Map3k1基因下調對B6小鼠眼瞼角質形成細胞增殖和遷移能力的影響*
2018-03-30 00:47:22劉莉莉夏婷婷朱順星
重慶醫學 2018年9期
景 瑾,劉莉莉,尤 杰,夏婷婷,張 進,劉 春,朱順星△
(1.南通大學杏林學院,江蘇南通 226001;2.南通大學實驗動物中心,江蘇南通 226001)
研究表明眼瞼發育異常主要是由于眼瞼角質形成細胞遷移受阻所致[1]。大量資料顯示,Map3k1基因在小鼠眼瞼形態建成過程中起到關鍵作用。Map3k1是絲裂原活化蛋白激酶(MAPK)家族中的重要成員,其表達的蛋白細胞外信號調節激酶1(MEKK1)具有廣泛的生物學功能。MEKK1蛋白是MAPK信號傳導通路的結點,可以調節JNK[2]、ERK1/2[3]、P38[4]、IKK-NFκB[5]等信號通路,這些信號通路參與調控相關細胞的增殖、分化、凋亡、遷移和侵襲等[2-5],對細胞的生存、生長起著至關重要的作用。本研究擬采用RNA干擾技術,實現對Map3k1的特異性沉默,來檢測其對B6小鼠眼瞼角質形成細胞生物學功能的影響。
1 材料與方法
1.1材料
1.1.1實驗動物 B6(C57BL/6,B6)小鼠購自南通大學實驗動物中心,飼養在清潔級屏障動物房內, 室內溫度為(23±2)℃, 濕度控制為(55±5)%,自由采食和飲水,室內照明采用12 h∶12 h 明暗交替,定期更換籠具、墊料。
1.1.2主要儀器及試劑 CO2細胞培養箱購自Thermo公司(美國);倒置相差顯微鏡購自Leica公司(德國);抗菌藥物、大豆胰蛋白酶抑制劑購自Amresco公司(美國);角質形成細胞無血清培養基(DK-SFM)、0.25%胰蛋白酶-乙二胺四乙酸(EDTA)購自Gibco公司(美國);中性蛋白酶Ⅱ購自Roche公司(瑞士);24孔細胞培養板購自Corning公司(美國);質粒抽提試劑盒購自Omega公司(美國);Mouse monoclonal to MEKK1購自Abcam公司(英國);Anti-Mouse IgG、HRP-linked Antibody購自Cell Signaling公司(美國)。
1.2方法
1.2.1B6小鼠眼瞼角質形成細胞的分離和原代培養 取1 d B6小鼠,超凈臺中取眼瞼,在75%乙醇中清洗,然后在0.01 mol/L磷酸鹽緩沖液(PBS)中清洗;將眼瞼放入添加有中性蛋白酶Ⅱ(3 mg/mL)和抗菌藥物的DK-SFM 培養液中,4 ℃消化18 h;加入等體積的PBS,分離真皮和表皮,將表皮充分剪碎,加入0.25%胰蛋白酶-EDTA,37 ℃消化約10 min;再加入大豆胰蛋白酶抑制劑終止消化,1 200 r/min 室溫離心5 min,棄上清液,PBS清洗一遍,加入含有10 ng/mL表皮生長因子(EGF)的DK-SFM 培養液重懸細胞;調整細胞密度為2×105個/mL;將細胞接種于12孔板,37 ℃、5% CO2培養箱中培養;24 h后更換培養液,每隔1 d換液1次。
1.2.2靶向沉默Map3k1基因的amiRNA表達載體 本課題組在前期實驗中已成功構建靶向沉默Map3k1基因的amiRNA表達載體,并篩選出干擾效率最高的質粒[6]。Map3k1基因序列的miRNA靶向序列如下,miRNA3-F:5′-TGC TGA TCC ATC TCC TTT AAT AGG GAG TTT TGG CCA CTG ACT GAC TCC CTA TTA GGA GAT GGA T-3′;miRNA3-R:5′-CCT GAT CCA TCT CCT AAT AGG GAG TCA GTC AGT GGC CAA AAC TCC CTA TTA AAG GAG ATG GAT C-3。
1.2.3菌種復性與質粒抽提 -80 ℃冰箱中取出之前所保存的菌種,冰上溶解;吸取50 μL于1 mL LB液體培養基(含Amp+)中,37 ℃、220 r/min培養1 h;菌種狀態良好時,直接以劃線法涂LB平板[含氨芐青霉素(Amp+)],倒扣平板,37 ℃培養過夜;用10 μL槍頭挑取平板上單克隆菌落至5 mL LB液體培養基(含Amp+)中,37 ℃、220 r/min搖床培養過夜,1 mL保種,1 mL送上海生工生物工程股份有限公司進行測序;測序正確的再次搖菌,按試劑盒說明書提取質粒,獲得的質粒,核酸蛋白定量儀測濃度,-20 ℃存儲備用;分裝部分抽提的質粒DNA,送上海生工生物工程股份有限公司進行測序分析。
1.2.4Map3k1 amiRNA-3質粒轉染B6小鼠眼瞼角質形成細胞 取2 μL Map3k1 amiRNA-3質粒(568 ng/μL)溶于100 μL Opti-MEM基培中,取2 μL Lipofectamin 2000溶于100 μL Opti-MEM基培中,分別輕輕混勻,室溫各孵育5 min;將兩個混合液輕輕混勻,室溫孵育20 min;角質形成細胞換液,更換DK-SFM 培養液800 μL;混合液加入12孔板中,輕輕混勻,細胞于37 ℃,5% CO2培養箱中培養6 h后更換DK-SFM 培養液。細胞轉染后24、48、72 h分別提取總RNA和總蛋白。
