重組pIRES-AD7c-NTP質粒對神經細胞氧化應激水平的影響
2017-09-15 13:39:12白春艷孫宏俠胡軼虹李宗樹張明明
中國老年學雜志 2017年16期
關鍵詞:氧化應激
白春艷 孫宏俠 胡軼虹 劉 敏 周 艷 李宗樹 張明明 王 凱
(吉林省人民醫院,吉林 長春 130021)
重組pIRES-AD7c-NTP質粒對神經細胞氧化應激水平的影響
白春艷 孫宏俠 胡軼虹 劉 敏 周 艷 李宗樹 張明明 王 凱
(吉林省人民醫院,吉林 長春 130021)
目的將構建好的重組pIRES-AD7c-NTP質粒轉染至PC12細胞內,探究AD7c-NTP上調對神經細胞氧化損傷的影響。方法MTT實驗檢測重組pIRES-AD7c-NTP質粒轉染組對細胞增殖的抑制作用,研究受損細胞一氧化氮合酶(NOS)和超氧化物歧化酶(SOD)活性。結果重組質粒轉染組可見明顯的細胞增殖抑制作用,NOS活性明顯增高,而SOD活性明顯降低,出現明顯的細胞死亡。結論重組pIREs-AD7c-NTP質粒可導致神經細胞發生氧化損傷。
阿爾茨海默病相關神經絲蛋白;PC12細胞;超氧化物歧化酶;一氧化氮合酶
阿爾茨海默病(AD)相關神經絲蛋白(AD7c-NTP)在AD患者腦內選擇性增高。尿液中AD7c-NTP含量測定可以很好反映AD患者的腦內神經元內該蛋白濃度〔1〕,而且兼有較好的特異性和敏感性,操作簡便,是輔助診斷 AD較好的生物學指標。我們通過構建重組pIRES-AD7c-NTP質粒并表達在PC12細胞內,觀察其對神經細胞的氧化應激水平的影響。氧化應激與AD有著密切的關系〔2〕。本文觀察重組pIRES-AD7c-NTP質粒轉染至PC12細胞內,上調對神經細胞氧化損傷的影響。
1 材料與方法
1.1細胞株與試劑 PC12細胞株購自中國科學院上海細胞庫,辣根過氧化物酶標記羊抗兔IgG購自北京中杉生物技術公司,兔抗人AD7c-NTP抗體購自首都醫科大學宣武醫院中心實驗室,RT-PCR試劑盒由日本TaKaRa公司提供,一氧化氮合酶(NOS)及超氧化物歧化酶(SOD)檢測試劑盒由南京建成生物有限公司提供,MTT由Hyclone公司提供,pIRES-AD7c-NTP質粒由Clontech公司提供,重組表達載體pIRES-AD7c-NTP由本課題組制備。
1.2PC12細胞的培養及轉染 PC12細胞培養于含10%胎牛血清的RPMI-1640培養液中,置于5%的CO2、37℃恒溫孵箱中,2~3 d傳代1次。細胞轉染前24 h,用含10%胎牛血清的無抗生素培養液懸浮,接種在細胞培養板中,密度為5×105/ml。5%CO2,37℃培養18~24 h,使細胞融合達50%~80%。0.5 μl脂質體2000和0.2 μg質粒分別稀釋于25 μl不含血清和抗生素的培養液中,輕輕混勻,室溫靜置5 min。質粒稀釋液中加入上述25 μl脂質體2000稀釋液,輕柔混勻,室溫孵育20 min。將50 μl脂質體2000/質粒混合物滴加到96孔培養板中,輕微搖晃,使培養液混勻。5%CO2、37℃培養箱培養。轉染4 h后細胞換液,除去脂質體,5%CO2、37℃培養箱繼續培養。
1.3脂質體2000介導細胞轉染最佳質粒/脂質體比值的確定 取對數生長期細胞,消化后以5×105/ml的密度接種于96孔培養板中。37℃,5%CO2培養24 h,使細胞融合達50%~80%。棄去細胞培養液,用無血清培養液洗滌細胞1次,備轉染。分別按照一定含量配制脂質體2000/質粒混合物進行轉染。
1.4MTT法檢測細胞增殖變化 實驗共分三組:空白對照組:不轉染,細胞中加入同等體積無血清培養基;轉染pIRES-EGFP空白質粒的陰性對照組:用空質粒pIRES-EGFP/脂質體2000混合物進行轉染;pIRES-AD7c-NTP重組質粒轉染組:用重組載體pIRES-AD7c-NTP/脂質體2000混合物進行轉染。細胞增殖抑制率=(對照組A值-實驗組A值)/對照組A值×100%。
1.5NOS含量測定 將對數生長期細胞接種于6孔培養板中,分組進行轉染(方法同前)。轉染48 h后,取各組細胞培養上清液,按NOS試劑盒說明加入試劑,在紫外分光光度計(530 nm)測定吸光度,按公式計算培養液中NOS含量。
1.6SOD含量測定 對數生長期細胞接種于6孔培養板中,分組進行轉染(方法同前)。48 h后,收集各組細胞,用Triton X-100進行破碎,分別裝入5 ml有蓋試管中,按照試劑盒說明書加入各試劑,在紫外分光光度計(550 nm)測定吸光度,按公式計算培養液中的SOD含量。
