連續2代銅脅迫處理對海州香薷種子形態大小、萌發及幼苗生長的影響
2018-03-12 07:29:01梁慧娟李鈞敏
江蘇農業科學 2018年2期
梁慧娟, 閆 明, 李鈞敏
(1.山西師范大學生命科學學院,山西臨汾 041004; 2.浙江省植物進化生態學與保護重點實驗室,浙江臺州 318000; 3.臺州學院生態研究所,浙江臺州 318000)
目前,由于人類活動(如采礦等)所帶來的土壤重金屬污染已經成為全球環境污染治理的難題與焦點[1]。重金屬銅(Cu)既是植物光合作用、電子呼吸傳遞、蛋白質代謝、抗氧化反應等代謝過程所必需的元素,同時過量的銅又對植物生長和發育具有強烈抑制作用[2]。因此,尋找銅耐受或超積累植物已成為銅污染區域的植被恢復或修復的主要途徑之一[3]。
海州香薷(Elsholtziasplendens)為唇形科香薷屬的一年生草本植物,別稱“銅草”和“銅草花”,廣泛分布于我國的長江中下游地區。近年來的研究證實,海州香薷對銅具有較高的耐性和較強的累積能力,其根部銅含量高,在銅礦區生長異常旺盛,是礦區的優勢種之一,為銅礦的指示植物,是一種有潛力的修復植物[4-6]。當前較多研究集中在海州香薷對銅富集能力、光合作用、根系分泌物、銅結合蛋白質等一些生理生化機制上,對其后代適合度影響的研究較少,尤其是有關海州香薷的適應性進化幾乎沒有研究涉及。
種子作為植物的后代,其形態是植物在長期進化過程中對環境適應的結果[7],它與種子數量、擴散對策及幼苗競爭能力有密切關系,進而影響植物適合度的大小[8]。種子萌發是植物最早接受重金屬脅迫的階段,也是評價植物重金屬耐性的重要指標。在植物生長過程中,早期的幼苗大小是植物生存適合度的最主要指標[9],反映了子代幼苗建群的能力[10]。有學者等采用溶液培養的方法,發現海州香薷種子的發芽率隨著銅濃度的升高而先升后降,銅處理可以顯著抑制海州香薷胚根與胚軸的生長。目前尚未見通過多代培養試驗來分析銅脅迫對海州香薷后代適合度影響的相關報道。
本研究以經過連續2代重金屬銅脅迫的海州香薷種群和對照(同步培養2代,未經過銅脅迫處理的海州香薷種群)種子為供試材料,分析銅脅迫對海州香薷后代種子大小、種子萌發率及幼苗生長的影響,以更深入地認識該植物響應銅脅迫的生長與發育規律。
1 材料與方法
1.1 試驗材料