1.2.5Real-Time PCR檢測Map3k1 mRNA相對表達量 根據實驗時間點要求提取細胞總RNA,采用TRIzol法提取。使用逆轉錄試劑盒RevertAidTMFirst Strand cDNA Synthesis Kit(Roche公司)進行逆轉錄反應。根據Map3k1基因mRNA設計RT-PCR反應引物如下,GAPDH-F:5′-GGA GCG AGA CCC CAC TAA C-3′,GAPDH-R:5′-GGC GGA GAT GAT GAC CCT-3′;Map3k1-F:5′-GGT CCT AAG AGG TCA GCA GTA TG-3′,Map3k1-R:5′- GGA CAG GTG TGA CGG GAT G-3′。RT-PCR反應采用SYBR Green MiX試劑,反應體系如下:SYBR Green mix 10 μL、 Primer-F(10 pmol)0.5 μL 、Primer-R(10 pmol)0.5 μL 、cDNA 1 μL、ddH2O 8 μL。RT-PCR反應條件為:95 ℃,5 min;45個循環(95 ℃,15 s;60 ℃,20 s;72 ℃,30 s),每個循環在延伸階段收集熒光;產物熔解曲線分析:95 ℃,15 s;60 ℃,1 min;95 ℃,15 s。各組細胞目的基因均以內參基因GAPDH進行校正,為了減少誤差,實驗重復3批樣本,每個樣本均重復3次,使用2-ΔΔCT法進行相對定量分析。
1.2.6Western blot檢測Map3k1蛋白相對表達量 根據實驗時間點要求提取各組細胞總蛋白;并取適量以BCA蛋白定量試劑盒測定蛋白濃度;按說明書配制分離膠及濃縮膠:Gapdh配制12%的分離膠,Map3k1配制6%的分離膠;常規程序電泳,直至目的蛋白充分跑開;電泳結束后,Gapdh以300 mA,60 min的條件轉膜,Map3k1以110 V,120 min的條件轉膜;轉膜結束后將膜取出,置于封閉液中室溫封閉2 h;在膜上滴加適量1∶1 000稀釋的一抗,4 ℃孵育過夜;用TBST洗滌1次,TBS洗滌2次;加二抗室溫孵育2 h;再用TBST洗滌1次,TBS洗滌2次,取1 mL ECL顯色液顯色1 min。
1.2.7四甲基偶氮唑鹽(MTT)法檢測B6小鼠眼瞼角質形成細胞的增殖水平 取對數生長期細胞,胰酶消化后離心收集,加入DK-SFM 培養液重懸細胞,細胞計數調整為 2×104/mL。將制備好的細胞懸液邊輕輕混勻邊加入96 孔板,每孔加入 100 μL;將接種好的細胞培養板放入培養箱中培養過夜,第2天做轉染,分為Ctrl Map3k1組與Map3k1 amiRNA-3組,Ctrl Map3k1組只加入培養基不加細胞,每組設5個復孔取均值,實驗重復 3 次。在轉染24、48、72 h后MTT法測細胞活力;不同時間Map3k1 amiRNA-3組及Ctrl Map3k1組每孔加入10 μL MTT 溶液,繼續培養 4 h;再加入 100 μL Formanzan溶解液,培養箱繼續孵育約4 h,顯微鏡下觀察待結晶物充分溶解后,在酶標儀吸光度(A)570 nm處測量各孔的A值。
1.2.8劃痕實驗檢測B6小鼠眼瞼角質形成細胞的遷移能力 取對數生長期細胞接種到24孔板,每孔接種1×106個細胞,并設3個重復;Map3k1 amiRNA-3組與Ctrl Map3k1組于轉染36 h后,棄原培養液,加入含有10 μg/mL 絲裂霉素C的DK-SFM培養液培養12 h;然后用100 μL的槍頭在每個孔的中央進行十字劃痕;吸去培養液,0.01 mol/L PBS 輕洗;加入含有10 ng/mL EGF的DK-SFM 培養液,于十字劃痕處顯微鏡觀察拍照,記為0 h,每孔照數個不同的位置;培養24 h 后,依據記錄的大致位置再次拍照。
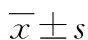
2 結 果
2.1Map3k1 amiRNA-3干擾質粒 測序結果與預期完全吻合(圖1),miRNA序列正確插入pcDNA-miRNA載體中。
2.2Map3k1 amiRNA-3質粒對Map3k1基因的干擾效率 Real-time PCR結果(圖2A)表明在轉染Map3k1 amiR-3質粒后48 h,B6小鼠眼瞼角質形成細胞Map3k1基因mRNA的表達量下降最為顯著,干擾效率高達70%(P<0.01);Western blot結果(圖2B、C)表明在轉染Map3k1 amiR-3質粒后72 h,B6小鼠眼瞼角質形成細胞Map3k1基因蛋白的表達量下降最為顯著,干擾效率高達70%(P<0.05)。

