天人菊與金雞菊幼苗生長生理特性比較
2018-01-12 11:39:25胡妍妍駱建霞曾麗榮于學(xué)江
江蘇農(nóng)業(yè)科學(xué) 2017年24期
胡妍妍, 張 希, 王 丹, 駱建霞, 曾麗榮, 于學(xué)江
(1.天津農(nóng)學(xué)院園藝園林學(xué)院,天津 300384; 2.天津正特園林綠化工程有限公司,天津 300060)
種間競爭是不同種群為爭奪生活空間、資源、食物等而產(chǎn)生的一種直接或間接抑制對方生長的現(xiàn)象。在有限的空間、土壤、水肥等條件下,花卉近距離種植,其相互之間的關(guān)系是競爭還是促進、能否和諧生長而保持良好穩(wěn)定的搭配形式關(guān)系到花卉的整體景觀效果。天人菊(Gaillardiapulchella)和金雞菊(Coreopsisdrummondii)是園林常見的2種花卉,景觀效果好、花色艷麗,可在北方露地越冬,且管理相對粗放,適合花境、花帶種植而廣泛應(yīng)用于各種綠地中。
目前,對菊科植物種間競爭的研究已取得一定進展[1-5],但多集中在菊科植物對其他植物種子萌發(fā)或生理特性的影響,而對不同菊科植物之間的競爭及從種子萌發(fā)到幼苗生長各生理指標(biāo)變化的研究鮮見報道。本試驗通過將同為菊科植物的天人菊與金雞菊進行混種,測定混種與單種時其種子的發(fā)芽情況和幼苗生長各項生理指標(biāo),分析菊科植物間的競爭效應(yīng),為花境植物材料的選擇、物種間的搭配、植物整體景觀效果設(shè)計等提供理論指導(dǎo),為觀賞植物的種間競爭研究提供參考。
1 材料與方法
1.1 材料
天人菊和金雞菊種子,采收于天津農(nóng)學(xué)院內(nèi),其栽培土由采自天津農(nóng)學(xué)院內(nèi)的園土與草炭土按體積1 ∶1配成,用盆口直徑為18 cm、深20 cm的塑料花盆裝入等量栽培土。
1.2 試驗處理
1.2.1 種子發(fā)芽試驗 試驗于2015年春夏在天津農(nóng)學(xué)院園林規(guī)劃實驗室進行,選擇飽滿、大小均勻一致的宿根天人菊和金雞菊種子,用0.1%高錳酸鉀溶液消毒5 min;無菌水沖洗,晾干,將天人菊與金雞菊種子按粒數(shù)比40 ∶0、25 ∶15、20 ∶20、15 ∶25、0 ∶40進行配比,分別播種于鋪有雙層濾紙的培養(yǎng)皿中,每個培養(yǎng)皿40粒種子;25 ℃恒溫箱中進行培養(yǎng),每天更換清水,統(tǒng)計種子發(fā)芽數(shù)量,當(dāng)連續(xù)3 d無新發(fā)芽種子出現(xiàn)時視為發(fā)芽過程結(jié)束。以胚根長為種子長的1/2為發(fā)芽標(biāo)準(zhǔn),以播種單一植物(單種)為對照。重復(fù)3次。參照鄭光華的方法[6]計算發(fā)芽勢、發(fā)芽率:
發(fā)芽勢=(7 d內(nèi)發(fā)芽的種子數(shù)/供試種子數(shù))×100;
發(fā)芽率=(發(fā)芽種子數(shù)/供試種子數(shù))×100%。
1.2.2 幼苗生長試驗 選出整齊一致的天人菊和金雞菊幼苗,按株數(shù)比1 ∶1混種栽入花盆中,每盆種植20株,重復(fù)5次,以單獨種植(單種)為對照,試驗期間確保各試驗苗受光均勻。幼苗生長培養(yǎng)80 d,從單種、混種盆中分別隨機選取天人菊和金雞菊幼苗15株,測定其根、莖、葉鮮質(zhì)量,用畫紙稱質(zhì)量法測定總?cè)~面積,計算比葉面積、根冠比。同時,分別用丙酮提取法、氯化三苯基四氮唑(TTC)法、氮藍四唑(NBT)法、愈創(chuàng)木酚法測定葉綠素含量、根系活力、超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性[7]。比葉面積(specific leaf area,SLA)是指單位干質(zhì)量的鮮葉表面積,計算公式為:
SLA=S/A。
式中:S為葉面積,cm2;A為葉干質(zhì)量,g。根冠比計算公式為:
根冠比=根鮮質(zhì)量/地上部鮮質(zhì)量。
1.3 數(shù)據(jù)分析
采用Excel 2003、SPSS 18.0軟件對試驗數(shù)據(jù)進行統(tǒng)計分析。
2 結(jié)果與分析
2.1 不同播種配比對天人菊和金雞菊種子發(fā)芽特性的影響
由表1可知,隨金雞菊種子使用比例的增加,天人菊種子的發(fā)芽率和發(fā)芽勢呈明顯的先升后降趨勢;混種時,天人菊種子的發(fā)芽率和發(fā)芽勢顯著高于單種(P<0.05);天人菊和金雞菊播種粒數(shù)比為20 ∶20時,天人菊種子的發(fā)芽率和發(fā)芽勢相對最高,分別為74.43%、74.47,分別為單種時的1.40、1.41倍;隨天人菊種子使用比例的增加,金雞菊種子的發(fā)芽率、發(fā)芽勢也呈明顯的先升后降趨勢;天人菊和金雞菊播種粒數(shù)比為15 ∶25時,金雞菊種子的發(fā)芽率、發(fā)芽勢相對最高,分別為45.00%、43.84,顯著高于單種(P<0.05);天人菊和金雞菊播種粒數(shù)比為25 ∶15時,金雞菊種子的發(fā)芽率、發(fā)芽勢顯著降低(P<0.05),分別是單種時的0.76、0.84倍。天人菊與金雞菊混種有利于天人菊種子的萌發(fā),適當(dāng)配比的混種有利于金雞菊種子發(fā)芽,而當(dāng)天人菊播種比例過高時,金雞菊種子的發(fā)芽率和發(fā)芽勢會降低。
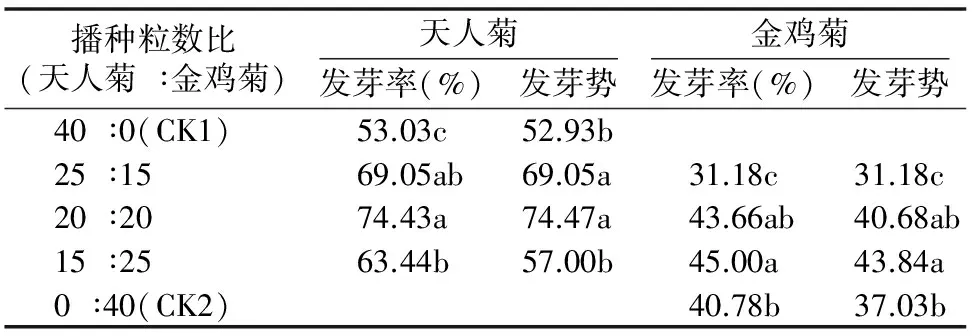
表1 天人菊和金雞菊不同播種配比時的發(fā)芽率、發(fā)芽勢情況