于2013年在湖北省紅安縣華河鎮的幾個自然村周圍,在無Cu污染的背景下,隨機采集相應的海州香薷種子。于2014年5月上旬對種子進行殺菌處理(0.5%次氯酸鈉)后放入48孔穴盤中萌發,待植物幼苗長至6葉苗齡時,取相似大小的幼苗進行移栽,采用筐栽方法進行重金屬馴化。培養基質由泥炭土、河沙、蛭石按 6 ∶3 ∶1的比例混合而成,基質均于121 ℃滅菌2 h。Cu處理為添加CuSO4(分析純)溶液(0.16 mol/L)至終濃度達1 000 mg/kg的土壤;對照(CK)添加等體積的無菌水,作為不添加Cu處理。處理后將土壤放在大棚中平衡1周后移栽海州香薷,每筐栽種80株。每個處理6次重復。于同年12月底收獲種子,備用。2015年5月開始參照2014年的方法對已收獲的海州香薷種子進行第2次重金屬馴化,于同年12月底收獲二代試驗種子,備用。
1.2 種子大小及形態分析
隨機抽取2個處理二代海州香薷種子各1 000粒,用精確度為萬分之一的天平稱其千粒質量,每筐種子重復3次。再隨機選取10粒種子,均勻撒在Epson掃描儀的玻璃板上,掃描獲得圖像后,用SmartGrain軟件直接分析種子的形態指標,獲得種子表面積、周長、長度、寬度和長寬比。
1.3 種子萌發試驗
隨機抽取2個處理的二代海州香薷種子,用0.3%次氯酸鈉表面消毒20 min,用蒸餾水沖洗干凈后,置于恒溫光照培養箱中萌發(25 ℃、16 h光照+20 ℃、8 h黑暗),用不同Cu2+濃度(0、5、20、100、250、500、1 000、2 000 μmol/L)溶液(主要成分為CuSO4·5H2O)處理9d。每個培養皿放置25粒種子,共3個重復。將處理后的種子以5×5的方陣形式整齊擺放在鋪有雙層濾紙的90 mm培養皿中,并添加等量的不同濃度的銅溶液,使濾紙完全濕潤,置于恒溫光照培養箱中萌發,于每天的固定時間統計種子發芽數,當胚根長度大于2 mm時計為萌發,并用移液槍統一補充適量銅溶液。在第3天統計發芽勢,在第8天統計發芽率。
1.4 幼苗生長指標測定
在第9天,用電子游標卡尺和電子天平測量幼苗的胚軸長度、胚根長度、胚軸鮮質量和胚根鮮質量,并分別按以下公式計算發芽率、發芽勢、發芽指數和活力指數等指標[11]:
發芽率(Gr)=(∑Gt/N)×100%;
發芽勢(Ge)=3 d內發芽種子數/N;
發芽指數(Gi)=(∑Gt/Dt);
活力指數(Vi)=Gi×S。
式中:Gt為在t天的發芽數,粒;Dt為相應的發芽時間,d;N為種子總數,粒;S為幼苗長度,mm。
1.5 數據分析
采用Excel 2010、SPSS 17.0對數據進行統計分析,采用獨立樣本t檢驗比較種群間各項指標平均數間差異的顯著性(α=0.05),采用Origin 8.5軟件輔助作圖。