A:Real-Time PCR檢測Map3k1 mRNA相對表達量;B:Western blot檢測Map3k1 amiRNA-3蛋白的相對表達量;C:Map3k1 amiRNA-3蛋白的相對表達量;a:P<0.05,b:P<0.01
圖2 Real-Time PCR、Western blot檢測Map3k1 amiRNA-3的干擾效率
2.3MTT法檢測B6小鼠眼瞼角質形成細胞的增殖水平 細胞增殖水平檢測結果顯示:Map3k1 amiRNA-3組在轉染24、48、72 h細胞增殖水平顯著低于Ctrl amiRNA組(P<0.05)。見表1、圖3。
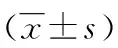
表1 MTT法不同時間細胞增殖水平檢測
a:P<0.05,b:P<0.01,與Ctrl amiRNA組比較
2.4劃痕實驗檢測B6小鼠眼瞼角質形成細胞的遷移能力 Map3k1 amiRNA-3組角質形成細胞,在劃痕后24 h,其穿過劃痕區的細胞數量與相對距離均明顯低于轉染空載體的Ctrl amiRNA組(圖4),定量分析發現Ctrl amiRNA組遷移的細胞數為(615±39)個,Map3k1 amiRNA-3組僅有(325±34)個,差異有統計學意義(P<0.05);細胞遷移劃痕區的相對距離,Ctrl amiRNA組為(0.339±0.015),MAp3k1 amiRNA-3組為(0.181±0.027),兩組差異有統計學意義(P<0.05)。

圖4 劃痕實驗檢測B6小鼠眼瞼角質形成細胞的遷移能力
3 討 論
眼瞼是眼球前的軟組織,當眼受到外界各種刺激時,可通過中樞神經的指揮及時將其關閉,從而阻擋外來異物或強光對眼睛造成傷害,對眼的發育和保護具有重要作用。若眼瞼發育異常便會受到各種眼部疾病的困擾,如角膜病、白內障、視力減退等。研究表明眼瞼發育異常主要是由于眼瞼角質形成細胞遷移受阻所致[1]。有研究發現通過MEKK1的轉導,可以引發角質形成細胞的遷移,MEKK1對角質形成細胞的運動有著至關重要的作用[7]。
RNA干擾是由雙鏈RNA(dsRNA)誘發同源mRNA高效且特異性降解的現象,可以特異性剔除或關閉目的基因的表達,該技術具有成本低廉、高效率、高特異性、操作方便等優勢[8-10],已成為各大實驗室探索基因功能的常用技術。
Map3k1基因位于5q11.2,其編碼的蛋白稱為MEKK1,全長196×103。MEKK1是MAPK家族中的重要成員,具有Ser/Thr激酶活性,其C-端為催化結構域,N-端為調節結構域,可以磷酸化與其結合的下游靶蛋白,從而介導蛋白間的相互作用并影響其行為和生物學功能。MEKK1蛋白是MAPK信號傳導通路的結點,可以調節JNK[2]、ERK1/2[3]、P38[4]、IKK-NFκB[5]等信號通路,這些信號通路參與調控相關細胞的增殖、分化、凋亡、遷移和侵襲等[2-5]。Map3k1對細胞的發生、發展起著至關重要的作用,有研究表明Map3k1具有可決定細胞命運的開關樣功能,即促進和抑制細胞凋亡的雙重作用[11]。而MEKK1 作為幾個 MAPK 通路的上游調節因子,亦涉及不同種類和細胞類型的生物學反應,包括角質形成細胞的分化、T細胞活化和壓力刺激誘導的細胞凋亡等。因而,本研究擬采用RNA干擾技術,實現對Map3k1的特異性沉默,來檢測其對B6小鼠眼瞼角質形成細胞生物學功能的影響。期許為更好的理解、掌握Map3k1在B6小鼠眼瞼角質形成細胞中的作用,及其在眼瞼發育過程中的作用提供理論依據。
首先通過前期實驗中已成功構建靶向沉默Map3k1基因的amiRNA表達載體[6],實現對Map3k1的特異性沉默,干擾效率高達70%,為研究B6小鼠眼瞼角質形成細胞生物學效應在干擾Map3k1前后的變化奠定了基礎。
Map3k1基因參與調節各種類型的細胞運動遷移,研究發現Map3k1基因敲除的新生鼠由于上皮細胞遷移缺陷,導致出生時眼瞼不能閉合[12],說明Map3k1基因在小鼠眼瞼形態建成過程中起到關鍵作用。Map3k1表達的缺乏可減少細胞的遷移和侵襲能力[13]。在本實驗中發現靶向抑制Map3k1的表達后,B6小鼠眼瞼角質形成細胞的遷移能力被顯著抑制,無論是遷移的距離還是遷移的細胞數量都顯著下調(P<0.05)。此外,還發現Map3k1的表達被抑制后,會影響B6小鼠眼瞼角質形成細胞的增殖能力(P<0.05)。這些結果表明靶向抑制Map3k1的表達能夠廣泛的影響B6小鼠眼瞼角質形成細胞的生物學行為,從而影響眼瞼的發育。
