豬感染PRRSV后Cathelicidins抗菌肽基因的表達情況
2018-03-12 07:28:34盧艷敏
江蘇農(nóng)業(yè)科學 2018年2期
盧艷敏
(衡水學院生命科學系,河北衡水 053000)
抗菌肽是動物天然免疫反應(yīng)的重要組成部分,具有廣譜抗菌活性,對細菌、真菌、原蟲和病毒均具有較強的殺傷作用[1],另外還可以抗癌、促進傷口愈合,因此具有較大的藥用開發(fā)價值。抗菌肽是生物體經(jīng)誘導產(chǎn)生的具有抗菌活性的小分子多肽,在機體抵抗病原入侵中起著重要作用[2],具有分子量小、抗菌譜廣、抗菌活性高、不易使病原菌產(chǎn)生耐藥性、不破壞動物體細胞、無免疫原性等優(yōu)點。抗菌肽作用機制獨特,主要有膜作用機制和胞內(nèi)作用機制。最新研究表明,抗菌肽還存在其他的殺菌機制,Chu等研究發(fā)現(xiàn),抗菌肽α-defensin 6可以結(jié)合到鼠傷寒沙門氏菌表面,通過自我組裝,在細菌周圍形成一些小纖維和納米樣纖維,從而減少細菌黏附到腸道黏膜,以達到保護機體的目的[3]。汪以真發(fā)現(xiàn),豬源hepcidin能夠使大腸桿菌K88聚集成團,形成網(wǎng)狀結(jié)構(gòu)包裹[4]。
豬源抗菌肽主要包括防御素(defensins)和cathelicidins兩大家族,目前已發(fā)現(xiàn)了約30種豬源抗菌肽,其中Cathelicidins有PR-39、PG1-5(protegrin)、PF1-2(prophenin)和PMAP等4類抗菌肽[1]。cathelicidins基因由4個外顯子和3個內(nèi)含子組成,1~3個外顯子編碼N-端信號肽區(qū)域(PRE)和保守的cathelin區(qū)域(PRO),第4個外顯子編碼Pro區(qū)的最后幾個殘基和高度特異的C-端成熟肽區(qū)域。與其他抗菌肽相比,cathelicidins家族的抗菌活性更強[5-6],抗菌作用迅速[7],對耐藥菌株也具有較強的活性[8]、低的溶血活性以及細胞毒性,在新型抗菌藥物開發(fā)領(lǐng)域顯示出巨大的潛力[9]。