2 結果與分析
2.1 銅脅迫對海州香薷種子形態及大小的影響
由表1可以看出,與對照相比,銅脅迫處理2代后海州香薷種子表面積、周長、長度和寬度均顯著低于對照種群,但種子長寬比與種子千粒質量沒有明顯變化。
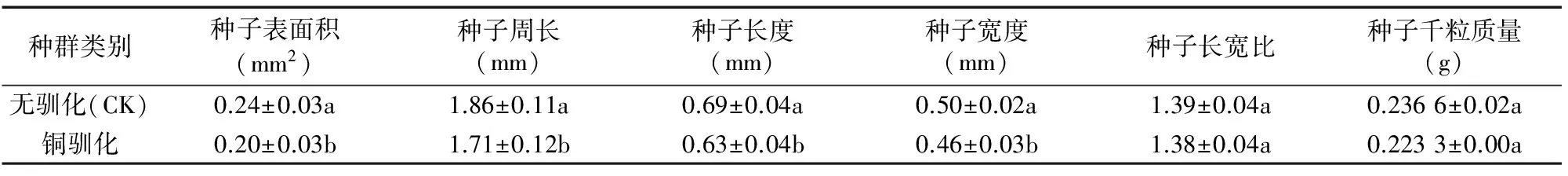
表1 銅馴化種群與無馴化種群海州香薷種子形態指標及千粒質量
注:表中數據格式為“平均值±SD”。同列數據后標有不同小寫字母者表示處理間存在顯著性差異(P<0.05)。
2.2 銅處理對不同種群海州香薷種子萌發的影響
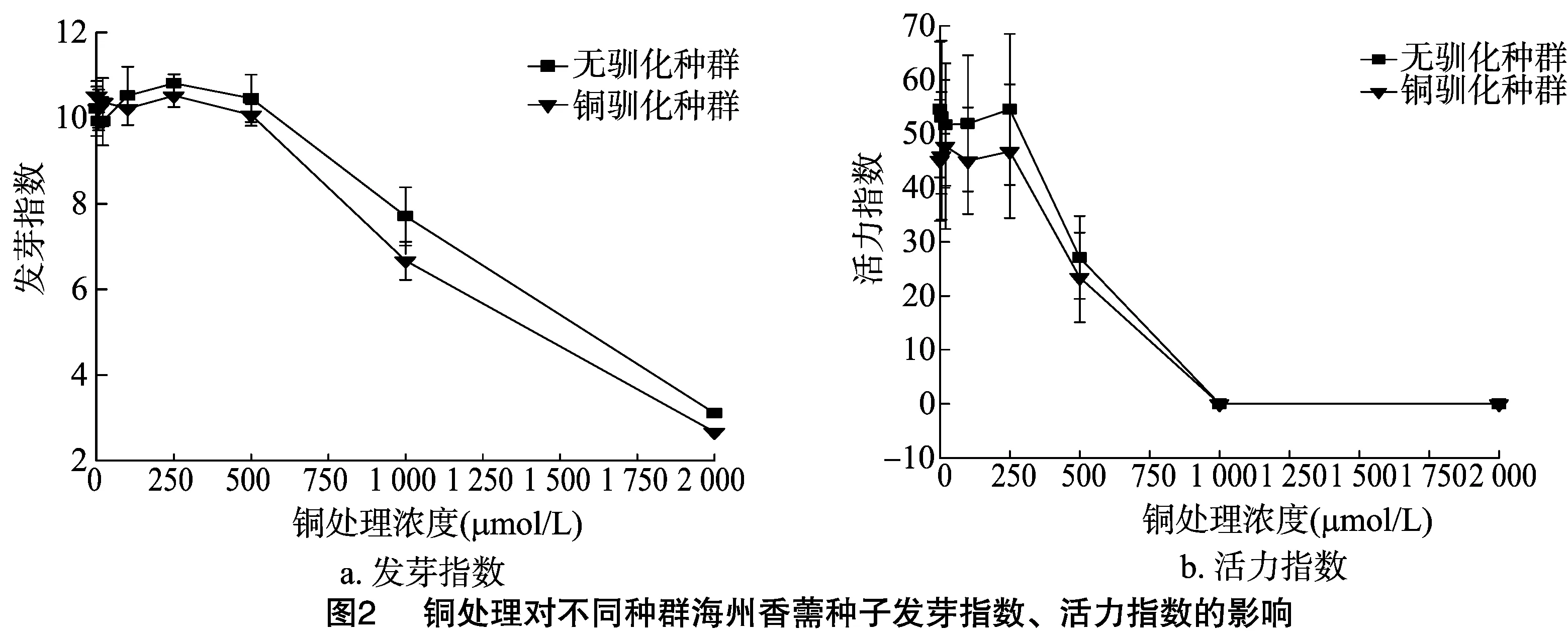
由圖1、圖2可見,隨著銅濃度增加,2個種群的海州香薷種子發芽率、發芽勢、發芽指數、活力指數大致呈先升高后降低的趨勢,表明高濃度的銅處理可以顯著抑制海州香薷種子的萌發。不管是在哪個銅濃度下,2個海州香薷種群的種子發芽率、發芽指數和第3天的發芽勢均不存在顯著差異,但銅脅迫處理種群的種子活力指數始終明顯低于對照種群。
2.3 銅處理對不同種群海州香薷幼苗生長的影響
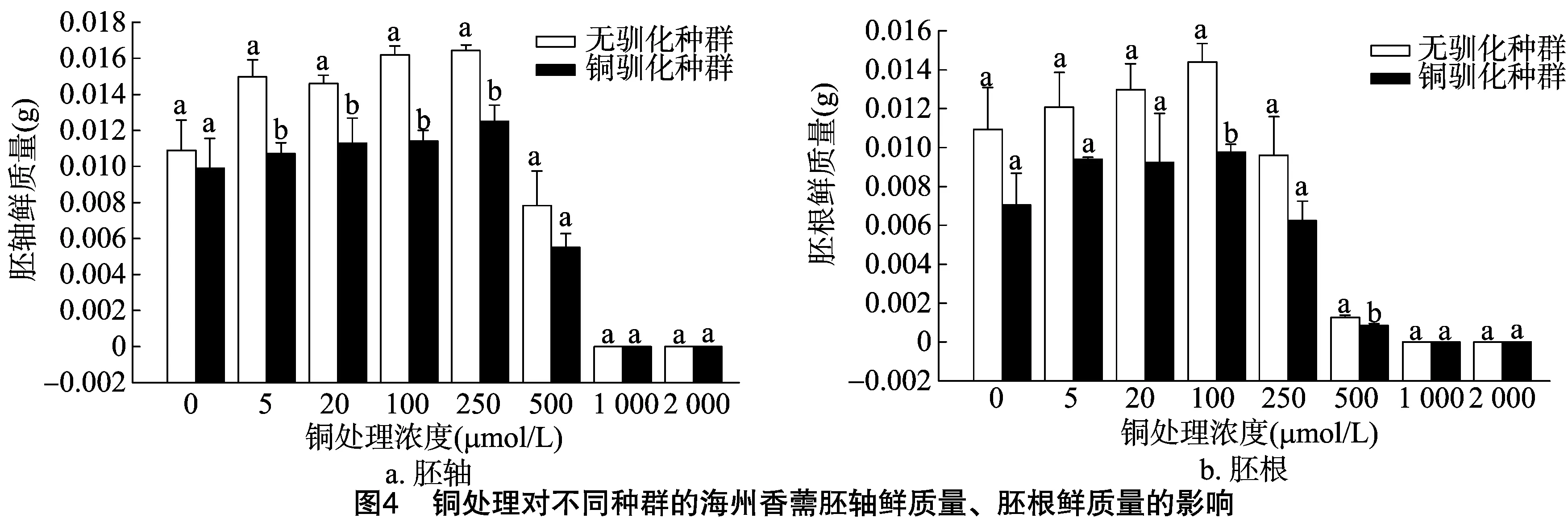
由圖3、圖4可以看出,隨著銅濃度的增加,海州香薷幼苗胚根和胚軸長度、鮮質量先逐漸升高再逐漸降低,表明高濃度的銅處理可以顯著抑制海州香薷的幼苗生長。當銅濃度為5、100 μmol/L時, 銅脅迫處理種群的胚軸長度顯著低于對照種群;但胚根長度在銅濃度為5、500 μmol/L時顯著高于對照種群。在銅濃度為5、20、100、250 μmol/L時,銅脅迫處理種群的胚軸鮮質量要顯著低于對照種群,而在銅濃度為100、500 μmol/L時的胚根鮮質量也顯著低于對照種群。


3 討論
種子萌發是健全的種子在充足的水分、適宜的溫度和足夠的氧氣等適宜條件下,能夠發芽并長成正常植株的能力。發芽率和發芽勢能較好地反映種子萌發的速度、整齊性,發芽指數、活力指數是種子萌發的綜合指標,能更全面地反映種子萌發期耐受重金屬的能力[11]。植物幼苗生長快慢可以反映植物生長形態的特征,也可以反映植物后期的生長狀況[12]。重金屬對種子萌發的影響存在低濃度下的刺激效應和高濃度下的抑制作用[13]。本研究發現,不管是銅脅迫處理過的海州香薷種群的種子還是對照種群的種子,其發芽率、發芽勢、發芽指數、活力指數、胚根和胚軸長度及鮮質量在響應不同濃度銅處理時均存在“低促高抑”的現象。過量的銅會對植物產生毒害,可降低植株的存活率、降低生物量、延遲開花和果實成熟時間以及籽粒的發育等[3]。