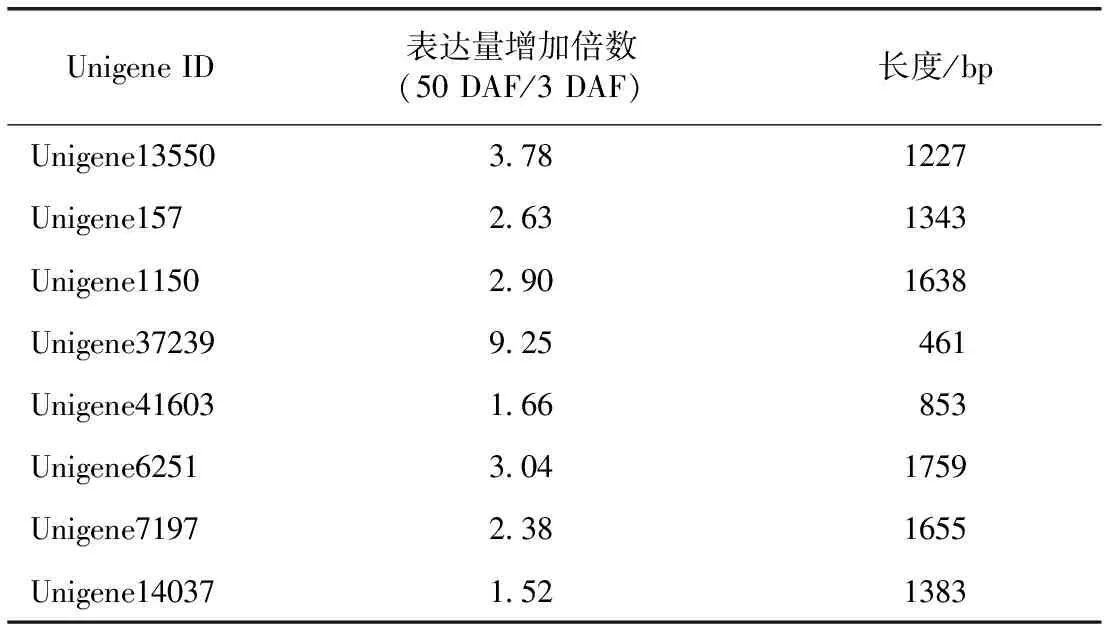
表3 8個(gè)可能參與羅漢果果實(shí)中黃酮類化合物生物合成的MYB轉(zhuǎn)錄因子Unigene
3.2 酵母單雜交的構(gòu)建
3.2.1 總RNA與ds cDNA的質(zhì)量檢測 采用trizol試劑提取羅漢果果實(shí)中的總RNA,凝膠電泳結(jié)果見圖3。從圖中可看到兩條清晰的主帶28S和18S,且28S條帶的亮度大于18S的亮度,表明所提取的RNA無降解現(xiàn)象,質(zhì)量良好。核酸定量檢測結(jié)果顯示總RNA的A260/A280值為1.98,符合要求,濃度為830 ng·μL-1。

注:M.1 kbplus DNAladder;1、2.提取的總RNA。圖3 羅漢果果實(shí)中提取的總RNA電泳分析
將純化的ds cDNA進(jìn)行瓊脂糖凝膠電泳分析,見圖4。從圖4中可看到ds cDNA呈明亮的彌散狀條帶,符合文庫的構(gòu)建要求。

注:M.1kb plus DNA ladder;1.純化的ds cDNA。圖4 羅漢果果實(shí)中純化后的ds cDNA電泳分析
3.2.2 酵母單雜交文庫的質(zhì)量檢測 將10 μL原始電轉(zhuǎn)化菌稀釋100倍后,取10 μL涂板,在平板上共長了約230個(gè)克隆子,共計(jì)5 mL的轉(zhuǎn)化后原始菌液,因此總庫容量為:5×230/10×100×1000=2.3×106cfu。插入片段的凝膠電泳圖見圖5,結(jié)果顯示插入片段的平均長度約1.5 kbp。

注:M.1kb plus DNA ladder;1~24.插入片段。圖5 酵母單雜交文庫插入片段電泳分析
4 討論
隨著生物技術(shù)的發(fā)展,轉(zhuǎn)錄組學(xué)已經(jīng)逐漸成為研究者探索生物體生理生化過程機(jī)制的有效手段。本研究利用不同時(shí)期羅漢果果實(shí)的轉(zhuǎn)錄組數(shù)據(jù)分析果實(shí)中的轉(zhuǎn)錄因子,篩選到4個(gè)與甜苷含量及合成酶基因表達(dá)變化協(xié)同一致的bHLH轉(zhuǎn)錄因子Unigene,提示它們可能參與調(diào)控羅漢果甜苷的合成。接下來可以對它們進(jìn)行克隆表達(dá),利用酵母單雜交、凝膠遷移實(shí)驗(yàn)(electrophoretic mobility shift assay,EMSA)等方法分析它們與甜苷合成酶基因啟動子區(qū)的結(jié)合能力;或?qū)λ鼈冞M(jìn)行RNA干擾及過表達(dá)以確認(rèn)它們的功能。
在羅漢果轉(zhuǎn)錄組數(shù)據(jù)中同時(shí)找到了50個(gè)MYB轉(zhuǎn)錄因子,MYB轉(zhuǎn)錄因子能夠調(diào)控高等植物中黃酮類成分的積累和相關(guān)基因的表達(dá)。在羅漢果果實(shí)中除了含有三萜皂苷外,還含有黃酮類化合物,主要成分為羅漢果素(grosvenorine)和槲皮素苷及它們的苷元山柰酚和槲皮素[13]。對不同時(shí)期羅漢果果實(shí)中總黃酮的含量進(jìn)行測定,發(fā)現(xiàn)其含量先逐漸增長,在50 DAF達(dá)到最大值,然后在60 DAF迅速下降至20 DAF水平[11]。因此本研究利用表達(dá)譜數(shù)據(jù)篩選了在50 DAF/3 DAF組中表達(dá)量上調(diào)的8個(gè)MYB轉(zhuǎn)錄因子Unigene,它們可能參與調(diào)控羅漢果果實(shí)中黃酮類化合物的生物合成。接下來,可以通過克隆已鑒定的羅漢果查爾酮合成酶基因SgCHS的啟動子序列[14],檢測這8個(gè)MYB轉(zhuǎn)錄因子Unigene與SgCHS基因啟動子區(qū)的結(jié)合情況。