豬繁殖與呼吸綜合征(PRRS)是豬的一種高度傳染性疾病,最早在北美洲和歐洲發(fā)現(xiàn),我國于1955年首次分離到豬繁殖與呼吸綜合征病毒(PRRSV)。PRRSV可引起妊娠母豬流產(chǎn)、死胎、木乃伊胎及產(chǎn)弱仔等繁殖障礙,并可引起各年齡階段豬的呼吸道癥狀,仔豬死亡率高,同時還會導致免疫抑制和持續(xù)性感染等癥狀。PRRSV對我國乃至世界養(yǎng)豬業(yè)造成了巨大的經(jīng)濟損失,已成為嚴重威脅養(yǎng)豬業(yè)發(fā)展的重要傳染病之一,而現(xiàn)有疫苗及藥物效果均不理想[10]。本試驗主要研究豬感染PRRSV后體內(nèi)cathelicidins抗菌肽的表達情況,為PRRSV新藥物的開發(fā)提供思路。
1 材料與方法
1.1 材料
1.1.1 試驗材料 PRRSV由國家野生動物疫病研究中心保存,PRRSV感染1月齡無特定病原體(SPF)豬(24頭),分別于感染后12、24、72、168 h取樣(以未感染病毒的1月齡SPF豬6頭作為對照),取肺臟、肺門淋巴結(jié)、脾臟、肝臟、骨髓等組織,將樣品置于凍存管中,液氮速凍,-80 ℃保存。
1.1.2 試劑 超純RNA提取試劑盒(Ultrapure RNA Kit),購自北京康為世紀生物科技有限公司;SupermoⅢM-MLV反轉(zhuǎn)錄酶,購自北京百泰克生物技術(shù)有限公司;dNTP Mixture、Ribonuclease Inhibitor和Oligo(dT)18Primers,購自寶生物工程(大連)有限公司(TaKaRa);TransStart Green qPCR SuperMix,購自北京全式金生物技術(shù)有限公司。
1.2 方法
1.2.1 RNA的提取及cDNA的合成 采用超純RNA提取試劑盒(Ultrapure RNA Kit)取凍存組織進行總RNA提取,操作過程按照試劑盒說明書進行。瓊脂糖凝膠電泳檢測RNA的質(zhì)量,紫外分光光度計下測定RNA濃度,隨后進行反轉(zhuǎn)錄,合成cDNA第一鏈。以合成的cDNA作為Real-Time PCR(即實時定量PCR)分析的模板,-20 ℃保存?zhèn)溆谩?/p>
1.2.2 Real-Time PCR Real-Time PCR(實時定量PCR)引物(由上海英駿生物技術(shù)有限公司合成)序列[1,11]見表1,按照北京全式金生物技術(shù)有限公司TransStart Green qPCR SuperMix說明書進行Real-Time PCR,反應(yīng)體系:P10.5 μL,P20.5 μL,2×TransStartTMGreen qPCR SuperMix 12.5 μL,Passive Reference Dye 0.5 μL,模板1.0 μL,ddH2O 10.0 μL。反應(yīng)程序:95 ℃ 30 s;95 ℃ 5 s,55 ℃ 15 s,72 ℃ 10 s,40個循環(huán)。相對定量分析參照2-ΔΔCT法[12]。實時定量PCR分析共進行3次生物學重復。