魚小軍等發現,隨著銅濃度的增大,顯著抑制了7種豆科牧草種子的發芽及幼苗生長[14]。王丹等也證實,高含量銅對小白菜種子萌發、幼苗生長產生顯著抑制作用[15]。本研究發現,1 000 mg/kg銅脅迫處理2代的海州香薷種群的種子表面積、周長、長度和寬度均顯著低于對照種群;不管是在哪個銅濃度下,銅脅迫處理種群的種子活力指數始終明顯低于對照種群;在銅濃度為5、100 μmol/L 時,銅脅迫處理種群后代的胚軸長度顯著低于對照種群;在銅濃度為5、20、100、250 μmol/L時,銅脅迫處理種群后代的胚軸鮮質量要顯著低于對照種群;在銅濃度為100、500 μmol/L時,銅脅迫處理種群后代的胚根鮮質量顯著低于對照種群。這些結果表明,1 000 mg/kg重金屬銅連續處理2代可顯著抑制海州香薷種子的發育、種子萌發及幼苗生長。李月靈研究也發現,銅處理可以推遲海州香薷的始花期、開花高峰期、終花期,縮短花期持續時間,使植物花序寬、花序生物量、種子總數量、總質量顯著下降[3],從而降低了海州香薷后代的適合度。
重金屬會對生物產生的一定的影響,而生活在重金屬環境中的生物也會對重金屬產生一定的適應性[16]。馴化在動物與微生物研究中應用較多。近年來的研究發現,植物會在長期進化中演變出耐受重金屬的多種響應機制[17]。重金屬處理可以顯著降低種子千粒質量[3]。本研究發現,銅脅迫連續處理2代的海州香薷種群的千粒質量與對照種群相似,表明銅脅迫處理2代后,海州香薷種子的成分組成可能發生了適應性改變,雖然表面積變小,但質量變化不大。本研究還發現,在銅濃度為5、500 μmol/L時,銅脅迫處理2年的海州香薷種群幼苗的胚根長度顯著高于對照種群,表明銅脅迫處理2代后,海州香薷幼苗會提高胚根長度以增強幼苗對外界養分的捕獲能力,從而減緩銅脅迫對后代生長的抑制作用[3]。后續研究將繼續增加培養代數,以進一步確認海州香薷對銅脅迫的快速適應性進化發生的可能性及機制。
[1]Li J M,Jin Z X,Gu Q P. Effect of plant species on the function and structure of the bacterial community in the rhizosphere of lead-zinc mine tailings in Zhejiang,China[J]. Canadian Journal of Microbiology,2011,57(7):569-577.