1.7統計學方法 采用SPSS11.0統計軟件行t檢驗。
2 結 果
2.1各處理組細胞增殖情況比較 PC12細胞轉染后48~72 h,倒置顯微鏡下觀察細胞形態變化,陰性對照組細胞生長狀態良好,細胞貼壁生長,表面光滑,折光性好,呈多角形生長,細胞表面形成突起,清晰可見,細胞增殖旺盛;重組質粒轉染組,隨著轉染時間延長,細胞逐漸變圓,胞質回縮,細胞突起消失,折光性差,細胞膜表面不光滑,出現發泡現象,胞質中出現顆粒狀物質,部分細胞出現死亡。空白與對照(0.892±0.004)組相比,陰性對照組細胞體外增殖活性無明顯差異(0.886±0.003,P>0.05),其抑制率為0.67%;重組質粒轉染組可見明顯的抑制細胞增殖作用細胞增殖為0.564±0.003,細胞生長抑制率為36.77%。
2.2各組細胞NOS及SOD活性比較 與空白對照組比較,重組質粒轉染組中NOS活性明顯增高(P<0.05),而陰性對照組與空白對照組無顯著差異(P>0.05)。與空白對照組比較,重組質粒轉染組中SOD活性明顯降低(P<0.05),而陰性對照組與空白對照組無顯著差異(P<0.05)。見表1。
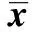
表1 各組細胞NOS、SOD活性的比較(±s,n=3)
與空白對照組比較:1)P<0.05
3 討 論
目前AD的病因及其病理機制仍未明確,存在多種學說和假設,包括氧化應激和線粒體缺陷學說、Aβ學說、膽堿能學說、基因學說、晚期糖基化終產物的諸多學說,國內外學者以此為依據形成了不同的診斷策略。在眾多學說中,氧化應激和線粒體缺陷學說占據著重要的地位〔3~6〕。腦組織病理檢查證明,AD患者腦內海馬和皮質處存在大量的神經元凋亡,所以盡管AD的發病機制非常復雜,一種假說難以完全解釋清楚,但腦內線粒體缺陷和氧化應激導致的神經元退變被認為是發生在AD最早期的事件,是造成AD的最重要的病理因素之一。因此,從氧化應激和線粒體缺陷角度研究AD的發病機制,將為AD的診斷和治療提供新的思路。
正常情況下,機體內活性氧(ROS)產生和清除處于動態平衡,當ROS蓄積過多就會攻擊機體,ROS產生過多是氧化應激的主要原因。在生理狀態下的ROS具有刺激細胞生長的作用;而在外界各種因素作用(如Aβ沉積、鐵離子生成等)下,ROS產生和清除之間的動態平衡就會被打破,就會導致大量ROS在體內堆積,對神經系統造成損傷。在AD患者腦內,特別是在神經元纖維纏結(NFT)中,發現了大量氧化應激終產物〔7~9〕如:丙二醛、過氧化亞硝酸鹽、羰基、糖基化終末產物、血紅素氧合酶-1和SOD-1等;AD患者尸檢資料也證實,在NFTs形成部位存在著蛋白質氧化產物、羰基和脂質過氧化物的增加,這都提示了自由基所介導的氧化應激參與了AD患者特征性病理結構的形成過程。
體外實驗表明,胰島素刺激SH-Sy5y和PNET 2細胞會引起內源性AD7c-NTP表達的增加和磷酸化反應增強〔10〕。另有學者發現,在AD7c-NTP轉染的細胞中,受損的胰島素/胰島素樣生長因子(IGF)-1信號通路可能介導了神經元的死亡,而且中樞神經元如果含有大量胰島素或IGF-1受體,可能對AD7c-NTP引起的損害效應尤其易感〔11〕。人類組織學研究也揭示,AD7c-NTP在AD腦顳葉和額葉的表達水平較對照組高,并且通過原位雜交和免疫組化染色發現,在組織學尚完整的變性神經元中出現 AD7c-NTP表達的增加〔10〕。所以AD7c-NTP不僅是AD神經元變性的一種標記分子,也在AD發病機制中發揮著重要作用。
本文結果顯示,AD7c-NTP的過度表達可導致細胞凋亡及氧化損傷。關于氧自由基與AD7c-NTP的關系以及AD7c-NTP功能的研究目前尚不多見,它究竟是突變基因表達的產物,還是蛋白質轉運和加工過程異常累積的結果,它在腦的正常功能中占據什么樣的地位,在AD病理改變中發揮什么樣的作用以及導致細胞凋亡的原因,有待于進一步研究。
1Goodman I,Golden G,Fliman S,etal.A multicenter blinded prospective study of urine neural thread protein measurements in patients with sus-pected Alzheimer's disease〔J〕.J Am Med Dir Assoc,2007;8(1):21-30.