注:同列數(shù)據(jù)后不同小寫字母表示處理間差異顯著(P<0.05)。
2.2 單種與混種對天人菊和金雞菊株高、植株干質(zhì)量的影響
由表2可知,天人菊混種時的株高顯著高于單種(P<0.05),是單種的1.39倍;金雞菊混種和單種時的株高差異不顯著(P>0.05);天人菊混種時的植株干質(zhì)量極顯著高于單種(P<0.01),是單種的1.65倍,而金雞菊表現(xiàn)則相反,金雞菊混種時的植株干質(zhì)量極顯著低于單種(P<0.01),僅為單種的0.39倍。天人菊和金雞菊混種對各自生長產(chǎn)生較為明顯的影響,天人菊更具競爭優(yōu)勢。
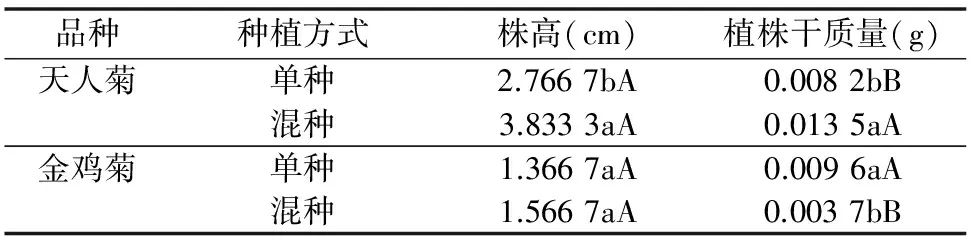
表2 天人菊和金雞菊在單種和混種時的株高和植株干質(zhì)量
注:同一品種同列數(shù)據(jù)后不同小寫、大寫字母表示處理間差異顯著(P<0.05)、極顯著(P<0.01)。下同。
2.3 單種與混種對天人菊和金雞菊比葉面積和根冠比的影響
植物比葉面積越大,即單位干質(zhì)量的葉面積越大,葉片越薄,越有利于捕獲更多的光能,使植物具有更快生長速度的可能性[8]。由表3可知,天人菊混種、單種時的葉面積和葉質(zhì)量變化差異不顯著(P>0.05),而比葉面積、根質(zhì)量、根冠比差異極顯著(P<0.01),天人菊混種時的比葉面積比單種增加4.97%,而根質(zhì)量、根冠比分別比單種下降68.42%、64.52%,混種時天人菊根的生長受到明顯影響;金雞菊混種與單種相比,除根冠比差異不顯著(P>0.05)外,金雞菊混種時的葉面積是單種的1.50倍,顯著高于單種(P<0.05),葉質(zhì)量和根質(zhì)量分別是單種的2.67、3.00倍,極顯著高于單種(P<0.01),而比葉面積是單種的0.56倍,極顯著低于單種(P<0.01),混種時金雞菊比葉面積降低,其單位干質(zhì)量的葉片面積減小,而葉質(zhì)量的增加說明植物葉片厚度增加。
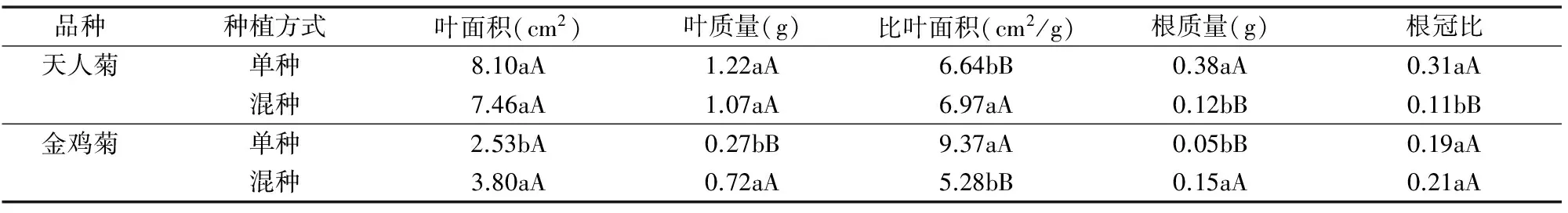
表3 天人菊和金雞菊在單種和混種時的比葉面積和根冠比
2.4 單種與混種對天人菊和金雞菊葉綠素、類胡蘿卜素含量和根系活力的影響
類胡蘿卜素除在光合作用中收集和傳遞光能外,在細胞內(nèi)還起到清除氧自由基的作用,以防止細胞膜脂過氧化[9]。由表4可知,天人菊單種、混種時的色素含量、根系活力差異不顯著(P>0.05);金雞菊混種時的葉綠素含量比單種提高33.52%,極顯著高于單種(P<0.01),類胡蘿卜素素含量、根系活力分別比單種提高23.07%、155.95%,顯著高于單種(P<0.05),這說明混種時的金雞菊可能通過提高色素合成、根系活力來提高與天人菊的競爭。
2.5 單種與混種對天人菊和金雞菊抗氧化酶活性的影響