表1 Cathelicidin引物序列
2 結(jié)果與分析
2.1 PR39的表達情況
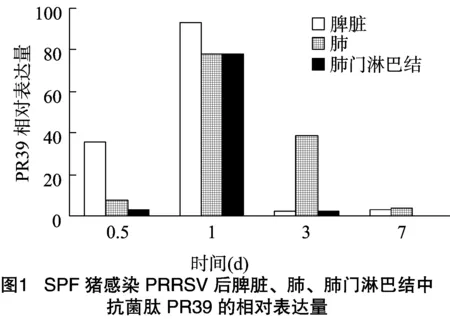
豬感染PRRSV之后,在脾臟、肺、肺門淋巴結(jié)中均檢測到PR39的表達,且隨著感染時間的增長,表達量也有所變化。如圖1所示,豬感染PRRSV后,PR39在脾臟中的表達量在 0.5、1、3和7 d的4個感染時間段中,感染1 d后表達量最高,其次為感染0.5 d;豬感染PRRSV后,PR39在肺中的表達量在0.5、1、3和7 d的4個感染時間段中,感染1 d后表達量最高,其次為感染3 d;豬感染PRRSV后,PR39在肺門淋巴結(jié)中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染1 d后表達量最高,然后依次是感染0.5、3、7 d,但感染0.5 d和 3 d 后的表達量差異不大。
綜上所述,豬感染PRRSV后,PR39在肺、肺門淋巴結(jié)和脾臟3種組織中的表達量均呈現(xiàn)先升高后降低的趨勢,且都是在感染1 d后表達量達到最高值。
2.2 Progerin 1的表達情況
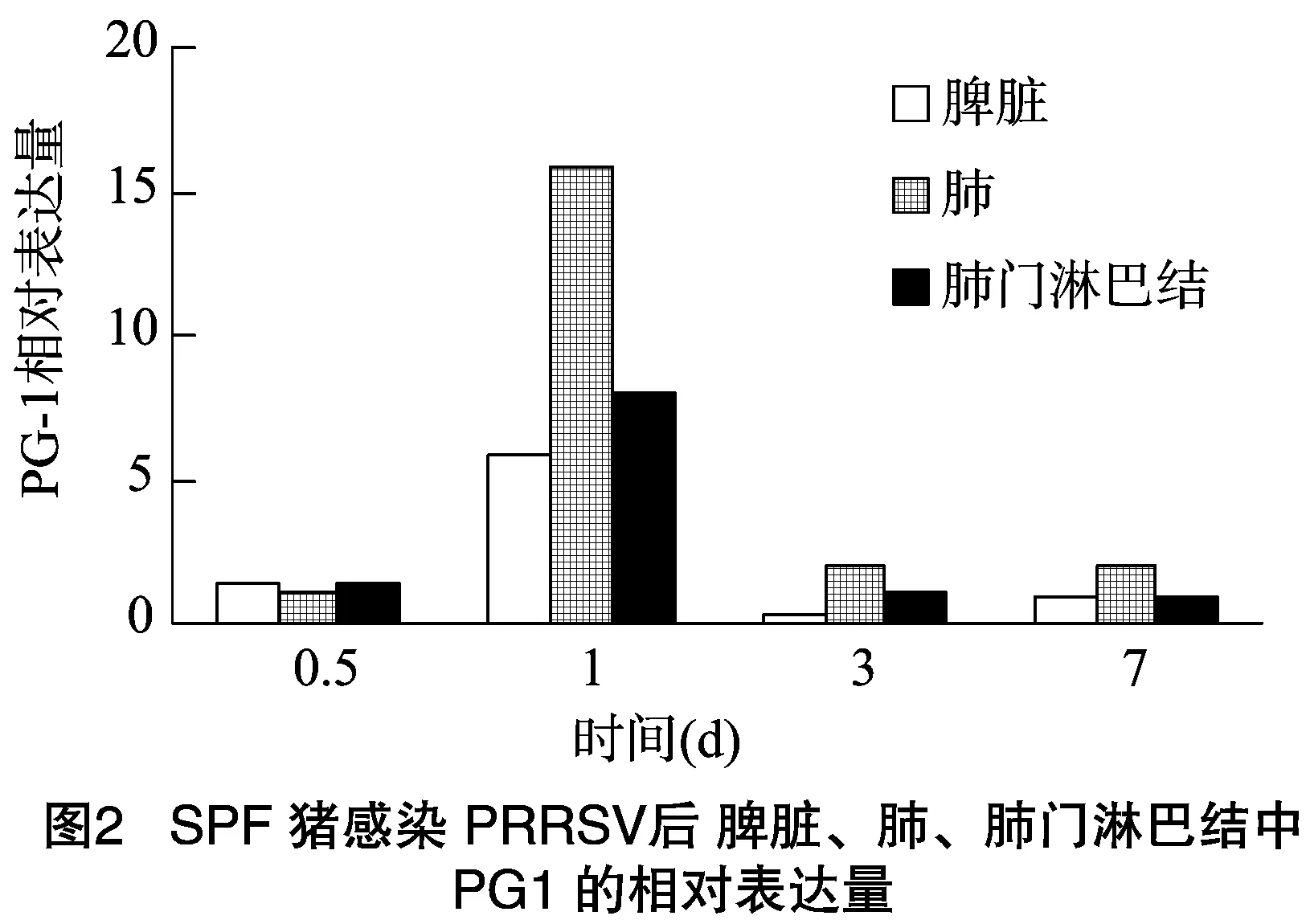
豬感染PRRSV之后,在脾臟、肺、肺門淋巴結(jié)中均檢測到Progerin1(PG1)的表達,感染時間不同,其表達量有所變化,但差異較小。如圖2所示,豬感染PRRSV后,Progerin1在脾臟中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染 1 d 后表達量最高,其次為感染0.5、7、3 d;豬感染PRRSV后,Progerin1在肺中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染1 d后表達量最高,其次為感染3、7、0.5 d;豬感染PRRSV后,Progerin1在肺門淋巴結(jié)中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染1 d后表達量最高,然后依次是感染0.5、3、7 d。綜上所述,豬感染PRRSV后,Progerin1在脾臟、肺和肺門淋巴結(jié)3種組織中的表達量均呈現(xiàn)先升高后降低的趨勢,且都是在感染1 d后表達量達到最高值,Progerin1的表達量在3種組織中的變化均不明顯。