在羅漢果的同屬植物黃瓜中,發(fā)現(xiàn)了與氧化鯊烯環(huán)化酶基因Bi(登陸號:KM655855.1)基因啟動子區(qū)E-box結(jié)合的bHLH轉(zhuǎn)錄因子[15],Bi與羅漢果甜苷合成關(guān)鍵酶基因CS(登錄號:HQ128567.1)有86%的同源性,且CS基因啟動子區(qū)也含有E-box,提示羅漢果中可能也存在能夠與CS基因啟動子區(qū)結(jié)合的bHLH類轉(zhuǎn)錄因子。然而,本課題組通過CS基因啟動子區(qū)E-box構(gòu)建的誘餌載體及已成功建立的cDNA文庫,利用酵母單雜交實(shí)驗(yàn)進(jìn)行轉(zhuǎn)錄因子的篩選時(shí),在52個(gè)陽性克隆中未發(fā)現(xiàn)bHLH轉(zhuǎn)錄因子,這可能與所選取的E-box與bHLH轉(zhuǎn)錄因子結(jié)合能力較弱有關(guān):已知能被bHLH蛋白識別的核心序列為E-box(5′-CANNTG-3′),中間兩個(gè)核苷酸的不確定性使E-box有不同的種類,其中80%以上的序列為G-box(5′-CACGTG-3′)[16]。CS基因啟動子區(qū)不含有G-box,而含有不同類型的E-box,有文獻(xiàn)對這些E-box與bHLH蛋白的結(jié)合能力進(jìn)行了檢測,發(fā)現(xiàn)與G-box相比它們的結(jié)合能力下降了50%以上[17]。
本研究成功構(gòu)建了酵母單雜交文庫,有助于通過建立誘餌載體進(jìn)行酵母單雜交實(shí)驗(yàn),篩選與誘餌DNA序列結(jié)合的轉(zhuǎn)錄因子,為尋找羅漢果果實(shí)中具有調(diào)控作用的轉(zhuǎn)錄因子,闡明羅漢果中相關(guān)活動的基因調(diào)控網(wǎng)絡(luò)及分子機(jī)制奠定基礎(chǔ)。
[1] LI C,LIN L M,SUI F,et al.Chemistry and pharmacology ofSiraitiagrosvenorii:a review[J].Chin J Nat Med,2014,12(2):89-102.
[2] Kasai R,Nie R L,Nashi K,et al.Sweet Cucurbitane Glycosides from Fruits ofSiraitiasiamensis(chi-zi luo-han-guo),a Chinese Folk Medicine[J].Agric BiolChem,1989,53(12):3347-3349.
[3] Kosugi S,Ohashi Y.PCF1 and PCF2 Specifically Bind tocisElements in the Rice Proliferating Cell Nuclear Antigen Gene[J].Plant Cell,1997,9(9):1607-1619.
[4] Zhang H B,Bokowiec M T,Rushton P J,et al.Tobacco Transcription Factors NtMYC2a and NtMYC2b Form Nuclear Complexes with the NtJAZ1 Repressor and Regulate Multiple Jasmonate-Inducible Steps in Nicotine Biosynthesis[J].Mol Plant,2012,5(1):73-84.
[5] Zarka D G,Vogel J T,Cook D,et al.Cold Induction of ArabidopsisCBFGenes Involves Multiple ICE (Inducer ofCBFExpression) Promoter Elements and a Cold-Regulatory Circuit That Is Desensitized by Low Temperature[J].Plant Physiol,2003,133(2):910-918.