由表5可知,天人菊混種時的SOD活性與單種相比差異不顯著(P>0.05),而POD活性顯著降低(P<0.05),比單種低41.35%;金雞菊混種時的SOD活性比單種顯著上升37.67%(P<0.05),而POD活性比單種極顯著下降83.50%(P<0.01)。這說明天人菊的抗氧化酶活性受栽種環(huán)境影響相對較小,而混種時的金雞菊SOD活性升高、POD活性下降,金雞菊生長受到一定的抑制作用。
3 結(jié)論與討論
植物間的競爭既存在促進作用,又存在抑制作用[3,10]。
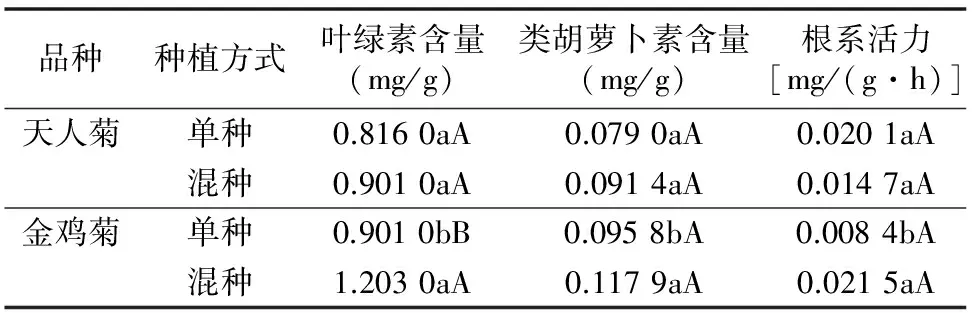
表4 天人菊和金雞菊在單種和混種時的葉綠素、類胡蘿卜素

表5 天人菊和金雞菊在單種和混種時的SOD和POD活性
天人菊與金雞菊混種時,天人菊種子的發(fā)芽率、發(fā)芽勢較單種時有所提高,說明其種子萌發(fā)受到一定的促進作用。而當(dāng)天人菊播種比例過高時,金雞菊種子的發(fā)芽率和發(fā)芽勢會降低,此時金雞菊種子的萌發(fā)會受到一定的抑制作用。天人菊與金雞菊混種時,天人菊株高、植株干質(zhì)量顯著或極顯著高于單種,而根質(zhì)量、根冠比極顯著低于單種(P<0.01),說明混種時的天人菊地上器官(莖、葉)生長相對較快,消耗同化產(chǎn)物相對較多,供應(yīng)地下部根系生長的營養(yǎng)物質(zhì)相對較少,導(dǎo)致根冠比下降;混種時的金雞菊葉面積、葉質(zhì)量、根質(zhì)量顯著或極顯著高于單種,比葉面積相對降低,說明金雞菊葉片厚度增加。天人菊株高高于金雞菊,具有一定的競爭優(yōu)勢,天人菊與金雞菊混種時,金雞菊可能通過增加葉片厚度、促進根系生長來保證其與天人菊的競爭,從而反映出混種時金雞菊生長受到一定抑制,通過自身調(diào)節(jié)以保證其正常生長。超氧化物歧化酶(SOD)和過氧化物酶(POD)的作用是清除活性氧而避免對細胞造成傷害。混種時的天人菊SOD活性與單種相比差異不顯著(P>0.05),而POD活性顯著低于單種(P<0.05),而金雞菊混種時為適應(yīng)環(huán)境而啟動防御系統(tǒng)酶,SOD活性顯著上升37.67%(P<0.05),POD活性極顯著下降83.50%(P<0.01),說明植物在逆境生長中起保護作用的各種酶是相互協(xié)調(diào)、配合作用的[11-14]。
金雞菊和天人菊混種時,金雞菊對天人菊種子的萌發(fā)起到一定的促進作用,對其幼苗生長影響不大,而天人菊對金雞菊種子的萌發(fā)及幼苗生長均起到一定的抑制作用。因此,在實際栽植過程中,金雞菊和天人菊的搭配方式和入地栽植數(shù)量有待進一步研究。
[1]杜 峰,梁宗鎖,胡莉娟. 植物競爭研究綜述[J]. 生態(tài)學(xué)雜志,2004,23(4):157-163.