2.3 progerin 1-5的表達情況
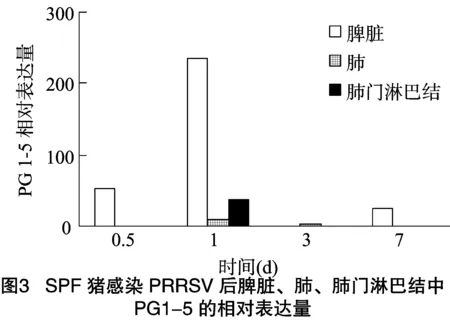
豬感染PRRSV后,在脾臟、肺、肺門淋巴結(jié)中均檢測到Progerin1-5(PG1-5)的表達,在脾臟中表達量較高,而在肺與肺門淋巴結(jié)中表達量較少,隨著感染時間的增長,表達量也有所變化。如圖3所示,豬感染PRRSV后,progerin1-5在脾臟中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染 1 d 后表達量最高,然后依次為0.5、7、3 d;豬感染PRRSV后,progerin1-5在肺中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染1 d后表達量最高;豬感染PRRSV后,progerin1-5在肺門淋巴結(jié)中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染1 d后表達量最高,然后依次是感染0.5、3、7 d,但感染3 d與7 d后的表達量差異不大。
綜上所述,豬感染PRRSV后,progerin1-5在脾臟、肺和肺門淋巴結(jié)3種組織中的表達量均呈現(xiàn)先升高后降低的趨勢,且都是在感染1 d后表達量達到最高值,其中在脾臟中的表達量變化較大,而在肺中的表達量變化不明顯。
2.4 PMAP23的表達情況
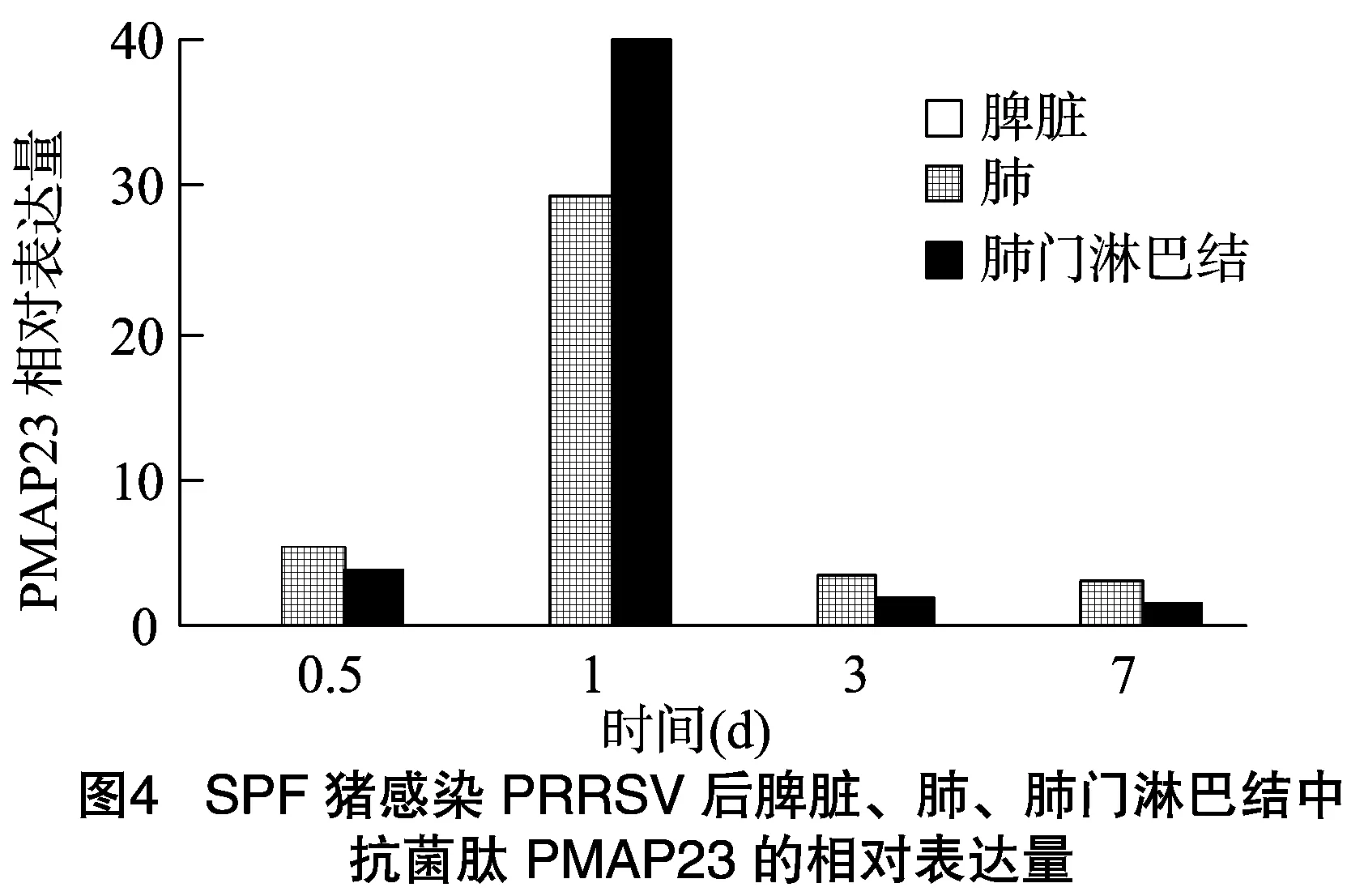
豬感染PRRSV之后,在脾臟中未檢測到PMPA23的表達,肺、肺門淋巴結(jié)中檢測到PMPA23的表達,且隨著感染時間的增長,肺、肺門淋巴結(jié)中PMAP23的表達量有所變化。如圖4所示,豬感染PRRSV后,PMAP23在肺、肺門淋巴結(jié)中的表達量在0.5、1、3和7 d 4個感染時間段中,在感染1 d后表達量最高,然后依次為感染0.5、3、7 d。
綜上所述,豬感染PRRSV后,PMAP23在肺、肺門淋巴結(jié)中的表達量均呈現(xiàn)先升高后降低的趨勢,且都是在感染1 d后表達量達到最高值。
3 討論