[2]Kamali M,Pour M S,Moud A A M. Copper effects on growth parameters of hollyhock (AlthaearoseaL.)[J]. Journal of Ornamental and Horticultural Plants,2012,2(2):95-101.
[3]李月靈. 接種叢枝菌根真菌對海州香薷耐受銅脅迫的影響及機制[D]. 上海:上海師范大學,2014:59-73.
[4]柯文山,席紅安,楊 毅,等. 大冶銅綠山礦區海州香薷(Elsholtziahaichowensis)植物地球化學特征分析[J]. 生態學報,2001,21(6):907-912.
[5]柯文山,熊治廷,金則新,等. 海州香薷(ElsholtziahaichouensisSun)不同生態型Cu吸收和酸性磷酸酶活性差異[J]. 生態學報,2007,27(8):3172-3181.
[6]Ke W S,Xiong Z T,Chen S J,et al. Differences of Cu accumulation and Cu-induced ATPase activity in roots of two populations ofElsholtziahachowensisSun[J]. Environmental Toxicity,2008,23(2):193-199.
[7]周 旋,何正飚,康宏樟,等. 溫帶-亞熱帶栓皮櫟種子形態的變異及其與環境因子的關系[J]. 植物生態學報,2013,37(6):481-491.
[8]王桔紅,杜國禎,崔現亮,等. 青藏高原東緣61種常見木本植物種子萌發特性及其與生活史的關聯[J]. 植物生態學報,2009,33(1):171-179.
[9]武高林,杜國禎. 植物種子大小與幼苗生長策略研究進展[J]. 應用生態學報,2008,19(1):191-197.
[10]張麗坤,王 朔,馮玉龍. 紫莖澤蘭種子形態特征和萌發特性與其入侵性的關系[J]. 生態學報,2014,34(13):3584-3591.
[11]陳俊任,柳 丹,吳家森,等. 重金屬脅迫對毛竹種子萌發及其富集效應的影響[J]. 生態學報,2014,34(22):6501-6509.
[12]張凡凡,于 磊,魯為華,等. 不同馴化年限野生線葉野豌豆種子特性及幼苗耐寒性[J]. 草業科學,2013,30(11):1771-1777.
[13]陳 偉,張苗苗,宋陽陽,等. 重金屬脅迫對4種草坪草種子萌發的影響[J]. 草地學報,2013,21(3):556-563.
[14]魚小軍,張建文,潘濤濤,等. 銅、鎘、鉛對7種豆科牧草種子萌發和幼苗生長的影響[J]. 草地學報,2015,23(4):793-803.
[15]王 丹,魏 威,王松山,等. 銅、鉻單一及復合污染對小白菜種子萌發及根長的生態毒性[J]. 西北農林科技大學學報(自然科學版),2010,38(12):63-68,74.
[16]徐 池. 重金屬Cu對蚯蚓的馴化研究[D]. 南京:南京農業大學,2012:38-46.
[17]李 洋,于麗杰,金曉霞. 植物重金屬脅迫耐受機制[J]. 中國生物工程雜志,2015,35(9):94-104.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