[6] Qian W Q,Tan G H,Liu H X,et al.Identification of a bHLH-type G-box binding factor and its regulation activity with G-box and Box I elements of thePsCHS1 promoter[J].Plant Cell Rep,2007,26(1):85-93.
[7] Chatel G,Montiel G,Pre M,et al.CrMYC1,aCatharanthusroseuselicitor-and jasmonate-responsive bHLH transcription factor that binds the G-box element of the strictosidine synthase gene promoter[J].J Exp Bot,2003,54(392):2587-2588.
[8] 李書濤.調(diào)控紫杉醇合成轉(zhuǎn)錄因子TcMYC和TcWRKY1的克隆及功能研究[D].武漢:華中科技大學(xué),2012.
[9] Tang Q,Ma X J,Mo C M,et al.An efficient approach to findingSiraitiagrosvenoriitriterpene biosynthetic genes by RNA-seq and digital gene expression analysis[J].BMC Genomics,2011,12:343.
[10] 唐其.羅漢果轉(zhuǎn)錄組、表達(dá)譜的高通量測序及甜苷V生物合成關(guān)鍵酶的克隆[D].北京:北京協(xié)和醫(yī)學(xué)院,2010.
[11] 陳全斌,義祥輝,余麗娟,等.不同生長周期的羅漢果鮮果中甜甙V和總黃酮含量變化規(guī)律研究[J].廣西植物,2005,25(3):274-277.
[12] Liu J Y,Osbourn A,Ma P D.MYB Transcription Factors as Regulators of Phenylpropanoid Metabolism in Plants[J].Mol Plant,2015,8(5):689-708.
[13] 斯建勇,陳迪華,常琪,等.鮮羅漢果中黃酮甙的分離及結(jié)構(gòu)測定[J].藥學(xué)學(xué)報(bào),1994,29(2):158-160.
[14] 王志強(qiáng),蒙姣榮,鄒承武,等.羅漢果查爾酮合成酶基因的生物信息學(xué)分析[J].基因組學(xué)與應(yīng)用生物學(xué),2010,29(3):577-583.
[15] Shang Y,Ma Y S,Zhou Y,et al.Plant science.Biosynthesis,regulation,and domestication of bitterness in cucumber[J].Science,2014,346(6213):1084-1088.
[16] 張鑫,宋經(jīng)元,胡鳶雷,等.bHLH轉(zhuǎn)錄因子調(diào)控植物活性成分生物合成的研究進(jìn)展[J].藥學(xué)學(xué)報(bào),2014,49(4):435-442.
[17] Donmbrecht B,Xue G P,Sprague S J,et al.MYC2 differentially modulates diverse jasmonate-dependent functions inArabidopsis[J].Plant Cell,2007,19(7):2225-2245.
TranscriptomeAnalysisonTranscriptionFactorsofSiraitiagrosvenoriiFruitsatDifferentGrowingStagesandConstructionofYeastOne-HybridLibrary
ZHANG Kailun1,LUOZuliang1,GUOYuhua1,SHIHongwu1,MAXiaojun1,2*
(1.InstituteofMedicinalPlantDevelopment,ChineseAcademyofMedicalSciencesandPekingUnionMedicalCollege,Beijing100193,China;2.YunnanBranchofInstituteofMedicinalPlantDevelopment,ChineseAcademyofMedicalSciences,Jinghong666100,China)
Objective:To study transcription factors (TFs)in the fruits ofSiraitiagrosvenorii.MethodsNon repeating TFs unigenes were selected in transcriptome ofS.grosvenoriifruits at 3,50 and 70 DAF and their expressions were investigated in gene expression profiles;Matchmaker Yeast One-Hybrid Library Construction system was applied to construct yeast one-hybrid cDNA library.Results119 differentially expressed TFs unigenes which belong to 38 TFs families were found out.The numbers of TFs unigenes in 70 DAF/3 DAF and 50 DAF/3 DAF groups were greater and the numbers of the up-regulated and down-regulated TFs unigenes were similar.What’s more,4 bHLH (basic helix-loop-helix,bHLH) TFs unigenes (bHLH014、bHLH025、bHLH093、bHLH096) which might involve in the regulation of mogrosides synthesis and 8 MYB TFs unigenes which might involve in the regulation of flavonoids synthesis were screened.The yeast one-hybrid library pGADT7-Rec2-Lib was successfully constructed with 2.3×106cfu storage capacity.And the inserted PCR fragments sizes are about 1.5 kbp.ConclusionTranscriptome analysis and yeast one-hybrid system are able to help the selection of transcription factors with regulatory effect inS.grosvenoriifruits.
Siraitiagrosvenorii;transcription factor;transcriptome;yeast one-hybrid library
2016-05-04)
國家自然科學(xué)基金(81373914,81573521)
*
馬小軍,研究員,研究方向:分子生物學(xué);E-mail:mayixuan10@163.com
10.13313/j.issn.1673-4890.2016.8.001
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