抗菌肽分子量小,大部分具有12~50個氨基酸,具有強堿性、熱穩(wěn)定性以及廣譜抗菌性等特點,在農(nóng)業(yè)、醫(yī)藥等領(lǐng)域受到廣泛的重視,全球科研人員對抗菌肽的作用機制、藥物療效、安全性等進行了深入的研究,并且建立了抗菌肽數(shù)據(jù)庫(APD:http://aps.unmc.edu/AP/main.php)[13]。目前,該數(shù)據(jù)庫已收錄2 700多種抗菌肽,其中272種抗菌肽來源于細菌,4種抗菌肽來源于古細菌,8種抗菌肽來源于原生生物、13種抗菌肽來源于真菌,335種抗菌肽來源于植物,2 043種抗菌肽來源于動物。這些抗菌肽具有抗細菌、抗病毒、抗真菌、抗寄生蟲、抗癌、抗原蟲等活性。
研究發(fā)現(xiàn),多種抗菌肽具有抗病毒活性,如抗艾滋病病毒、皰疹病毒。LL-37是人體內(nèi)唯一的cathelicidins,通過直接的滅活試驗發(fā)現(xiàn),LL-37可以顯著降低幾種病毒對人的感染[14-17]。動物抗菌肽通過與病毒糖蛋白或細胞表面的病毒受體的相互作用,可以直接抑制病毒的感染,也可以通過激活其他抗病毒反應(yīng)來間接地抑制病毒感染[18]。
本試驗測定了PRRSV在感染豬后的不同時間段內(nèi),脾臟、肺、肺門淋巴結(jié)中cathelicidins表達量的變化情況。研究結(jié)果表明,PR39、PG1、PG1-5、PMAP23的表達受到PRRSV的影響,豬感染PRRSV后,脾臟、肺、肺門淋巴結(jié)中的PR39、PG1、PG1-5、PMAP23表達量上調(diào),其中PRRSV對PR39與PG1-5表達量的影響較大。下一步工作合成PR39、PG1、PG1-5、PMAP23多肽,測定抗菌肽對PRRSV的滅活效果以及對細胞的毒性,為PRRSV新藥物的開發(fā)提供新的思路。
[1]Sang Y M,Ruchala P,Lehrer R I,et al. Antimicrobial host defense peptides in an arteriviral infection:differential peptide expression and virus inactivation[J]. Viral Immunology,2009,22(4):235-242.
[2]劉 偉,皮雄娥,王 欣. 抗菌肽與腸道健康研究進展[J]. 微生物學報,2016,56(10):1537-1543.