[2]徐高峰,張付斗,李天林,等. 不同密度五種植物對薇甘菊幼苗的競爭效應(yīng)[J]. 生態(tài)環(huán)境學(xué)報,2011,20(5):798-804.
[3]慕小倩,羅瑪霞,段琦梅,等. 10種菊科植物水浸液對小麥幼苗生長的影響[J]. 西北植物學(xué)報,2003,23(11):2014-2017.
[4]杜明利,高 巖,張汝民,等. 大花金雞菊水浸液對6種常見園林植物種子萌發(fā)的化感作用[J]. 浙江農(nóng)林大學(xué)學(xué)報,2011,28(1):109-114.
[5]王乃亮,馬瑞君,孫 坤,等. 5種菊科植物水浸液對黃帚橐吾種子萌發(fā)期化感作用研究[J]. 西北植物學(xué)報,2003,23(11):2014-2017.
[6]鄭光華. 種子生理研究[M]. 北京:科學(xué)出版社,2004:623-624.
[7]張治安,張美善,蔚榮海. 植物生理學(xué)實驗指導(dǎo)[M]. 北京:中國農(nóng)業(yè)出版社,2004:138-145.
[8]張 林,羅天祥. 植物葉壽命及其相關(guān)葉性狀的生態(tài)學(xué)研究進展[J]. 植物生態(tài)學(xué)報,2004,28(6):844-852.
[9]Willekens H,van Camp W,van Montagu M,et al. Ozone,sulfur dioxide,and ultraviolet B have similar effects on mRNA accumulation of antioxidant genes inNicotianaplumbaginifoliaL.[J]. Plant Physiology,1994,106(3):1007-1014.
[10]郭榮群,趙 宏,張 柯,等. 劍葉金雞菊水浸提液對種子萌發(fā)的化感作用[J]. 北方園藝,2010(4):45-48.
[11]杜國平,鄒 青,連芳青,等. 黑心菊抗旱性生理生化指標(biāo)的研究[J]. 江西農(nóng)業(yè)學(xué)報,2012,24(12):42-45.
[12]高福元,張吉立,劉振平. 冬季低溫對4種彩葉植物SOD、POD活性影響的研究[J]. 中國農(nóng)學(xué)通報,2010,26(5):169-173.
[13]王 鑫,朱 悅,劉濱碩,等. 鹽堿脅迫下羊草抗氧化酶活性的變化[J]. 江蘇農(nóng)業(yè)科學(xué),2015,43(5):209-211.
[14]田丹青,葛亞英,潘剛敏,等. 低溫脅迫對3個紅掌品種葉片形態(tài)和生理特性的影響[J]. 園藝學(xué)報,2011,38(6):1173-1179.
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44

- 江蘇農(nóng)業(yè)科學(xué)的其它文章
- 農(nóng)業(yè)科研單位科技報告質(zhì)量控制評價及提升對策
- 江蘇省草莓生產(chǎn)現(xiàn)狀調(diào)查及經(jīng)濟效益分析
- 農(nóng)業(yè)高等職業(yè)院校教師職業(yè)認(rèn)同與教師效能感、應(yīng)對方式的關(guān)系分析
- 農(nóng)業(yè)供給側(cè)結(jié)構(gòu)性改革及實現(xiàn)形式
——田園綜合體 - 基于Malmquist模型下的農(nóng)產(chǎn)品流通模式優(yōu)化
- 精準(zhǔn)扶貧:現(xiàn)狀、問題與路徑優(yōu)化
——基于宿州市9個典型鄉(xiāng)鎮(zhèn)的調(diào)查