[1]KATO S,MOHRI Y,MATSUO T,et al.Eye-open at birth phenotype with reduced keratinocyte motility in LGR4 null mice[J].FEBS Lett,2007,581(24):4685-4690.
[2]LU H J,NING X J,TAO X E,et al.MEKK1 associated with neuronal apoptosis following intracerebral hemorrhage[J].Neurochem Res,2016,41(12):3308-3321.
[3]CHENG Y,LIN C H,CHEN J Y,et al.Induction of connective tissue growth factor expression by hypoxia in human lung fibroblasts via the MEKK1/MEK1/ERK1/GLI-1/GLI-2 and AP-1 pathways[J].PLoS One,2016,11(8):1-23.
[4]HUANG D S,LI X J,SUN L,et al.Regulation of hippo signalling by p38 signalling[J].J Mol Cell Biol,2016,8(4):328-337.
[5]AL-SADI R,GUO S H,YE D M,et al.TNF-alpha modulation of intestinal tight junction permeability is mediated by NIK/IKK-alpha axis activation of the canonical NF-kappa B pathway[J].Am J Pathol,2016,186(5):1151-1165.
[6]王勝潔,劉春,汪海峰,等.靶向抑制MAP3K1對小鼠乳腺癌細胞生物學行為的影響[J].黑龍江畜牧獸醫,2016,290:26-31.
[7]XIA Y,KAO W W.The signaling pathways in tissue morphogenesis:a lesson from mice with eye-open at birth phenotype[J].Biochem Pharmacol,2004,68(6):997-1001.
[8]SUGAHARA R,TANAKA S,JOURAKU A,et al.Geographic variation in RNAi sensitivity in the migratory locust[J].Gene,2017,605:5-11.
[9]KAMATH R S,FRASER A G,DONG Y,et al.Systematic functional analysis of the Caenorhabditis elegans genome using RNAi[J].Nature,2003,421(6920):231-237.
[10]VOLPE T,KIDNER C,HALL I M,et al.Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi[J].Science,2002,297(5588):1833-1837.
[11]ZEBROWSKI D C,ALCENDOR R R,KIRSHENBAUM L A,et al.Caspase-3 mediated cleavage of MEKK1 promotes p53 transcriptional activity[J].J Mol Cell Cardiol,2006,40(5):605-618.
[12]YUJIRI T,WARE M,WIDMANN C,et al.MEK kinase 1 gene disruption alters cell migration and c-Jun NH2-terminal kinase regulation but does not cause a measurable defect in NF-kappa B activation[J].Proc Natl Acad Sci U S A,2000;97:7272-7277.
[13]STEED E,ELBEDIWY A,VACCA B,et al.MarvelD3 couples tight junctions to the MEKK1-JNK pathway to regulate cell behavior and survival[J].J Cell Biol,2014,204(5):821-838.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