2Fernandez AP,Pozo-Rodrigalvarez A,Serrano A,etal.Nitric oxide:target for therapeutic strategies in Alzheimer's disease〔J〕.Curr Pharm Des,2010;16(25):2837-50.
3Onyango IG,Khan SM.Oxidative stress,mitochondrial dysfunction and stress signaling in Alzheimer's disease〔J〕.Curr Alzheimer Res,2006;3(4):339-49.
4Moreira PI,Duarte AI,Santos MS,etal.An integrative view of the role of oxidative stress,mitochondria and insulin in Alzheimer's disease〔J〕.J Alzheimers Dis,2009;6(4):741-61.
5Zhu DL,Shi JP,Zhang YD,etal.Central angiotensin Ⅱ stimulation promotes β amyloid production in Sprague Dawley rats〔J〕.PLoS One,2011;6(1):el6037.
6Li S,Wang W,Wang C,etal.Possible involvement of NO/NOS signaling in hippocampal Amyloid-production induced by transient focal cerebral ischemia in aged rats〔J〕. Neurosci Lett,2010;470(2):106-10.
7Gilgun-Sherki Y,Melamed E,Offen D.Antioxidant treatment in Alzheimer's disease:current state〔J〕.J Mol Neurosci,2003;21(1):1-11.
8Polidori MC.Oxidative stress and risk factors for Alzheimer's disease:clues to prevention and therapy〔J〕.J Alzheimer Dis,2004;6(2):185-91.
9Lakshmi BV,Sudhakar M,Prakash KS.Protective effect of selenium against aluminum chloride-induced Alzheimer's disease:behavioral and biochemical alterations in rats〔J〕.Biol Trace Elem Res,2015;12(1):125-31.
10de la Monte SM,Ghanbari K,Frey WH,etal.Characterization of the AD7C-NTP cDNA expression in Alzheimer's disease and measurement of a 41-kD protein in cerebrospinalfluid〔J〕.J Clin Invest,1997;100(3):3093-104.
11de la Monte SM,Wands JR.Alzheimer-associated neuronal thread protein mediated cell death is linked to impaired insulin signaling〔J〕.J Alzheimers Dis,2004;6(1):231-42.
〔2016-02-03修回〕
(編輯 苑云杰/曹夢園)
吉林省科委課題(No.200705464)
孫宏俠(1964-),女,碩士,教授,碩士生導師,主要從事認知障礙及腦血管病研究。
白春艷(1974-),女,主任醫師,醫學博士,碩士生導師,主要從事周圍神經疾病研究。
R749
A
1005-9202(2017)16-3948-02;doi:10.3969/j.issn.1005-9202.2017.16.017
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