[3]Chu H,Pazgier M,Jung G,et al. Humanα-defensin 6 promotes mucosal innate immunity through self-assembled peptide nanonets[J]. Science,2012,337(693):477-481.
[4]汪以真. 動物源抗菌肽的研究現(xiàn)狀和展望[J]. 動物營養(yǎng)學報,2014,26(10):2934-2941.
[5]Wang Y P,Lu Z K,F(xiàn)eng F F,et al. Molecular cloning and characterization of novel cathelicidin-derived myeloid antimicrobial peptide fromPhasianuscolchicus[J]. Developmental and Comparative Immunology,2011(3):314-322.
[6]Feng F F,Chen C,Zhu W J,et al. Gene cloning,expression and characterization of avian cathelicidin orthologs,Cc-CATHs,fromCoturnixcoturnix[J]. FEBS Journal,2011,278(9):1573-1584.
[7]Wang Y P,Hong J,Liu X H,et al. Snake cathelicidin from bungarus fasciatus is a potent peptide antibiotics[J]. PLoS One,2008,3(9):e3217.
[8]Turner J,Cho Y,Dinh N N,et al. Activities of LL-37,a cathelin-associated antimicrobial peptide of human neutrophils[J]. Antimicrobial Agents and Chemotherapy,1998,42(9):2206-2214.
[9]廣慧娟. Cathelicidin家族抗菌肽研究進展[J]. 動物學研究,2012,33(5):523-526.
[10]韓 軍,楊漢春. 豬源抗菌肽的研究進展[J]. 中國獸醫(yī)雜志,2004,40(5):29-32.
[11]Cheng Q,Jiang Z G,Xu C C,et al. CpG oligodeoxynucleotide promotes protective immunity in the enteric mucosa and suppresses enterotoxigenicE.coliin the weaning piglets[J]. International Immunopharmacology,2010,10(10):1249-1260.
[12]Livak K J,Schmittgen T D. Analysis of relative gene expression data using realtime quantitative PCR and the 2-ΔΔCTmethod[J]. Methods,2001,25(4):402-408.
[13]Wang G S,Li X,Wang Z. APD2:the updated antimicrobial peptide database and its application in peptide design[J]. Nucleic Acids Research,2009,37(S1):D933-D937.
[14]Bergman P,Walter-Jallow L,Broliden K A,et al. The antimicrobial peptide LL-37 inhibits HIV-1 replication[J]. Current HIV Research,2007,5(4):410-415.
[15]Gordon Y J,Huang L C,Romanowski E G,et al. Human cathelicidin (LL-37),a multifunctional peptide,is expressed by ocular surface epithelia and has potent antibacterial and antiviral activity[J]. Current Eye Research,2005,30(5):385-394.
[16]Howell M D,Jones J F,Kisich K O,et al. Selective killing of vaccinia virus by LL-37:implications for eczema vaccinatum[J]. Journal of Immunology,2004,172(3):1763-1767.
[17]Howell M D,Gallo R L,Boguniewicz M,et al. Cytokine milieu of atopic dermatitis skin subverts the innate immune response to vaccinia virus[J]. Immunity,2006(24):341-348.
[18]Klotman M E,Chang T L. Defensins in innate antiviral immunity[J]. Nature Reviews Immunology,2006(6):447-456.

- 江蘇農(nóng)業(yè)科學的其它文章
- 不同經(jīng)濟發(fā)展水平區(qū)域的農(nóng)戶宅基地退出意愿
——基于江蘇省577個農(nóng)戶的調(diào)研 - 職業(yè)糧農(nóng)適度規(guī)模經(jīng)營的扶持政策需求及其影響因素分析
- 基于消費者視角的雞蛋質(zhì)量安全可追溯意愿分析
——基于新疆5市365位消費者的調(diào)查 - 農(nóng)村土地承包經(jīng)營權(quán)抵押貸款意愿的影響因素
——基于農(nóng)戶視角的調(diào)研分析 - 質(zhì)量控制、供貨能力對“農(nóng)超對接”參與行為的影響
——基于合作社視角 - 基于Logistic模型的連片貧困區(qū)農(nóng)戶精準扶貧滿意度的影響因素
——以湖北大別山片區(qū)為例